
A. A. Ali
Department of Botany, Faculty of Science, University of Zagazig, Zagazig, Sharkia, Egypt
C. L. Mulchi
University of Maryland, Department of NRSLA, College Park, MD 20742, USA
I.A. Hassan
Department of Botany, Faculty of Science, University of Alexanderia, Alexanderia, Egypt
Y. A. El-Zawahry
Department of Botany, Faculty of Science, University of Zagazig, Zagazig, Sharkia, Egypt
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 7 | Page No.: 798-801







ABSTRACT
Open-top chambers (OTC’s) were conducted to determine interactive effects of atmospheric CO2 and O3 air pollution on photosynthetic responses in wheat. The plants were grown full-season in these chambers supplied with charcoal-filtered air (CF) as control, CF + 155μL CO2 L–1 as high CO2, non-filtered air (NF) + 40nL O3 L–1 as high O3 and NF + 155μL CO2 L–1 + 40nL O3 L–1 as combined high CO2 and high O3. Photosynthetic rates (Pn) were measured three times during vegetative and reproductive stages with portable gas exchange system (LI-COR 6200). In general, Pn rates were stimulated by high CO2 and reduced by high O3 but in some cases, combined high CO2 and high O3 increased the Pn rates. The data showed continuous increases in Pn rates during pre-flowering and early seed formation and drops during late seed formation stage. This study supports that the concentration of CO2 at 5 ± 500μL L–1 treatment had a protective role against adverse impacts of O3 exposure at concentration 5 ± 60nL L–1 treatments.
PDF Abstract XML References Citation
How to cite this article
A. A. Ali, C. L. Mulchi, I.A. Hassan and Y. A. El-Zawahry, 2002. Reversibility of Photosynthetic Inhibition After Long-term Exposure of Wheat Plants (Triticum aestivum L. Cvs. Sesquehanna and Gore) to Elevated Levels of Ozone. Pakistan Journal of Biological Sciences, 5: 798-801.
DOI: 10.3923/pjbs.2002.798.801
URL: https://scialert.net/abstract/?doi=pjbs.2002.798.801
DOI: 10.3923/pjbs.2002.798.801
URL: https://scialert.net/abstract/?doi=pjbs.2002.798.801
INTRODUCTION
Many models designated to assess the future impacts of complex phenomenon as climate change on crop growth and productivity have typically considered the beneficial effects of rising atmospheric CO2 concentration on plants, while largely neglecting the potential negative impacts of phytotoxic gases such as O3 (Adams et al., 1990; Stockle et al., 1992).
Carbondioxide and ozone are commonly referred to as “greenhouse” gases because of their abilities to absorb infrared radiation being emitted by earth resulting in the re-emission of this energy into the troposphere. Tropospheric O3 concentrations vary widely over the earth’s surface and are influenced by a number of factors including: 1) localized meteorological parameters; 2) levels of solar radiation as influenced by latitude; 3) proximity to carbon emission centers; 4) background levels of O3 precursors including VOC’s and other reactive organic compounds in the air mass; and 5) long range transport processes (Krupa and Kickert, 1989; Barnes and Wellburn, 1999). Readers are encouraged to consult reviews regarding more in-depth discussions on the processes influencing CO2 and O3 levels in the atmosphere.
Both CO2 and O3 have fundamental effects on CO2 exchange by plants. The CO2 uptake (photosynthesis) may be affected, with the net C gain allocated to different plant processes. In general, elevated CO2 enhances the rate of CO2 uptake for C3 plants (Bowes, 1993). However, due to acclimation in some cases the rates of photosynthesis at elevated CO2 may not be as high as expected based on ambient CO2 plants (Amthor, 1997).
The most famous physiological parameters affected by O3 is photosynthesis (Pn) (Saxe, 1991). Studies of photosynthesis are important to understand the effect of air pollutants including stress on crop growth and production (Miller, 1988). Lehnherr et al. (1988) noted that Pn rates were reduced mainly when they exposed wheat plants during senescence to several concentrations of O3 such as 15, 30, 70 and 100 nmol O3 mol–1 full season for 8 h day–1. Soybean plants exposed to chronic O3 doses also had reduced leaf Pn rates with increased O3 concentrations (Reich et al., 1986; Mulchi et al., 1992). Chernikova (1998) found only minimal responses in Pn to increased O3 exposures for soybean cultivars during pre-flowering; however, during podfill, Pn rates declined in a linear fashion over the range of O3 levels 27 to 60 nmol O3 mol–1.
The reduction in Pn rates of bean plants during chronic O3 exposure observed early in the growing season (0 to 44 days after emergence), but recovered over night. Later in the season, (i.e. 60 days after emergence), photosynthetic capacity and stomatal conductance gradually decreased as the severity of O3 injury increased (Sanders et al., 1992).
Photosynthesis is stimulated in C3 species under increased intercellular CO2 concentration due to increased carboxylation of Rubisco (Bowes, 1991). However, sensitivity to high CO2 concentrations might be reduced over time due to saturated CO2 binding to Rubisco and limited regeneration of ribulose 1,5-bisphosphate (RuBP) and/or inorganic phosphorus (Pi) (Stitt, 1991).
Few studies investigated the combined effects of O3 and CO2 on photosynthesis, with some suggest. McKee et al. (1997) reported that Triticum aestivum had a large increase in Rubisco activity with elevated CO2 at high but not low O3, and had little change in conversion activity with elevated CO2 at either O3 level. Similarly, Kull et al. (1996) indicated slightly greater increase in photosynthetic rate for Populus tremuloides with elevated CO2 at low compared to high O3, and a greater decrease in photosynthetic rate with elevated O3 at high compared to low CO2.
The main objective of this study was to investigate the possible interactive effects of CO2 and O3 on one of the biochemical responses of wheat plants grown in open-top chambers.
MATERIALS AND METHODS
Design and treatments: All measurements were performed in 16 open-top chambers (OTC’s) as described by Heagle et al. (1973) during the period of early March to late June. The studies were carried out at the South farm, United States Department of Agriculture (USDA)-Beltsville Agriculture Research Center (BARC) near Washington, DC, USA. The soil was amended with fertilizers at the rates recommended for wheat plants and pre- or post-emergence herbicide was applied to control weeds. Sprinkler irrigation units were utilized to maintain soil moisture levels near field capacity. Wheat plants were grown in plots equipped 3-m diameter x 2.5-m high open top chambers. Two replicates with 8 chamber treatments per replicate i.e. 4 air quality treatments, 2 moisture regimes and 2 cultivars. Air quality treatments are charcoal-filtered air (CF) as a control, CF + 155μL CO2 L–1 (high CO2), non-filtered air (NF) + 40nL O3 L–1 (high O3) and NF + 155μL CO2 L–1 + 40nL O3 L–1 (high CO2 + high O3). The two moisture regimes are well-watered and restricted water conditions. Wheat cultivars are gore and sesquehanna. The treatments were arranged in complete factorial design 4 x 2 x 2. The CO2 treatments were applied from a bulk tank and injected to the OTC’s box fans 18h day–1 (0400-2200 h EST) at rates necessary to raise the ambient CO2 levels in the high CO2 treatments by 155 μL CO2 L–1. The O3 was generated from cylinder O2 using a Griffin model FTCIA O3 generator (Griffin Technique Co., NJ) and introduced into the blowers of the OTC between the particulate filter and fan.
Leaf photosynthetic rates (μmol m–2 s–1): Photosynthetic rate under different treatments was measured from all chambers with a portable closed gas exchange system (Model LI-6200 primer, LI-COR, Lincolin, NE). Photosynthesis measurements where taken three times during vegetative and reproductive growth stages of wheat plants: one before flowering (pre-flowering); the second one at early seed formation; and the third one after seed formation on expanded leaves of the upper canopy under direct sun light during middle of the day. The Pn rates were always performed on three plants per cultivar per chamber between 11.00 to 14.00 h EST.
Wheat yield: It was calculated for harvested plants after reaching the maturity. By removing seeds from each plant, these seeds were left until reaching the constant weight. The collected seeds were weighed for each meter square and expressed as g/m2.
Statistical analysis: Data was analyzed using analysis of variance (ANOVA) procedures. Treatment means were separated using least significant difference (LSD) comparisons where ANOVA’s f-tests were significant (Gomez and Gomez, 1984). Significance was tested at P ≤ 0.05 level. The software developed by the Statistical Analysis System (SAS Institute, 1990) was used to perform all analysis.
RESULTS AND DISCUSSION
The measurements of 4-hours temperature for the air surrounding wheat plants inside the OTC’s and soil temperature are shown in Fig. 1 and 2.
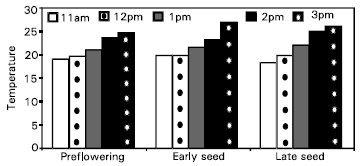 | |
| Fig. 1: | Mean values of air temperature (°C) in OTC’s of wheat at different growth stages under four air quality treatments and two soil moisture regimes. |
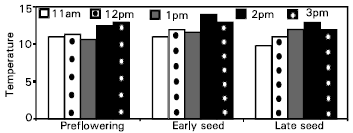 | |
| Fig. 2: | Mean values of temperature (°C) in OTC’s of wheat at different growth stages under four air quality treatments and two soil moisture regimes. |
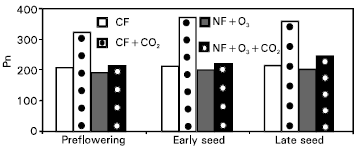 | |
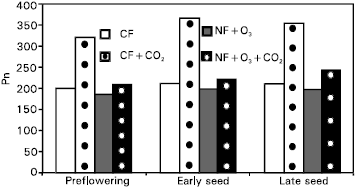
| Fig. 3: | Photosynthetic rates (μmole m–2 s–1) of wheat leaves at different growth stages under four air quality treatments and wet soil. |
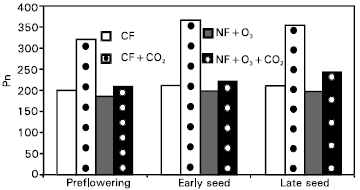 | |
| Fig. 4: | Photosynthetic rates (μmol m–2 s–1) of wheat leaves at different growth stages under four air quality treatments and dry soil. |
 | |
| Fig. 5: | Photosynthetic rates (μmol m–2 s–1) of wheat leaves for its cultivars at different growth stages under treatments and two soil moisture regimes. |
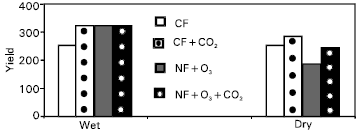 | |
| Fig. 6: | Mean values wheat yield (g/m2) under four air quality treatments and soil moisture regimes. |
Gradual increase occurred in air temperature for all growth stages till reaching 2pm then slight decrease at 3pm. The soil temperature for all growth stages was not taking clear manner. Da Costa et al. (1986) suggested that CO2 release or uptake of a full crop in the field could be predicted with reasonable accuracy from knowledge of the air temperature and soil moisture content.
Effects of CO2 and O3 on photosynthesis Pn leaf measurements in winter at three growth stages are summarized in Fig. 3, 4 and 5, respectively. Also, these values were higher under wet conditions in comparing with that at dry treatments. The Pn rate values were increased gradually starting from pre-flowering growth stage but decreased slightly through the stage of late seed formation. Wheat cultivars exhibited big difference in Pn values for gore and sesquehanna (Fig. 5). The data showed that wheat gore was more sensitive to treatments than sesquehanna.
In general, wheat plants grown under enriched CO2 had higher Pn rates than plants grown under ambient CO2 (Fig. 3 and 4). In few cases, the CO2 enrichment had no significant effect on Pn rates under dry conditions in comparison with combination between CO2 and O3 (Fig. 6), which may be attributed to the fact that plants were fully acclimated to CO2 enriched environment (Allen, 1990). During the later stages of vegetative growth, perhaps CO2 was no longer a limiting growth factor. During this period, sink capacity becomes limited and Pn rates were likely reduced due to a possible accumulation of starch grains in the chloroplast which triggers feedback mechanisms that inhibit photosynthesis (Stitt, 1991). However, during the late and early handing process significant CO2 effects were again observed which might be attributed to the greater demand for carbohydrates in response to increased sink capacity by the plant (Woodword et al., 1991; Stitt, 1991). Later in the season, when plants were in the ripening stage, no significant difference was found but plants grown under enriched CO2 presented higher Pn rates for each of the two last readings (early seed formation and late seed formation), respectively. This can be attributed to a small delay in leaf senescence observed for plants grown under enriched CO2. Barnes et al. (1995) reported that interactions between carbon assimilation, carbohydrate status and chemical composition (nutrient status) may dictate the manner in which plants respond to rising CO2 concentrations, and governed the ability of the plant to sustain its positive response to CO2 enrichment.
Chronic O3 exposure tended to reduce Pn rates (Fig. 3 and 4) but the results were significant primarily when combined over dates. Significant lower Pn rates were observed at pre-flowering growth stage and at the early and late ripening stages, respectively. The overall reduction in Pn rates observed during ripening can be attributed to the early senescence noted for plant leaves exposed to chronic O3 stress (Heck, 1990). The overall effects of O3 on Pn rates were significant with high-O3 treatments reducing Pn rates. Since measurements were taken preferentially on green leaves avoiding measurements on senesced leaf tissue the Pn rates on the high-O3 treatment plants were likely over estimated.
The overall treatment effects on Pn showed that CO2 enrichment prevented O3 exposed plants from reduction in Pn rates below that observed from the control (Fig. 3 and 4). Photosynthetic rates for the combined enriched CO2 level with atmospheric O3 were significant under wet treatments for all growth stages compared with dry ones, which most probably can be attributed to the reduced temperature. The Pn rate values were significant for early and late seed formation stages in comparison with high O3 treatments.
The sensitivity of Pn to air pollutants is affected by genotypes (Reich and Amundson, 1985; Miller, 1987), development stage (Lehnherr et al., 1988), and various environmental factors such as light intensity, ambient CO2 levels, nutrient status and water availability (Darral, 1989; Runeckles, 1992). The reduction in Pn rates of wheat plants during senescence are associated with increased stomatal conductance and decreased in various components of the photosynthetic apparatus such as chlorophyll concentration, soluble protein, adenylates, RuBP regeneration, and Rubisco (ribulose 1,5-bisphosphate carboxylase/oxygenase) activity. Farage et al. (1991) concluded that the first inhibitory effect of O3 on Pn is the loss of carboxylation efficiency (i.e. CO2 uptake/ internal leaf CO2 concentration) due to decreased activity of Rubisco.
The effects of CO2 and O3 on wheat yield quality are illustrated in Fig. 6. Both gases were found to cause large changes in grains quality of wheat. The elevated CO2 significantly increased the wheat yield while high O3 reduced the quality. The interactive effect of high CO2 and high O3 exhibited significant increase in grain quality of wheat in comparison with atmospheric O3 treatments. Reduction in grain yield of wheat in response to O3 levels induced stress was attributed to reduced Pn due to early senescence and reduced capacity of plants to provide photosynthetic assimilate to grains (Amundson et al., 1987; Lehnherr et al., 1987; Miller, 1987). Also, the decrease in photosynthetic rates paralleled the content of Rubisco (Lehnherr et al., 1987) in response to premature senescence of the flag leaf triggered by O3-induced stress (Amundson et al., 1987; Lehnherr et al., 1987). Kull et al. (1996) explained the impact of O3 on plants by depression of photosynthetic activity and the accelerated senescence of leaves. Moreover, a genotype considered tolerant with normal CO2 levels appears to have decreased O3 tolerance with elevated CO2.
Electron microscope examination of O3 injury revealed that the considerable disruption including tonoplast rupture which may have caused a complete disruption of the osmotic balance within the cell inactivating the photosynthetic process (Sanders et al., 1992).
There was no evidence of an interactive effect as elevated CO2 increased and high O3 decreased the photosynthesis to a similar extent at both levels of the other gas on Triticum aestivum (Rudorff et al., 1996). Thus, the response to O3 and CO2 likely was additive in these species, with the response to O3 and CO2 at least partially canceling each other out. Picea abies showed little effect of elevated CO2 on photosynthesis at either O3 levels or an increase in photosynthesis at both CO2 levels (Lippert et al., 1997). The wheat plant (C3) species exhibited significant response to the atmospheric CO2 and O3 treatments. The mechanism(s) involved concerning a possible protective role of CO2 against O3 are largely unknown. In addition, the results from present study support a hypothesis that the added CO2 somehow protects the wheat’s ability to partition the photosynthates to developing sinks as grains. The collected data supported that the elevated CO2 treatments can increase the productivity of wheat plant.
REFERENCES
- Adams, R.M., C. Rosenzweig, R.M. Peart, J.T. Ritchie and B.A. McCarl et al., 1990. Global climate change and US agriculture. Nature, 345: 219-224.
CrossRefDirect Link - Allen, L.H., 1990. Plant responses to rising carbon dioxide and potential interactions with air pollutants. J. Environ., Qual., 19: 15-34.
Direct Link - Amthor, J.S., 1997. Plant Respiratory Responses to Elevated Carbon Dioxide Partial Pressure. In: Advances in Carbon Dioxide Effects Research. Allen, L.H., M.B. Kirkham, D.M. Olszyk and C.E. Whitman (Eds.). ASA, Madison, WI., USA., pp: 35-77.
Direct Link - Amundson, R.G., R.J. Kohut, A.W. Schoettle, R.M. Raba and P.B. Reih, 1987. Correlative reductions in whole-plant photosynthesis and yield of winter wheat caused by ozone. Physiol. Biochem., 77: 75-79.
Direct Link - Barnes, J.D., T. Pfirrmann, K. Steiner, C. Lutz, U. Busch, H. Kuchenhoff and H.D. Payer, 1995. Effects of elevated CO2, Elevated O3 and potassium deficiency on Norway spruce [icea abies (L.) Karst]: Seasonal changes in photosynthesis and non-structural carbohydrate content. Plant Cell Environ., 18: 1345-1357.
- Bowes, G., 1993. Facingthe inevitable: Plants and increasing atmosphere CO2. Ann. Rev. Plant Physiol. Plant Mol. Biol., 44: 309-332.
CrossRefDirect Link - Farage, P.K., S.P. Long, E.G. Lechner and N.R. Baker, 1991. The sequence of change within the photosynthetic apparatus of wheat following short-term exposure of ozone. Plant Physiol., 95: 529-535.
Direct Link - Gomez, K.A. and A.A. Gomes, 1984. Statistical Procedures for Agriculture Research. 2nd Edn., John Wiley and Sons, New York, ISBN: 9780471870920, Pages: 680.
Direct Link - Heagle, A.S., D.E. Body and W.W. Hech, 1973. An open-top field chamber to assess the impact of air pollution on plants. J. Environ. Qual., 20: 562-570.
Direct Link - Kull, O., A. Sober, M.D. Coleman, R.E. Dickson, J.G. Isebrands, Z. Gagnon and D.G. Karnosky, 1996. Photosynthetic responses of aspen clones to simultaneous exposures of ozone and CO2. Can. J. For. Res., 26: 639-648.
CrossRefDirect Link - Lehnherr, B., F. Machler, A. Grandjean and J. Fuhrer, 1988. The regulation of photosynthesis in leaves of field grown spring wheat (Triticum aestivum L. cv. Albis) at different levels of O3 in ambient air. Plant Physiol., 88: 1115-1119.
Direct Link - Lippert, M., K. Steiner, T. Pfirrmann and H.D. Payer, 1997. Assessing the impact of elevated O3 and CO2 on gas exchange characteristics of differently K supplied clonal Norway spruce trees during exposure and the following season. Trees Struct. Funct., 11: 306-315.
Direct Link - McKee, I.F., J.F. Bullimore and S.P. Long, 1997. Will elevated co2 concentrations protect the yield of wheat from o3 damage? Plant Cell Environ., 20: 77-84.
CrossRefDirect Link - Mulchi, C.L., L. Slaugter, M. Saleem, E.H. Lee, R. Pausch and R. Rowland, 1992. Growth and physiological and physiological characteristics of soybean in open-top chambers in response to ozone and increased atmospheric CO2. Agric. Ecosyst. Environ., 38: 107-118.
CrossRefDirect Link - Reich, P.B. and R.G. Amundson, 1985. Ambient levels of ozone reduce net photosynthesis in tree and crop species. Science, 230: 566-570.
CrossRef - Rudorff, B.F.T., C.L. Mulchi, E.H. Lee, R.A. Rowland and R. Pausch, 1996. Photosynthetic characteristic in wheat (Triticum aestivum) exposed to elevated O3 and CO2. Crop Sci., 36: 1247-1251.
Direct Link - Sanders, G.E., J.J. Colls and G. Clark, 1992. Physiological changes in Phaseolus vulgaris in response to long-term ozone exposure. Ann. Bot., 69: 123-133.
Direct Link - Stockle, C.O., P.T. Dyke, J.R. Williams, C.A. Jones and N.J. Rosenberg, 1992. A method for estimating the direct and climate effects of rising atmosphere carbon dioxide on growth and yield of crops. II. Sensitivity analysis at three sites in the Midwestern USA. Agric. Syst., 38: 239-256.
Direct Link - Woodword, F.I., G.B. Thompson and I.F. Mckee, 1991. The effects of elevated concentration of CO2 on individual plants, populations, communities and ecosystems. Ann. Bot., 67: 23-38.
Direct Link