
M.A. H. Chowdhury
Department of Agricultural Chemistry, Bangladesh Agricultural University, Mymensingh, 2202, Bangladesh
R. Begum
Department of Agricultural Chemistry, Bangladesh Agricultural University, Mymensingh, 2202, Bangladesh
M. R. Kabir
Department of Agricultural Chemistry, Bangladesh Agricultural University, Mymensingh, 2202, Bangladesh
H. M. Zakir
Department of Agricultural Chemistry, Bangladesh Agricultural University, Mymensingh, 2202, Bangladesh
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 7 | Page No.: 736-739







ABSTRACT
Microbial respiration (CO2 evolution),inorganic SO4-S and available P were monitored in a S deficient soil amended with dhaincha, cowdung and rice straw @ 1 g per 50 g soil during 30 days incubation. A basal dose of 250 μg N, 200 μg P and 250 μg K g -1 soil was applied. Dhaincha showed the highest decomposition rate and rice straw showed the lowest rate, while that of cowdung was intermediate. After 30 days of incubation, 53, 17 and 39% C was decomposed from cowdung, rice straw and dhaincha, respectively. Decomposition was also found to be related to its nutrient status (C:N:S ratio). Considerable S mineralization was recorded in dhaincha and cowdung amended soil and attained the maximum at 15th day (13.8 μg g -1S) and 30 (9.8 μg g -1 S), respectively. During rice straw decomposition S was immobilized markedly. Application of plant and animal residues caused significant increase in P mineralization. Net mineralization was 95.5, 87.6 and 50.8 μg P g -1 soil in dhaincha, cowdung and rice straw amended soil, respectively.
PDF Abstract XML References Citation
How to cite this article
M.A. H. Chowdhury, R. Begum, M. R. Kabir and H. M. Zakir, 2002. Plant and Animal Residue Decomposition and Transformation of S and P in Soil. Pakistan Journal of Biological Sciences, 5: 736-739.
DOI: 10.3923/pjbs.2002.736.739
URL: https://scialert.net/abstract/?doi=pjbs.2002.736.739
DOI: 10.3923/pjbs.2002.736.739
URL: https://scialert.net/abstract/?doi=pjbs.2002.736.739
INTRODUCTION
Organic matter helps to build stable soil aggregates and improves soil structure, tilth, aeration and moisture holding capacity. Plant and animal residues are the major source of organic inputs to soil. This plant residue input has a primary role in maintaining soil organic matter content, microbial biomass activity and the size of the soil nutrient pool (Sanchez et al., 1989). Plant residues incorporated into the soil are decomposed by various soil organisms and become a part of the soil. Soil microorganisms grow rapidly during the decomposition of plant/animal residues (Brookes et al., 1990) and consequently some of the available soil nutrients, particularly N and S may be immobilized. However, the interaction between microbial growth, the immobilization and availability of S and P following the soil incorporation of plant/animal residues remains unknown. Decomposition rate of different plant and animal materials show different patterns. Decomposition of rice straw and dhaincha helped to increase the available P in soil (Mirsa and Das, 2000). However, the objectives of the present study were to determine the decomposition pattern of plant and animal residues and S and P transformation in soil.
MATERIALS AND METHODS
The experiment was carried out at the Department of Agricultural Chemistry, Bangladesh Agricultural University, Mymensingh during June to November 2001.
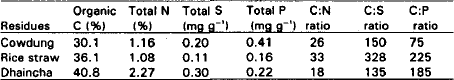
Plant and animal residues: Plant and animal residues viz., rice straw, dhaincha and cowdung were collected from Central Agricultural Farm, Agronomy Field and Dairy Farm of Bangladesh Agricultural University, Mymensingh, respectively. Some properties of the collected residues are presented in Table 1.
| Table 1: | Some selected characteristics of cowdung, rice straw and dhaincha used for amendment. |
 | |
Soil: The soil used in the experiment was S deficient silty loam soil with 0.61% organic C, CaCl2 extractable SO4-S of 6.0 μg g–1 soil and pH 6.17, collected from a selected area of Horticulture Farm, Bangladesh Agricultural University, Mymensingh. Undecomposed plant materials were removed by hand and the soil was sieved (<2mm). Samples were conditioned aerobically at room temperature and at 40% water holding capacity (WHC) for 7 days. This allowed the soil microbial population to stabilize, minimizing the effects of soil handling and preparation (Chowdhury et al., 2000). Immediately after conditioning, the soil was used for amending plant and animal residues.
Incubation experiment: Plant and animal residues were added to the soil at the rate of 1 g per 50 g (oven dry basis) soil and placed in a 100 mL glass jar. Each treatment had three replications. For each treatment, a nutrient solution was prepared by dissolving appropriate amounts of NH4NO3 and KH2PO4 in distilled water. 50 g soil (oven dry basis) sample was weighed in a 100 mL glass jar and amended with an aliquot of the nutrient solution (2 mL) having 250 μg N, 200μg P and 250 μg K g–1 soil. Two milliliters of nutrient solution was also added to the control soil to maintain N, P, K and moisture contents equivalent to those of amended soils.
Following amendment, glass jars were placed in 1L glass bottles, sealed and incubated at room temperature for 30 days. To trap CO2 evolved by soil microorganisms during incubation, 20 mL of 1M NaOH solution was placed inside each jar along with 10 mL distilled water at the bottom of the bottle to maintain internal humidity of the incubation environment. Available S and P were determined after 5, 10, 15 and 30 days of incubation.
Microbial activity: Microbial respiration were monitored from soil samples after 3, 6, 10, 15, 20 and 30 days of incubation. At each sampling, NaOH was renewed. Total CO2 was then titrated with standard HCl between pH 8.3 to 3.7 using pH meter (WTW pH 522). Microbial activity was expressed as μg CO2-C evolved g–1 soil day–1.
Soil chemical analyses: Available sulphur in soil was determined from CaCl2 (0.15%) extracts by colorimetric BaCl2 precipitation method as outlined by Page et al. (1982). Available soil phosphorus was extracted with 0.5 M sodium bicarbonate solution (Olsen et al., 1954) and develop blue colour by SnCl2 reduction and measured the colour colorimetrically at 660 nm (Black, 1965).
Statistical analysis: Collected data was statistically analyzed by the computer using statistical package programme MSTAT-C developed by Russel (1986). A one way ANOVA was made by F test. The pair comparisons were performed by least significant difference (LSD) test at 5% level of probability (Gomez and Gomez, 1984).
RESULTS AND DISCUSSION
The decomposition of plant and animal residues in the amended soils: Decomposition rates varied significantly among the treatments throughout the incubation period. Decomposition rate was measured as CO2-C (μg g–1 soil) evolution from soil. CO2 release varied making a peak at 3rd day for dhaincha (1143 μg CO2-C g–1 soil day–1) and cowdung (1044 μg CO2-C g–1 soil day–1) treatment while for rice straw it was (356 μg CO2-C g–1 soil day–1) at 30th day (Fig. 1).
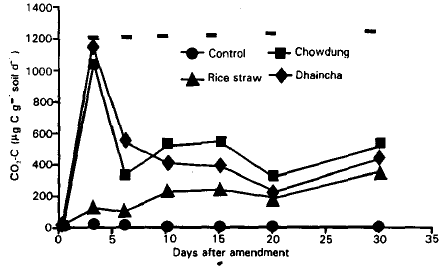 | |
| Fig. 1: | Decomposition of cowdung, rice straw and dhaincha as CO2 evolution (μg CO2-C g–1 soil day–1) in amended soil. Bars indicate LSD<0.05. |
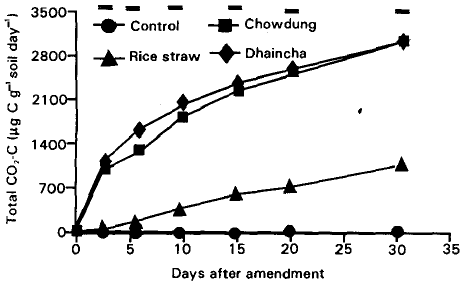 | |
| Fig. 2: | Decomposition of cowdung, rice straw and dhaincha as total CO2 evolution (μg CO2-C g–1 soil day–1) in amended soil. Bars indicate LSD<0.05. |
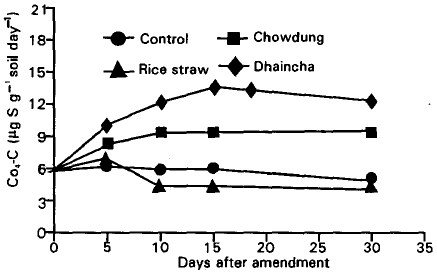 | |
| Fig. 3: | Sulphur transformation (μg S g–1 soil) in cowdung, rice straw and dhaincha amended soils during 30 days incubation. Bars indicate LSD<0.05. |
Up to 6th day, dhaincha amended soil showed the highest decomposition rate and rice straw amended soil showed the lowest rate with cowdung being intermediate. But from day 10 to the end of the incubation period, cowdung amended soil showed the maximum decomposition with dhaincha being intermediate and rice straw behaved similarly as early incubation days.
Decomposition of organic substrate is dependent on the nutrient status of the substrate itself. C:N ratio in the plant and animal residues is a determinant factor of organic matter decomposition (Waksman, 1961).
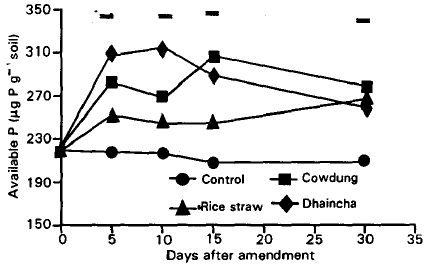 | |
| Fig. 4: | Phosphorus transformation (μg P g–1 soil) in cowdung, rice straw and dhaincha amended soil during 30 days incubation. Bars indicate LSD<0.05. |
In this experiment, plant and animal residues having different nutrient status were amended to the soil (Table 1). Dhaincha showed the highest decomposition rate (with C:N ratio 18) while rice straw (C:N ratio 33) decomposed very slowly all through the incubation period. These results confirmed the findings of Avnimelech (1986) who reported that a fast decomposition leads to a high rate of nutrient release only when the organic substrate is rich in nutrient (low C:N ratio). It was also observed that from the materials having high C:N and C:P ratios, nutrients will first be immobilized by growing microbial biomass and then be released from the manure to the soil. Melillo et al. (1982) also reported that decomposition rates were found to be related with the initial C:N ratio of the substrate and lower the nitrogen content or a wide C:N ratio of substrate, the slower the rate of decomposition.
After completion of incubation period, total 63, 3199, 1224 and 3159 μg C g–1 soil was respired for control, cowdung, rice straw and dhaincha amended soils, respectively. Initially 6019, 7212 and 8150 μg C g–1 soil was incorporated to the cowdung, rice straw and dhaincha amended soil, respectively. It was found that 53, 17 and 39% C was decomposed finally within cowdung, rice straw and dhaincha amended soil, respectively (Fig. 2). Both the plant and animal residues showed an exponential response in CO2- C production while unamended soil showed linear and lag phase. Wu et al. (1993) have shown that during early phase (0-30 days of incubation) of plant residue decomposition, priming effects are small, making it possible to use increase in CO2-C evolution in amended soils as an indication of the mineralization of the substrate during the early phase of decomposition.
Dhaincha and cowdung after amendments, decomposed rapidly than rice straw (Fig. 2). Decomposition was approximately 2 and 3 times double for dhaincha and cowdung, respectively as compared to rice straw in respect of added C to the soil as plant and animal residue. The rate and extent of mineralization of cowdung and dhaincha is comparable to that found using rape amendments (Wu et al., 1993) while rice straw with that of barley straw (Wu et al., 1993; Davenport et al., 1988) or cellulose amendments (Saggar et al., 1981).
The release and microbial immobilization of plant and animal residue-S: Total CaCl2-extractable S in the soil significantly varied during decomposition period following amendment (Fig. 3). Sulphur mineralization was observed for the amended plant and animal residues in soil. Immobilization took place just after 5 days in case of rice straw amended soil. On the other hand dhaincha and cowdung amended soil showed a considerable amount of S mineralization and attained the maximum at day 15 (13.8 μg S) and 30 (9.8 μg S), respectively.
A large amount of S was released from dhaincha (13.8 μg S) at day 15 and afterwards it decreased slightly. The amount of decreases was similar to that in the unamended soil and was due to the immobilization of native soil organic-S. These results suggest that the inorganic-S released from dhaincha during the first 15 days was not transformed further throughout the incubation and was available for plant uptake.
Residue-S release was comparatively slower but showed a similar pattern of S mineralization in cowdung amended soil but significantly higher than rice straw amended soil. The effect of straw incorporation on soil inorganic-S was different from those of dhaincha and cowdung. During the first 5 days following straw amendment, inorganic-S increased significantly. This increase was much smaller than those in the dhaincha and cowdung amended soil during the same period. In the rice straw amended soil inorganic-S dramatically decreased between 5 and 10 days from 7.2 to 4.6 μg S g–1 soil (Fig. 3).
The amount of inorganic-S in the rice straw amended soil was less than that in the unamended soil by day 10. Possibly, the S immobilization by the microbial biomass in the straw-amended soil during 5-10 days was from native S and included both soil inorganic-S and that converted from rice straw. Thus, in the straw amended soil, the immobilized soil available-S, together with all the inorganic-S released from the straw during 30 days incubation, was converted and transformed into soil organic fractions and as such would be no longer available for plant uptake.
Experimental results revealed that residues-S from dhaincha (easily-decomposable), cowdung (moderately-decomposable) and rice straw (relatively-slowly decomposable) is rapidly released following incorporation. This presumably because of the major S containing components of plant residues such as proteins and amino acids are very labile to soil microbial biomass and are decomposed prior to other more resistant fractions such as cellulose (Stevenson, 1986). Fitzgerald and Andrew (1984) have shown that methionine-S is rapidly converted to SO4-S and soil organic-S in 2 days following its addition to soil. Strickland et al. (1986) shown that most (70-100%) of the organic-S derived from forest litter can be mineralized or converted into other forms of organic-S after 7 days exposure in soil. Furthermore, results of the present study showed that once S was immobilized probably by the microbial biomass, S was directly transformed into the soil organic fraction. This suggests that the S immobilization by the microbial biomass and subsequent transformation into soil organic matter is not available for plant until it is remineralized.
Difference in organic S transformation due to incorporation of plant and animal residues in soil might be due to variation in decomposability and S content of the residues (Table 1). Barrow (1960) also reported that C:S ratio of organic materials was only a rough guide to their ability to release mineral S on incubation with soils. SO4-S usually accumulated in the soil when C:S ratio was below 200 and was usually immobilized when the ratio was above 400. Immobilization or mineralization could occur when the ratio ranges between 200 to 400.
The release and microbial immobilization of plant and animal residue-P: The amount of inorganic-P (Fig. 4) in the unamended soil slightly decreased during 30 days incubation (220 to 210 μg P g–1 soil) but amended soils showed significant increase in P content perhaps due to the mineralization of organic matter.
The results show that P mineralization pattern differed at each day of incubation. Highest net mineralization was 95.5 μg P g–1 soil in dhaincha amended soil at day 10 while that was 87.6 and 50.8 μg P g–1 soil at day 15 and 30 in cowdung and rice straw amended soil, respectively (Fig. 4). Both mineralization and immobilization of P were intermittently observed in plant and animal residues amended soil. At each day of incubation, availability of P differed significantly among the treatments. C:P ratio of the residues might result in different transformation pattern.
Microbial immobilization of P is a serious limiting factor in the availability and supply of this nutrient in soil particularly where the soil is low in P (Sauchelli, 1965). The soils used in this experiment were initially rich in P for both amended and unamended soils. After adding the residues, immobilization was recorded during microbial transformation of organic matter. Net immobilization was significantly lower in rice straw (having a wide C:P and C:N ratios) than dhaincha and cowdung (having low C:P and C:N ratio) amended soil.
A competition among microbial population in soil with low P content and low P availability has also been reported by Sauchelli (1965). Microbial immobilization of available P during organic matter decomposition may be beneficial since it may lead to a decrease in the fixation of inorganic P. Initial low release of inorganic P during decomposition of organic matter in present experiment provides a continuous supply of P with a minimal exposure to the different fixation mechanisms. The presence of organic matter in the soil effectively decreases the P fixation by the soil through acidifying and chelation mechanisms (Avnimelech, 1986).
The result showed that dhaincha decomposed more rapidly in soil than cowdung and rice straw, giving a pattern where dhaincha showed the highest decomposition rate and rice straw showed the lowest rate, while that of cowdung was intermediate. Decomposition rates were also found to be depended on C:N and C:S ratios of substrate. Application of plant and animal residues caused P and S mineralization in soil. It is important to note that, if a soil deficient in S is incorporated with plant and/or animal residues having wide C:S ratio, it should be followed by chemical fertilizer application. Otherwise the plants that will be grown, will suffer serious S deficiency.
REFERENCES
- Chowdhury, H., K.M. Kouno and T. Ando, 2000. Critical sulphur concentration and sulphur requirement of microbial biomass in a glucose and cellulose-amemded regosol. Biol. Fert. Soils, 32: 310-317.
CrossRef - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Melillo, J.M., J.D. Aber and J.F. Muratore, 1982. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology, 63: 621-626.
CrossRefDirect Link - Wu, J., A.G. O'Donnell and J.K. Syers, 1993. Microbial growth and sulphur immobilization following the incorporation of plant residues into soil. Soil Biol. Biochem., 25: 1567-1573.
CrossRefDirect Link