
Amina Darwish Mahmoud
Department of Biological and Geological Sciences, Faculty of Education, Alexandria University, Egypt
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 21 | Page No.: 1830-1836







ABSTRACT
Body temperature, microhabitat selection and thermoregulatory behaviour of field- active Acanthodactylus boskianus were evaluated for two sites at approximately equal elevation in El-Omayed region, west of Alexandria, on the west Mediterranean coastal region of Egypt: A. boskianus at coastal sand dunes had significantly lower body temperatures than those at second rocky ridge. The slope of a regression of body temperature on air temperature was significantly different between the two populations. The microhabitats in which individuals were first observed correlated with body temperatures at the coastal sand dunes, but not at the second rocky ridge. These data suggest that environmental temperature differences between sites influenced body temperature, thermoregulatory behaviour and microhabitat selection in A. boskianus.
PDF Abstract XML References Citation
How to cite this article
Amina Darwish Mahmoud, 2003. Variation in Thermal Relations Between Two Populations of the Sand Lizard, Acanthodactylus boskianus Living in Thermally Divergent Habitats of the Same Altitude. Pakistan Journal of Biological Sciences, 6: 1830-1836.
DOI: 10.3923/pjbs.2003.1830.1836
URL: https://scialert.net/abstract/?doi=pjbs.2003.1830.1836
DOI: 10.3923/pjbs.2003.1830.1836
URL: https://scialert.net/abstract/?doi=pjbs.2003.1830.1836
INTRODUCTION
In the half-century since Cowles and Bogert (1944) published their pioneering study of desert reptiles, physiological ecologists have worked to evaluate the extent and effectiveness of behavioural temperature regulation by field-active animals. Regulation allows some reptiles to maintain a high and relatively constant body temperature, Tb (Avery, 1982 and Huey, 1982), which may produce optimal organism performance in the field (Huey and Stevenson, 1979). Most reptiles regulate Tb behaviourally, although physiological adjustment may also be important (Bartholomew, 1982). Many other species of diurnal reptiles maintain high and relatively constant Tb’s despite wide fluctuations in environmental heat loads. They achieve these mainly by adjustments of their behaviour, in particular, restriction of activity times, shuttling between hot and cool microhabitats and postural modifications that alter the rates of heating and cooling (Hertz and Huey, 1981; Stevenson, 1985; Diaz, 1991; Shine and Madsen, 1996; Christian, 1998; Du et al., 2000 and Dzialowski and O’Connor, 2001a).
A large literature has established that the primary mechanisms of behavioural temperature regulation include the restriction of activity times, the selection of thermally appropriate microhabitats and postural adjustments that alter the rates of heat exchange (Hertz and Huey, 1981; Van Damme et al., 1989; Castilla and Bauwens, 1991, Shine and Madsen, 1996; Hussein, 1998 and Dzialowski and O’Connor, 2001 b). Under field conditions, reptiles often use these behaviours simultaneously, but few researchers have attempted to estimate their relative contributions to a regulated Tb (Porter et al., 1973; Muth, 1977). The contention that all species maintain a narrow range of body temperatures through physiological and behavioural adjustments has been modified by research showing that this range can be broadened when species are examined across seasons and populations (Huey et al., 1977; Grant and Dunham, 1988; Smith and Ballinger, 1994). For example, Bashey and Dunham (1997) have recently shown that microhabitat use and activity patterns differed between high elevation and low elevation populations of Cophosaurus texanus. These studies suggest that the thermal environment have the potential to influence the behaviour and physiology of lizards.
The lacertid lizard, Acanthodactylus boskianus is an abundant species that inhabits the semi-arid areas along the Mediterranean coast of Egypt. Like many other small diurnal lizards living in open habitats, exhibit high levels of aboveground activity and they were also found climbing the shrubs or sheltering under plant cover. The goal of the present study was to determine the influence of the thermal environment on body temperatures and thermoregulatory behaviour of this species and to evaluat the microhabitat use of lizards in thermally divergent habitats at the same elevation in El- Omayed region, part of western Mediterranean desert of Egypt. This work also estimates the relative contributions of microhabitat selection to temperature regulation, which are of paramount importance, too most aspects of reptilian ecology and behaviour. We hypothesize that the difference of the environmental temperatures between the thermally divergent habitats may explain many of the observed characteristic thermal differences in this species of lizards living on the same altitude.
MATERIALS AND METHODS
Study site and species: The study site is El-Omayed region, about 83 km west of Alexandria on the western Mediterranean coastal desert of Egypt (Fig. 1). The area is a part of El-Omayed Biosphere Reserve that covers a total of about 700 km2. This region has been selected for this study because of its characteristic features and the variety of its habitats. The habitat is composed of three parallel ridges in the area; a sand dunes coastal ridge and two consolidated ridges. The depressions formed between these ridges are flat composed of patches of bar sandy soil (Abdel-Kader and Coll, 1981). All data were collected from two sites at similar elevations (approximately 15 m above sea level). The studied sites are: 1) coastal sand dunes (CSD) which present active dunes on consolidated material and 2) second rocky ridge (SRR) summit of stretched hill (Eolithic limestone, ancient consolidated dune). The sites are separated by about 3.2 km, by first rocky ridge and by some depressions (Fig. 2).
Acanthodactylus boskianus (Reptilia, Squamata, Lacertidae) is an abundant species in the study site. Generally, the genus Acanthodactylus comprises the most lacertid species in this region (Michael et al., 1992).
A. boskianus is a small-sized lacertid (snout-vent length of adults: 60-80 mm; tail: 100-140 mm), diurnal, moves with great rapidity in open habitats and spend most of their activity period in open sandy habitat, climbing the small shrubs or sheltering under the branches of the plants. The plant cover in these sites are sparse and various. Ammophila arenaria and Ononis vaginalis are indicator species of the coastal dunes habitat and Herniaria hemistemon and Scorzonera alexandrina are indicator species for the rocky ridge habitat (Kamal, 1982).
Measurements: The fieldwork was conducted approximately two days weekly during the seasonal activity time from June to August 2002. The work was restricted at the morning and early afternoon hours (from 0800 to 1300 h). Body temperatures (Tb) were taken from active lizards that were chased less than 10 m. Nearly all individuals were sampled once because only previously unsampled areas were visited during each survey. Lizards were noosed and Tb’s taken within 10 sec from individuals handled. The lizards that we observed invariably retreated quickly to shelter under the plant branches. The sheltering lizards were captured by lifting up the branches and handily caught them. Measurements to the nearest 0.1°C were made using quick reading cloacal thermometer.
 | |
| Fig. 1: | A location map of the study site |
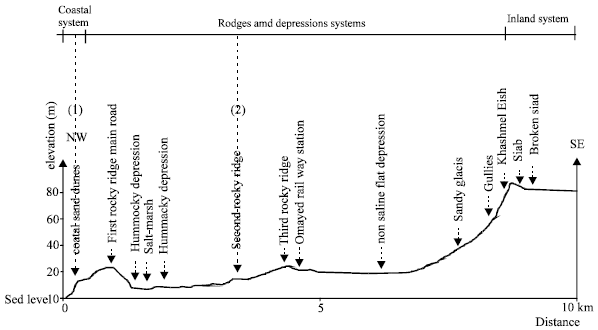 | |
| Fig. 2: | Location of physiographic units of El-Omayed region along the westren Mediterranean coastal literal of Egypt showing the location of the two study sites: 1) coastal sand dunes and 2) second rocky ridge |
Since there were no size or sexual differences in body temperatures (Hussein, 1992), none of the data are separated on the basis of age class or sex. Air temperature (Ta) was measured at 1 cm above the soil surface to the nearest 0.1°C by quick- reading thermometer. Soil temperatures (Ts) were recorded from just below the soil surface, with only approximately one layer of soil particles covering the top of the bulb. Each of these was taken at the location where a lizard was first observed.
Six categories were used to note the body condition and microhabitat where each lizard was first sighted: full sun (basking in the open habitat of the full sun), partial sun (basking in the full sun with very small shaded part), edge (< 50% estimated shade), shade (> 50% estimated shade), arboreal (within canopy of shrub) and sheltering (sheltering under the shrub).
ANOVA tests (Sokal and Rohlf, 1981) were used to analyze intrapopulational differences in Ta, Ts, Tb and body condition. For each population ANOVA tests were used to determine 1) the correlation between Tb and Ta; 2) the correlation between microhabitat and Tb; 3) the correlation of month on Tb, Ta and Ts and 4) correlation between time of day, Ta and Ts and microhabitat. T-test was also employed to compare the slopes of the regression of Tb on Ta in each population.
RESULTS
The averages air temperatures (Ta), soil temperatures (Ts) and body temperatures (Tb) in two populations of the desert lizard (Table 1). A. boskianus at coastal sand dunes (CSD) had significantly lower Tb’s than those at second rocky ridge (SRR). Air temperatures were also significantly lower at CSD than at SRR, as were Ts’s (Table 1).
Body temperature was positively correlated with Ta at both CSD and SRR, although Tb followed Ta more closely at CSD than at SRR (CSD: df = 172, r2 = 0.46, P < 0.001; SRR: df = 51, r2 = 0.39, P < 0.001); the slopes of the regressions differed significantly (t = 5.99, P < 0.001). At CSD, Tb (P = 0.17), Ta (P = 0.33) and Ts (P =0.76) did not vary among months. Conversely, Tb (P = 0.005), Ta (P = 0.0004) and Ts (P = 0.0004) were influenced by month at SRR. On the other hand, regression of Tb on Ta for A. boskianus from CSD was higher (0.77) than the slope calculated for A. boskianus at SRR (0.41) (Fig. 3).
Microhabitat was correlated with Tb at CSD (F = 6.55, P<0.001) but not at SRR (P>0.41, Fig. 4). Microhabitat selection varied with time of day in both populations (Fig. 5a). In early morning hours, CSD lizards basked in direct sunlight (full sun or partial sun). In each of these, lizards were positioned perpendicular to sun rays with dorsum facing the sun and the venter applied to the ground. As Tb’s increased lizards were found in the open habitat (still in direct sunlight) but with their heads facing the sun and their venters held above the ground. Above Tb’s of 36°C, lizards shuttled between partial sun, edge or full shade and the open habitats. The highest mean Tb’s were taken from arboreal individuals in second rocky ridge. Lizards of the coastal sand dunes were never observed in arboreal microhabitat even though this microhabitat was present.
| Table 1: | Comparison for air temperature (Ta), soil temperature (Ts) and body temperature (Tb) of Acanthodactylus boskianus at coastal sand dunes (CSD) and second rocky ridge (SRR). Sample sizes are presented in parentheses |
 | |
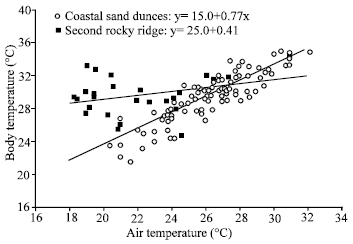 | |
| Fig. 3: | Relationship between body temperature for Aconthodactylus boskdonus from cosstal sand danes (cricles, deshed line) and second rocky ridge (squares, full line). Body temperature was positively correlated with air temerature at both sites. The slopes of the regression are significantly differnt |
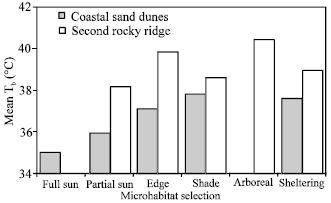 | |
| Fig. 4: | Mean body temperatures (Tb) organized by microhabitat of A. boskiamus at coastal sand dunes and second rocky ridge. Numbers above bars are sample sizes |
Generally, lizards basked in the partial sun and edge in the early morning while shade, arboreal and sheltering microhabitats was used increasingly during the late morning and early afternoon hours (Fig. 5b). Individuals used Scorzonera alexandrina exclusively for arboreal perches in SRR and the full sun microhabitat was not present at this region.
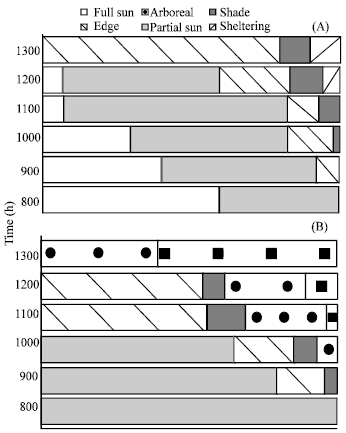 | |
| Fig. 5: | Proportion of lizard microhabitats selected separated by time of day (h) for A) coastal sand dunes and B) second rocky ridge |
DISCUSSION
Most terrestrial reptiles behaviourally regulate body temperature during periods of aboveground activity by exploiting spatial and temporal variations in microclimates. The body temperature of reptiles depends on both the magnitude of temperature variation present in the environment and the ability of these animals to regulate heat exchange with the environment. Variation in body temperature (Tb) and thermoregulatory behaviour within species are attributable to differences in the thermal environment via changing seasons, microhabitat selection and altitude (Smith et al., 1993; Smith and Ballinger, 1994; Hussein, 1998; O’Connor, 1999; Holtzclaw, 2001). However, no studies have shown differences in these thermal relationships across populations at similar elevations and seasons. In present study the two populations of A. boskianus inhabiting thermally divergent environments, separated by only 3.2 km at similar altitudes, showed differences in Tb’s and behavioural thermoregulation.
Thermoregulation in lizards may be evaluated by examining the variance in Tb’s of the field active individuals (Hertz et al., 1993). At CSD, Tb’s were significantly lower than Tb’s of lizards at SRR. This suggests that lizards at CSD cannot regulate body temperatures as effectively as individuals at SRR. The environmental properties (biotic and abiotic) of coastal sand dunes may hamper significant increases in Tb’s. For example, Ts’s at CSD were lower than Tb’s. Consequently, lizards at CSD may be mostly restricted to direct heat uptake from solar radiation (heliothermically basking). In contrast, Ts’s at SRR were higher than Tb’s, suggesting that relatively more heat energy is stored in the substrate during the day at SRR than at CSD. It is not surprising that the majority of SRR lizards found perching within canopy of shrubs or sheltering under plants due to high Ts’s.
All work to date has shown the ability of reptiles to thermoregulate by behavioural exploitation of thermal environment. A regression of Tb on Ta is another method of evaluating temperature regulation in lizards (Avery, 1982; Arnold and Peterson, 1989; Hussein, 1992; Bauwens et al., 1996). The slope from this linear relationship can indirectly identify the extent of thermoregulation. A slope near 0 suggests more precise regulation, whereas a slope near 1 suggests that body temperature depend upon ambient temperature. The slope calculated for A. boskianus population at CSD is high (0.77) compared with slope calculated for population at SRR (0.41), as well as compared to the slopes for other desert lizards. For example, reported slopes for Sceloporus jarrovi were 0.40 (Middendorf and Simon, 1988). Smith et al. (1993) calculated a slope of 0.23 for Sceloporus scalaris and the slope for Urosaurus ornatus was recorded as 0.30 (Smith and Ballinger, 1995). Of those lizards studied, it appears that most species are relatively precise thermoregulator, whereas A. boskianus shows interpopulational differences in control over thermoregulation.
Most studies to date have emphasized the success of lizards in maintaining relatively constant, often high, Tb and consequences of this stenothermy for organism performance. Body temperature regulation via behavioural adjustments allows lizards to maintain fairly constant Tb’s in an environment. Lizards of thermally heterogeneous environments have the potential to increase and maintain optimal Tb’s that maximize performance, such as locomotion (Walsberg, 1986; Hussein, 1992; Hertz et al., 1993; Firth and Belan, 1998; El-Masry and Hussein, 2001). However, when the thermal conditions are relatively homogeneous, lizards could experience less that adequate thermal condition (Huey and Bennett, 1987) or remain inactive for extended periods of time (Grant and Dunham, 1988). In present study, the microhabitat use, an index of behavioural thermoregulation, was different between lizards at CSD and SRR and that microhabitat influenced Tb’s at CSD, but not the Tb’s at SRR. This suggests that the CSD habitat is more thermally homogeneous than the environment at SRR. Moreover, the lizards at CSD appear to thermoregulate in a limited sense by increasing Tb’s throughout the morning and remaining active into the afternoon. In contrast, lizards at SRR appear to thermoregulate more carefully by obtaining activity Tb’s at a relatively faster rate in the morning then behaviourally regulate Tb’s throughout the rest of the afternoon by shuttling between thermally appropriate microhabitats.
In SRR, A. boskianus restricts its perching and sheltering to its preferred microhabitat. The majority of lizards were found sheltering under the branches of Scorzonera alexandrina. The consequences of this extreme restriction may indicate that this plant act as special microhabitat offer superior opportunities for thermoregulation compared with other available vegetation (Avery, 1982; Castilla and Bauwens, 1991). In addition, the stay of lizards under certain plant cover may have many other advantages such as providing safe hiding places inaccessible to most potential predators. Also, most of the lizards prey items have been encountered among the plant branches suggesting that lizards may forage for food while they remain under cover.
Generally, the data presented in this study suggest that Tb’s, thermoregulation and microhabitat selection vary between populations of A. boskianus found in diverse thermal environment due to environmental temperature differences between sites which may explain many of the observed thermal differences in this species of lizards inhabiting the same elevation.
REFERENCES
- Arnold, S.J. and C.R. Peterson, 1989. A test for temperature effects on the ontogeny of shape in the garter snake, Thamnophis sirtalis. Physiol. Zool., 62: 1316-1333.
Direct Link - Bauwens, D., P.E. Hertz and A.M. Castilla, 1996. Thermoregulation in a lacertid lizard: The relative contributions of distinct behavioral mechanisms. Ecology, 77: 1818-1830.
CrossRefDirect Link - El-Masry, A.A. and H.K. Hussein, 2001. Thermal relations, metabolism and winter dormancy of the sand lizard, Acanthodactylus boskianus. Pak. J. Biol. Sci., 4: 492-497.
CrossRefDirect Link - Huey, R.B. and R.D. Stevenson, 1979. Integrating thermal physiology and ecology of ectotherms: A discussion of approaches. Am. Zool., 19: 357-366.
CrossRefDirect Link - Huey, R.B., E.R. Pianka and J.A. Hoffman, 1977. Seasonal variation in thermoregulatory behavior and body temperature of diurnal Kalahari lizards. Ecology, 58: 1066-1075.
CrossRefDirect Link - Walsberg, G.E., 1986. Thermal consequences of roost-site selection: The relative importance of three modes of heat conservation. Auk, 103: 1-7.
Direct Link