
Shazia Afrasayab
Department of Botany, University of the Punjab, Quaid-e-Azam Campus, Lahore-54590, Pakistan
Azra Yasmin
Department of Botany, University of the Punjab, Quaid-e-Azam Campus, Lahore-54590, Pakistan
Shahida Hasnain
Department of Botany, University of the Punjab, Quaid-e-Azam Campus, Lahore-54590, Pakistan
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 7 | Page No.: 792-797







ABSTRACT
Among six Hg-resistant isolates (SHg-7, SHg-8, SHg-9, SHg-10, SHg-11, SHg-12), SHg-8 and SHg-12 could tolerate 450 μg ml–1 of HgCl2 in L-agar medium, whereas rest of the strains could tolerate 500 μg ml–1 of HgCl2 in this medium were Gram-negative rods (SHg-7, SHg-8, SHg-9, SHg-10, SHg-11) and SHg-12 were Gram-positive rods. SHg-11 & SHg-12 were strictly aerobic, remaining were facultatively aerobic. On the basis of morphological and biochemical characterization isolates SHg-7, SHg-8, SHg-9, SHg-10 could be affiliated with family Vibrionaceae and SHg-11 shared characters with Pseudomonadaceae, while the affinities of SHg-12 remained uncertain. All of them had a single plasmid band except SHg-11, but these plasmids were non-conjugative. They had broad pH range (5-11) with pH optima either at 6 or 7. Their optimal growth temperature was 32°C (SHg-9, SHg-10, SHg-11) and 37°C (SHg-7, SHg-8, SHg-12). All of them were sensitive to Sm, Km and Tc, while they showed resistance against Ap and Cm. They could also tolerate other heavy metal salts.
PDF Abstract XML References Citation
How to cite this article
Shazia Afrasayab, Azra Yasmin and Shahida Hasnain, 2002. Characterization of Some Indigenous Mercury Resistant Bacteria from Polluted Environment. Pakistan Journal of Biological Sciences, 5: 792-797.
DOI: 10.3923/pjbs.2002.792.797
URL: https://scialert.net/abstract/?doi=pjbs.2002.792.797
DOI: 10.3923/pjbs.2002.792.797
URL: https://scialert.net/abstract/?doi=pjbs.2002.792.797
INTRODUCTION
The haphazard growth of industries, exploding population and agricultural revolutions have affected greatly the man's physical environment. Beside drugs, antibiotics and radioactive substances, industrial wastes contain heavy metals, which are mutagenic, carcinogenic and teratogenic. Many of the heavy metals have deleterious effects on biological life (Blaudez et al., 2000; Seeber et al., 2002; Akermoun et al., 2002). Mercury is one of the major pollutant in the environment which is highly toxic. Mercury has mutagenic, growth inhibitory and toxic effects and causes many serious human disease syndromes (Meyer-Baron et al., 2002).
Besides human beings, plants and animals Hg also affects bacterial growth. However some bacteria could grow in the mercury rich environment (Iwahori et al., 2000). Metal resistant systems are well known in many bacterial types. Bacteria have developed a variety of means at cellular and molecular level to overcome the stresses in their external environment (Hobman and Brown, 1997; Silver, 1998). Biomethylation is one of the defense mechanism which helps bacteria to transform ionic mercury to highly neurotoxic compound, methyl-mercury and further reduction of this compound to Hg°, a volatile element which possess less environmental risk (King et al., 2000). Sulfate reducing bacteria (SRB) are the major biological contributors of methyl mercury formation in sediments (King et al., 2000; Benoit et al., 2001). Bacterial resistances to heavy metals are encoded by chromosomal genes (Gupta et al., 1999) or plasmid mediated (Ravel et al., 2000).
The uptake of heavy metals, present in the industrial wastes, and the detoxification of metal ions by bacteria provide an additional mechanism of environmental protection. In the present work the isolation of Hg-resistant bacteria from the polluted water samples and their characterization is reported. The bacteria were also studied for the presence of plasmid and conjugal transfer.
MATERIALS AND METHODS
Three samples of polluted water were collected for the isolation of Hg-resistant bacteria. One sample was taken from the main drain of Township (Lahore) and two from polluted area around Sheikhupura road.
For the isolation of Hg-resistant bacteria, polluted water samples and their dilutions were plated on L-agar plates supplemented with 100 μg ml–1 of HgCl2. Purified colonies of the isolates were characterized morphologically, biochemically, physiologically and genetically. Different biochemical tests were performed (Gerhardt et al., 1994) for characterization of Hg-resistant bacteria. Cytochrome oxidase and phenylalanine deaminase tests were accomplished by using CO and PD strips (DESTO Laboratories Karachi). Spore forming ability was checked (Moir, 1981).
Resistance of strains to different antibiotics [streptomycin (Sm-500 μg ml–1), kanamycin (Km-40 μg ml–1), ampicillin (Ap-300 μg ml–1), tetracycline (Tc-25 μg ml–1 ), chloramphenicol (Cm-5 μg ml–1)] and different concentrations of metallic salts such as NiSO4, CoCl2, MnSO4, K2CrO4, Pb(NO3)2, CdCl2, ZnSO4 and CuSO4 were also checked. The effects of environmental factors like time period (0-48 hours), temperature (24, 28, 32, 37 and 42°C) and pH (5-11) were also studied on the growth of Hg-resistant bacteria. The mercury uptake of Hg-resistant isolates was determined by the method of Rand et al. (1979). Bacterial strains were screened for the presence of plasmid by the gel electrophoresis of total cell lysate (Thomas, 1984). Conjugation experiments were performed for the characterization of plasmid using broth mating techniques (Willetts, 1988). Escherichia coli strains HB101 [leu pro lac gal str his recA hsdR hsdM F–] and CSR 603 [recA1 phr1 derivative of AB1886 (thr-1 leu6 lacY galK ara-14 xyl-15 mtl-1 proA his-4 str-31 tsx-33 sup-37 uvxA6)] were used as recipients. Transconjugants were scored on L-agar plates supplemented with 50 μg ml–1 HgCl2 and 500 μg ml–1 Sm, at 37°C.
RESULTS
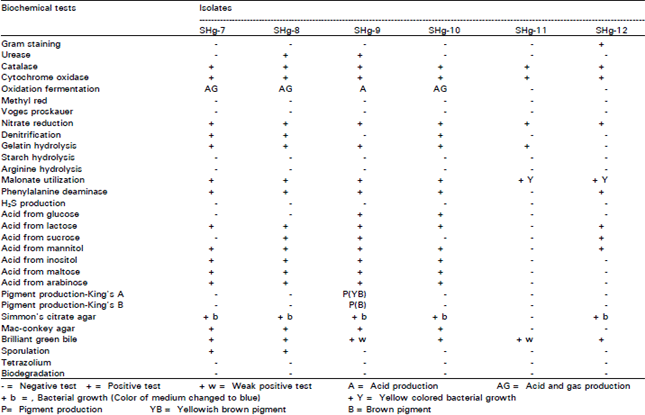
Strains of SHg-7, SHg-8, SHg-9, SHg-10, SHg-11 and SHg-12 were grown on L-agar plates supplemented with 100-500 μg ml–1 of HgCl2. The strains SHg-8 and SHg-12 could tolerate 450 μg ml–1 of HgCl2 in solid medium, while rest of the strains could resist 500 μg ml–1 of HgCl2 . These strains could resist 50-150 μg ml–1 of HgCl2 in the liquid medium (Fig. 1). All of them had off-white, circular colonies with entire margins. Cells of these strains were motile and non-capsulated. SHg-12 was gram +ve rods, whereas SHg-7, SHg-8, SHg-9, SHg-10 and SHg-11 were gram -ve rods. All the strains had catalase, urease and cytochrome oxidase enzymes. SHg-7, SHg-8, SHg-9 and SHg-10 were facultative anaerobes, while SHg-11 and SHg-12 were strictly aerobic bacteria (Table 1). SHg-7 and SHg-8 were spore-formers under stress conditions. All strains showed positive results for nitrate reduction, denitrification (except SHg-9, SHg-11, SHg-12), gelatin hydrolysis (except SHg-12), malonate utilization, phenylalanine deaminase (except SHg-11) tests. SHg-9 produced yellowish brown pigment on King's A medium supplemented with HgCl2 and brown pigment on King's B medium supplemented with HgCl2.
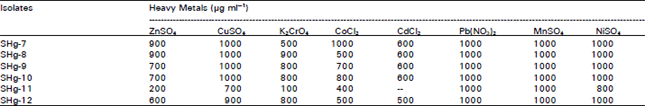
All the strains could resist Ap and Cm, while they were sensitive to Sm, Km and Tc (Table 2). They could bear 100-1000 μg ml–1 of salts of different metals in the medium (Table 3), except SHg-11, which was sensitive to CdCl2. pH range of isolates varied from 5-11. SHg-8, SHg-9 and SHg-10 could grow best at pH-6, which was reflected by increase in value of optical density at that pH. SHg-7, SHg-11 and SHg-12 showed maximum growth at pH-7.
| Table 1: | Biochemical characterization of Hg-resistant bacteria. |
 | |
| Table 2: | Antibiotics resistance profile of Hg-resistant bacteria |
 | |
| Table 3: | Heavy metal resistance of Hg-resistant bacteria |
 | |
At pH extremes bacterial density was drastically affected (Fig. 2). Optimal temperature for the growth of these strains varied. SHg-7 and SHg-12 showed maximum growth at 37°C, while SHg-9, SHg-10 and SHg-11 showed best growth at 32°C (Fig. 3), whereas optimum growth of SHg-8 was found both at 37 and 42°C. Hg-resistant strains showed variability in the growth pattern with time (Fig. 4). Generally lag phase persisted from less then 1-2 hours. SHg-10 and SHg-12 had optimum growth after 18 hours of incubation and after that time growth progressively decreased. SHg-7 and SHg-9 showed best growth after 10 hours, while SHg-8 and SHg-11 gave maximum growth after 16 hours of incubation at 37°C.
Gel-electrophoresis of total cell lysate showed that all the strains had a single plasmid band, except SHg-11, in which no plasmid band was discerned. No transconjugants were scored on mercury + Sm supplemented plates, when E. coli strains were used as recipient.
The mercury uptake ability of the Hg-resistant strains was checked by the digestion of bacterial pellet, washing of the pellet and supernatant of bacterial cultures. No mercury was detected in the washing of any strain. SHg-7 showed maximum uptake of Hg. Mercury was not detected in the pellet and washing of SHg-9 and SHg-11, hence they were unable to take-up mercury, but comparatively less amount of Hg in the supernatant revealed that the mercury was transformed into volatile form by the strains. SHg-8, SHg-10 and SHg-12 were also able to take-up Hg but less than SHg-7 (Fig. 5). The fact that amount of mercury supplied in the medium is not accounted for in pellet, washing and supernatant, demonstrate that inorganic mercury might be transforming to volatile form.
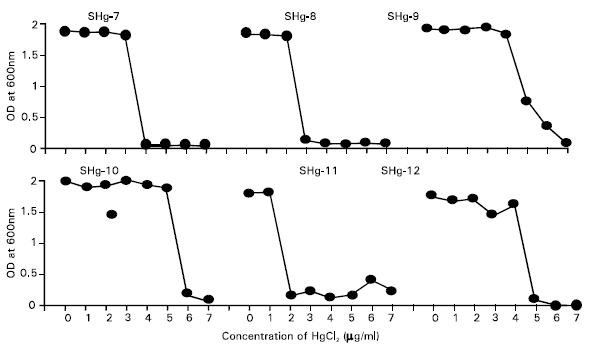 | |
| Fig. 1: | Growth of isolates in L-Broth supplemented with different concentrations of HgCl2 (in the figure along x-axis 0, 1, 2, 3, 4, 5, 6, 7, represents 0, 25, 50, 75, 100, 125, 150, 175 μg ml–1 of HgCl2 , respectively). |
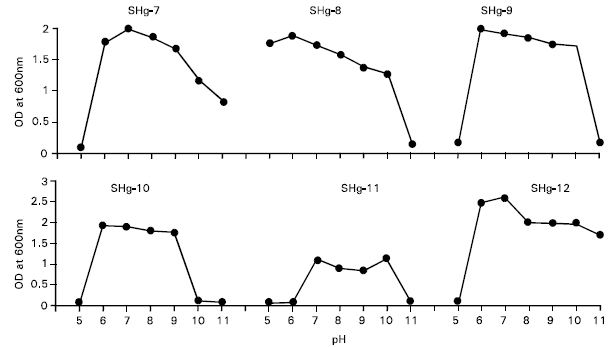 | |
| Fig. 2: | Growth of Hg-resistant bacteria under different pHs |
DISCUSSION
Six Hg-resistant strains SHg-7, SHg-8, SHg-9, SHg-10, SHg-11 and SHg-12 were isolated from three polluted water samples. These Hg-resistant strains were characterized morphologically, biochemically, physiologically and genetically. All the strains gave healthy growth at 100 μg ml–1 of HgCl2, but growth gradually decreased at higher concentrations of HgCl2 (450-500 μg ml–1). SHg-7, SHg-9, SHg-10 and SHg-11 could tolerate 500 μg ml–1 of HgCl2, but SHg-8 and SHg-12 could tolerate 450 μg ml–1 of HgCl2. The decreased growth at higher concentrations of HgCl2 revealed that Hg affects the growth of bacteria. Cell elongation, cell division and cell metabolism are affected under stress environmental conditions. In liquid medium the strains could tolerate 50-150 μg ml–1 of HgCl2. Thus the strains could resist relatively low concentrations of HgCl2 in the liquid medium as compared to solid medium. Perhaps this was due to the reason that in liquid medium, the bacterial cells are surrounded on all the sides with metal ions, while in solid medium, the Hg ions bound with the components of the agar medium. All the strains showed decreased growth in Hg supplemented medium while SHg-11 showed relatively better growth in Hg supplemented medium (25-50 μg ml–1 of HgCl2) as compared to medium where no Hg was present. This behaviour of strain may be due to genetic adaptation under extreme environment. Hg-resistant strains isolated by Zareen et al. (1997) could resist 200 μg ml–1 of HgCl2 in the solid medium. Hence the strains reported here could tolerate much higher level of mercury salt. Iwahori et al. (2000) reported Thiobacillus ferrooxidans SUG 2-2 strain isolated from hot spring water which was most resistant to mercury toxicity.
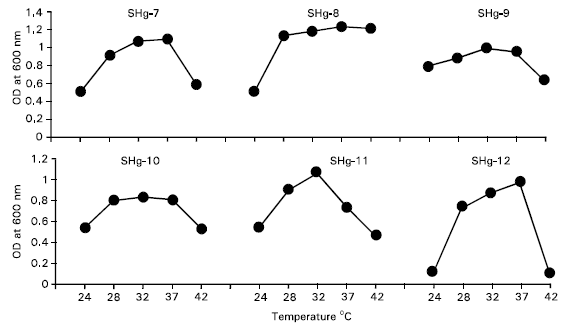 | |
| Fig. 3: | Growth of Hg-resistant bacteria at different temperatures. |
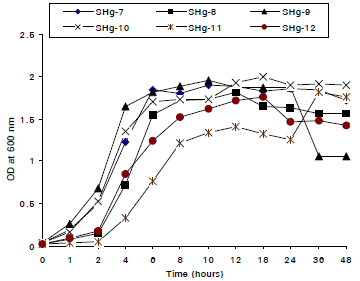 | |
| Fig. 4: | Growth curves of Hg-resistant bacteria (0-48 hours of incubation). |
All the strains were motile with off-white, circular colonies having entire margins. The strains were Gram-negative rods, except SHg-12, which were Gram-positive rods. All Gram negative and Gram positive and some Gram variable systems are resistant to a broad range of mercuric compounds including organomercurials like phenylmercuric acetate (PMA) (Hobman and Brown, 1997). Strains gave positive results for the presence of catalase, urease and cytochrome-oxidase enzymes. SHg-7, SHg-8, SHg-9 and SHg-10 were facultative anaerobes, while SHg-11 and SHg-12 were strictly aerobic bacteria. The experimental trends were similar to what were observed by Yasmin et al. (1997) and Afrasayab et al. (1998). Only SHg-7 and SHg-8 were spore formers under mercury stress conditions. The results of spore formation on Shaeffer's medium (with HgCl2 ), was indicated by the production of brown pigment in the center of the colonies. The same strains SHg-7 and SHg-8 gave positive results for spore staining under Hg stress. But they were non spore-former under unstressed conditions. SHg-7 and SHg-8 showed no response to tetrazolium dye, which means that they were unable to germinate.
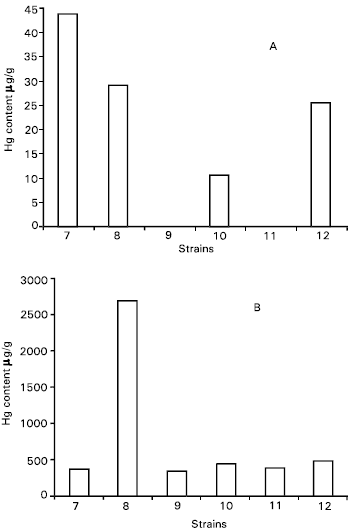 | |
| Fig. 5: | Amount of mercury present in the bacterial pellet (A) and in the supernatant (B) of mercury resistant bacteria |
It appears that some specific compound is synthesized/ accumulated in the walls of these strains which render them as spore like bodies but these spores/ bodies could not be germinated. It might be some adaptive/ protective mechanism under stress. On the basis of morphological and biochemical characterization isolates SHg-7, SHg-8, SHg-9, SHg-10 could be affiliated with family Vibrionaceae and SHg-11 shared characters with Pseudomonadaceae (Holt et al., 1994), while the affinities of SHg-12 remained uncertain.
Metal resistance test of Hg-resistant bacteria showed that all the strains had the ability to resist Pb(NO3)2 and MnSO4, but had different levels (100-1000 μg ml–1 ) of tolerance to other metallic salts i.e. CoCl2 , CdCl2 ,CuSO4 , ZnSO4 , NiSO4 and K2CrO4 (Table 3). The multiple metal resistance may be due to the presence of markers for these metallic salts on the same genetic determinant. Metal resistant systems are well known in many bacterial types. The genes governing these resistances are generally found on plasmids and encode resistances to toxic metal ions including Ag+, Cd+2, Co+2, CrO¯2, Cu+2, Hg+2, Ni+2, Pb+2 (Silver, 1998).
Hg-resistant strains were screened for the antibiotic resistances. All the strains were resistant to Ap and Cm but sensitive to Sm, Km and Tc. Results of McArthur and Tuckfield (2000) implied that increased antibiotic resistance is positively correlated with mercury concentration. Genes that code for antibiotic resistance and genes that code for metal resistance are often carried on the same plasmids or mobile genetic elements (Wireman et al., 1997). Multiple resistances of strains isolated from natural environment, to antibiotics is alarming as it reflect the frequent usage of antibiotics and their subsequent release in the environment.
pH range of isolates varied from 5-11. They showed weak growth at pH 5-11, but healthy growth between pH 6-10. Cells maintain relatively constant intracellular pH over a wide range of external values by pH homeostasis. Strain of SHg-8, SHg-9 and SHg-10 were acidophilic (pH-6). Strains SHg-7, SHg-11 and SHg-12 were neutrophilic and showed maximum growth at pH-7. Zareen et al. (1997) described Hg-resistant acidophilic strains from polluted waters. Temperature range of isolates varied from 24-42°C. Strain of SHg-9, SHg-10 and SHg-11 showed maximum growth at 37°C and SHg-7, SHg-8 and SHg-12 gave best growth at 32°C. All the strains gave weak growth at 24°C and 42°C. Temperature affects microbial cells by altering the rate of biochemical reactions and also by the induction of enzyme synthesis. Increased temperature causes change in membrane composition, stimulation in protein catabolism as well as cell death. Enzyme (α-galactosidase) of Thermus sp. strain T2 was most active at 75°C and most stable over a broad range of pH, 6-13 at 40°C (Ishiguro et al., 2001).
Gel-electrophoresis of total cell lysate revealed that all the strains harbour plasmid (except SHg-11). To probe whether mercury resistance in these strains is plasmid encoded and transferable, conjugation experiments were performed using E. coli K12 strains as recipient. No transconjugants were obtained in any case. These results reflect that either Hg-resistance was not determined by the plasmid present in these bacteria or that plasmid was not transferable. Alternatively host restriction mechanism or narrow host range of plasmid may be the reasons of failure in getting any transconjugants. Newby et al. (2000) introduced plasmid pJP4 from E. coli to indigenous soil population. Plasmid encoded mercury resistance and partial 2,4-Dichlorophenoxyacetic acid degradation and the transconjugants obtained also had the same ability. It has been shown that transfer of a catabolic plasmid from an introduced donor to indigenous microbial recipient may provide an environmentally stable host for the plasmid. Furthermore, remediation of contaminated soils may be enhanced as a result of such transfers.
Bacteria have the ability to combat heavy metals by a variety of means such as physical sequestration, exclusion, efflux reduced uptake, detoxification and synthesis of binding proteins (Nies et al., 2000; Zeroual et al., 2001). The resistance mechanism to Hg involves: the uptake of Hg+2 into the cytoplasm, reduction of ionic mercury Hg+2 by mercuric reductase, encoded by merA gene, to Hg° and release of Hg° from the cell by diffusion through cell membrane (Hobman and Brown, 1997). Mercury volatilization by Thiobacillus ferooxidans SUG 2-2 strain supplemented with 6 μM Hg+ was activated in the presence of Fe+2 (Iwahori et al., 2000) this strain could volatilize 86% of the total mercury added to the culture medium. Klebsiella pneumoniae, a mercury resistant strain, could effectively volatilize Hg and detoxify Hg compounds. The efficiency of clean up in waste water was 100% and volatilization rate was 89% (Zeroual et al., 2001). In the present case Hg resistant strains had the ability of uptake of Hg+2 into the cell except SHg-9 and SHg-11 (Fig. 5). Previously Hasnain and Co-workers (Yasmin et al., 1997; Afrasayab et al., 1998) have reported Hg resistant bacteria exhibiting high Hg uptake ability.
Mercury content was not detected in the pellet and washing of SHg-9 and SHg-11, which indicated that these isolates had changed the Hg to volatile form. Total amount of Hg supplied was 36896 μg l–1. SHg-7 showed the maximum uptake (43.75 μg/g fresh weight) and SHg-10 had least uptake (10.16 μg/g fresh weight). Absence of Hg in the washing of all these strains showed that mercury was not loosely or externally bound to the cells which has been observed in case of some other metals (Corcuera et al., 1993). A great deal of evidence indicates that exploitation of microbial based technologies for removal of toxic metals from industrial effluents and waste water provide an additional means of environmental protection. Hence bacteria are increasingly used for decontaminating polluted environments. The gene transfer from bacteria to plants has received much attention. Bizily et al. (2000) engineered transgenic Arabidopsis thaliana plants to express bacterial gene merA. Modified plants were able to grow on media containing 50-fold higher methyl-mercury concentration than wild type plants. The use of genetically engineered plants for environmental restoration is an emerging clean up technology. The present studies revealed that these strains had the ability to detoxify the mercury in the medium either by the uptake or by transforming it to volatile form or by utilizing both mechanisms. These bacteria can be used for environmental restoration in mercury polluted areas.
REFERENCES
- Bizily, S.P., C.L. Rugh and R.B. Meagher, 2000. Phytodetoxification of hazardous organomercurials by genetically engineered plants. Nat. Biotechnol., 18: 213-217.
CrossRef - Blaudez, D., B. Botton and M. Chalot, 2000. Effects of heavy metals on nitrogen uptake by Paxillus involutus and mycorrhizal birch seedlings. FEMS Microbiol. Ecol., 33: 61-67.
CrossRefDirect Link - Gerhardt, P., R.G.E. Murray, W.A. Wood and N.R. Krieg, 1994. Methods for General and Molecular Bacteriology. 2nd Edn., American Society for Microbiology, Washington, DC., ISBN-10: 1555810489.
Direct Link - McArthur, J.V. and R.C. Tuckfield, 2000. Spatial patterns in antibiotic resistance among stream bacteria: Effects of industrial pollution. Applied Environ. Microbiol., 66: 3722-3726.
Direct Link - Moir, A., 1981. Germination properties of a spore coat-defective mutant of Bacillus subtilis. J. Bacteriol., 146: 1106-1116.
PubMedDirect Link - Ravel, J., J. Diruggiero, F.T. Robb and R.T. Hill, 2000. Cloning and sequence analysis of the mercury resistance operon of Streptomyces sp. strain CHR28 reveals a novel putative second regulatory gene. J. Bacteriol., 182: 2345-2349.
CrossRefPubMedDirect Link - Silver, S., 1998. Genes for all metals: A bacterial view of the periodic table. J. Ind. Microbiol. Biotechnol., 20: 1-12.
Direct Link