
S. A. Farooq
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box-36, PC 123, Sultanate of Oman
T. T. Farooq
Department of Biology, College of Science, Sultan Qaboos University, P.O. Box-36, PC 123, Sultanate of Oman
T. V. Rao
Tissue Culture Laboratory, Department of Botany, Osmania University, Hyderabad 500007
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 1 | Page No.: 43-46







ABSTRACT
Large-scale micropropagation has been successfully achieved in Annona squamosa L. from nodal explants. Several media with different hormones and culture conditions were tried in standardization of in vitro propagation of this plant. Effect of various surface sterilants in eliminating the contamination from the explants collected from the field has been worked out. Activated charcoal has a profound effect in preventing the polyphenol leaching in cultures. BAP, Kinetin, IAA, IBA and NAA were used either alone or in combination to induce shoot buds. Explants produced shoot buds in 7-14 days. Effect of hormones on rhizogenesis was studied. IBA was found to be more effective in root induction. True to type plantlets were obtained from the explants from elite trees. Present protocol is rapid, reproducible and less cumbersome over the conventional methods.
PDF Abstract XML References Citation
How to cite this article
S. A. Farooq, T. T. Farooq and T. V. Rao, 2002. Micropropagation of Annona squamosa L. Using Nodal Explants. Pakistan Journal of Biological Sciences, 5: 43-46.
DOI: 10.3923/pjbs.2002.43.46
URL: https://scialert.net/abstract/?doi=pjbs.2002.43.46
DOI: 10.3923/pjbs.2002.43.46
URL: https://scialert.net/abstract/?doi=pjbs.2002.43.46
INTRODUCTION
Advances in plant biotechnology research in the last decade has opened new vistas in the propagation of plants with improved resistance to diseases, pests, herbicides and stress etc. Micropropagation of fruit trees is an invaluable aid in the production of elite, disease free, breeding true-to-type in large numbers in short duration.
Annona squamosa L. (Custard apple) is a favourite table fruit of common man in Indian subcontinent. It belongs to family Annonaceae. It is cultivated throughout the tropical and subtropical regions of the world for its delicious and nutritive fruits. The pulp surrounding the seeds of ripe fruit is very delicious and nutritionally valuable with over 20% sugars. The fruit is usually eaten fresh, besides making an ingredient of refreshing drinks and also used to flavour the icecreams and puddings. A highly purgative tea is made from the roots and crushed leaves. Seeds have insecticidal properties and have oil that can be used in paint industry, while the seed cake is used as a manure.
The clonal propagation in Annona species is carried out by grafting and budding which are time consuming, while seedling rootstocks are highly variable resulting in the decreased productivity (George and Nissen, 1987). The seed propagation results immense variability affecting yield, size and quality of fruits. Due to the lack of germplasm of improved varieties its cultivation is limited. Micropropagation has been used in other species of Annona viz. Cherimoya A. cheimola (Tizzari et al., 1990; Rasai et al., 1995) and A. muricata (Lemos and Blake, 1996), but the reports on Annona squamosa are scanty. Present study deals with the establishment of standard protocols for the micropropagation of the large-scale rapid multiplication of improved genotypes, free of seed borne bacterial and viral diseases.
MATERIALS AND METHODS
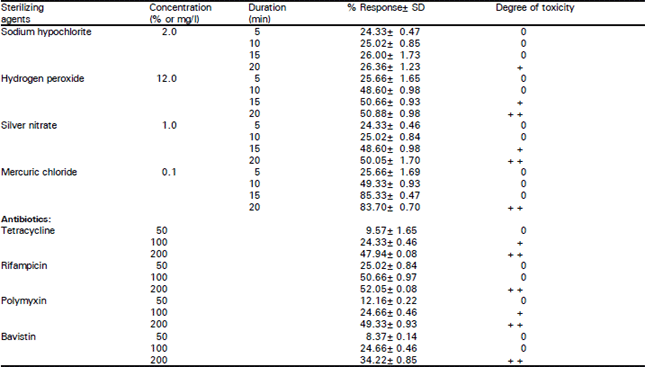
Plant material of Annona squamosa var. Balanagar was obtained from Fruit Research Station, Medak, A.P. in 1998. Nodal explants of 3 to 4 cm in length were collected from the upper, middle and lower regions of the newly sprouted branches of mature trees. Leaves were trimmed and explants were washed with enzyme based detergent for 20 min, followed by rinsing in chloroxylenol and washing several times with sterile distilled water containing 0.1% Tween. Damaged cut ends were removed, trimmed to a size of 1 to 1.5 cm with each explant having at least one node. To eliminate the contamination in the cultures, a comparative study was undertaken with various sterilants (Table 1) viz., Mercuric chloride, Sodium hypochlorite, Hydrogen peroxide, Silver nitrate and antibiotics such as Tetracycline, Rifampicin, Polymyxin B and Bavistin (systemic fungicide) and their efficiency was evaluated.
Selection of an appropriate culture medium and the use of correct growth regulators were critical for the optimum growth response of the explants. Three different types of media such as MS medium (Murashige and Skoog, 1962) WPM (Woody Plant Medium, Lloyd and McCrown, 1980) and SH medium (Schenk and Hilderbrandt, 1972) along with different growth regulators were used.
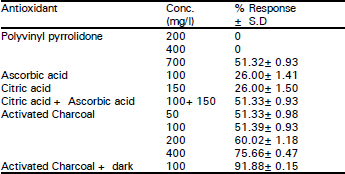
To prevent the leaching of polyphenols from the cut ends of the explants, the following procedures were employed and their effectiveness was studied. I) Pretreatment of explants before inoculation with antioxidant solution (Citric acid 150 mg/l + Ascorbic acid 100 mg/l). ii) Adding antioxidants such as Charcoal (0.1 to 0.5 %), Polyvinylpyrrolidone PVP (200 to 700 mg/l), Ascorbic acid (100 mg/l), Citric acid (150 mg/l) to the medium. iii) Incubation of explants in darkness prior to inoculation. All the aseptic manipulations were carried out in laminarflow. After inoculation, cultures were incubated in cool white fluorescent light (2000 lux) at photoperiods ranging from 12 to 18 hours at a temperature of 24 ± 2 oC. Experiments were conducted with different light intensity viz. 1000-4000 lux. The explants were oriented in horizontal, vertical and slant positions to find out optimum response. The response of the explants in the culture was estimated as the ratio of number of explants responding to the treatment to the total number of the explants inoculated, and is expressed as percentage.
Results and Discussion
The success of micropropagation of Annona squamosa through nodal culture involves a number of factors, which affect directly or indirectly the proper establishment of explants in the medium. These are systematically studied to standardize a suitable protocol for the large-scale in vitro clonal propagation.
Of all the treatments for controlling contamination, the most efficient response (85.33%) was obtained by immersion and continuous agitation of explants in 0.1% Mercuric chloride for 15 min. followed by rinsing twice with sterile distilled water. Longer treatments were found to cause irreversible damage to the explants, while treatment of less than 15 min. were relatively less efficient (49.33%) (Table 1).
| Table 1: | Response of explants to the sterilizing agents |
 | |
| 0 = No toxicity, + = less than 50%, ++ = more than 50% | |
| Table 2: | Effect of antioxidants in controlling the polyphenol leaching into the media |
 | |
Treatment of explants with 12% Hydrogen peroxide or 1% Silver nitrate over 15 minutes resulted in 50% surface sterilization but most of them turned black due to toxicity. Sodium hypochlorite was found to be least effective in controlling the contamination.
Different antibiotics and fungicides at varying concentrations were incorporated into the medium to eliminate the microbial contaminants from the explants (Table 1). Most of these produced undesirable effects on explants such as delay in bud induction and fewer buds per explant. Phillips et al. (1981) reported similar observations in Helianthus tuberosus. Of all the antibiotic treatments, Rifampicin (100 mg/l) was found to be very effective in controlling the bacterial contamination. Polymyxin B and Tetracycline (100 mg/l) were found to be ineffective and toxic to the explants. Bavistin (100 mg/l) was found to be least toxic and was effective in controlling only the fungal contamination. These results are in conformity with the reports of D’Silva and D’Souza (1993) in cashew nut. The use of antibiotics to control the growth of contaminants in the medium was earlier reported by Pollock et al. (1983) and fungicides by Shields et al. (1984).
The nodal explants of Annona squamosa release a number of polyphenol compounds into the medium, which hamper its establishment. To prevent the polyphenol leaching, various antioxidants were incorporated in the medium. The best response (91.88%) was obtained with activated Charcoal and dark incubation of the cultures (Table 2). Explants showed faster shoot bud induction along with the production of maximum number of buds. At lower concentrations of Charcoal lesser response was obtained. The advantage of Charcoal in the medium was earlier reported by Fridborg et al. (1978), Weatherhead et al. (1978), D’Silva and D’Souza (1993). Activated charcoal absorbs the polyphenolic compounds and the dark incubation minimizes the synthesis of polyphenols by explants. Addition of Citric acid and Ascorbic acid (100 mg/l) individually to the medium was not effective in preventing the leaching and resulted in the discoloration of the explants. Pre-treatment of explants with Citric acid and Ascorbic acid (150+100 mg/l) was partially effective with only 51.33% of explants responding. PVP at lower concentrations (100 to 400 mg/l) was not effective in controlling the leaching of polyphenols, whereas, higher concentrations (500 to 800 mg/l) delayed the shoot bud induction.
Slightly horizontal orientation of the explants in culture medium produced the best response. In the vertical orientation the response was relatively less. Explants placed in the inverted position resulted in delayed shoot bud induction and callus production was prominently observed at the cut ends. The present findings are not in agreement with the observations of Amin and Jaiswal (1987) in guava, where initial culturing of explants in the vertical plane and then shifting to horizontal position resulted in maximum shoot induction. In apple, inverted orientation of explants gave maximum shoot bud induction (Zimmerman and Ingrid, 1989).
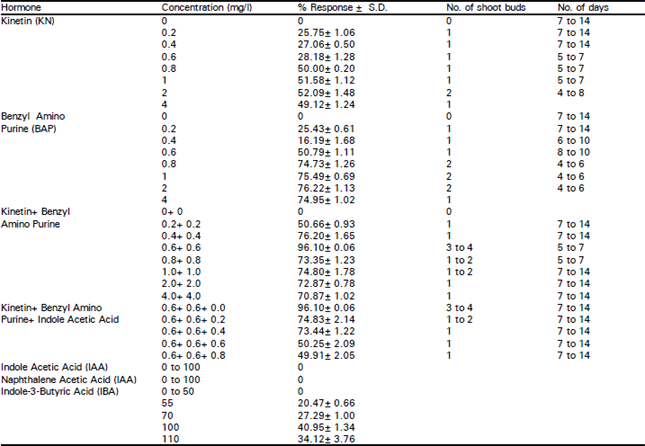
| Table 3: | Effect of hormones on the shoot bud induction |
 | |
| Table 4: | Evaluation of different media for the efficiency of shoot induction |
 | |
Light influences the micropropagation through photoperiod, intensity and spectral quality (Athanasios et al., 1987). The light intensity levels were studied at 1000 to 4000 lux. Of all the light treatments 2000 lux for 18 hr resulted in maximum shoot bud induction within a short time. While light below 2000 lux delayed it and resulted in yellowing of shoots. Light above 2000 lux resulted in rapid shoot bud initiation, but they failed to grow further.
The effect of three different media (MS, WPM and SH) on the shoot induction from nodal explants was studied (Table 4). These media differed mainly in their basal inorganic components such as nitrates and sulfates. The compared media were supplemented with the same amounts of sucrose, agar and phytohormones. MS medium was found to be superior since the number of days for shoot bud induction was lesser followed by WPM and SH (Rao and Farooq, 1994).
Carbohydrates act as a source of energy and as osmotic agents (Deburg, 1977). Sucrose is supplied as the main carbohydrate in the medium. Different concentrations of sucrose (0, 0.5, 1, 2, 3, 4 and 5%) were studied. Of which 2% sucrose was found to be optimum. Sucrose lower than 2% delayed shoot induction, whereas higher concentrations induced shoot buds, which failed to grow further. Similar findings were earlier reported in tobacco callus cultures by Brown et al. (1979).
Different phytohormones (BAP, Kinetin, NAA, IAA and IBA) at varying concentrations were used in the medium either singly or in combination with others and their effects on the shoot bud induction, growth of shoots and rhizogenesis was systematically evaluated (Table 3). Kinetin and BAP (0 to 4 mg/l) were used either singly or in combination with each other. The frequency of shoot induction with either Kinetin or BAP alone was very low and time for the initial leaf pulvinus abscission and shoot bud formation was 4 to 14 days. Usually one or occasionally two shoot buds formed in most of the nodal explants. Callusing was observed at higher concentrations of Kinetin and BAP (4 mg/l) along with a decrease in number of buds. When Cytokinin was eliminated there was no induction of buds in explants.
Double hormone treatments (Kinetin and BAP) in equimolar concentration had a profound influence on the initial leaf pulvinus abscission and the time taken for shoot bud induction. Similar observation was reported by Nair et al. (1984 b). The best response was observed with Kinetin 0.6 mg/l + BAP 0.6 mg/l, where as with Kinetin 0.8 mg/l + BAP 0.8 mg/l slight callusing was observed with decrease in number of buds. With BAP 1 mg/l + Kinetin l-4 mg/l, there was a gradual fall in number of induced buds per explant with increase in the callusing at cut ends. Polyphenol leaching from the explants was also prominent.
Triple hormone treatments included auxin (NAA) along with BAP and Kinetin in equimolar concentrations. With Kinetin 0.6 mg/l + BAP 0.6 mg/l + NAA 0.2 mg/l, shoot bud induction and the number of buds per explant was reduced. However, when NAA was doubled to 0.4 mg/l keeping the other two constant, there was an increase in callusing with decreased shoot bud formation. Similarly, when NAA was increased to 0.6 mg/l there was further increase in callus.
The nodal explants along with induced buds were transferred to media containing MS basal + Kinetin 0.6 mg/l + BAP 0.6 mg/l + 3% sucrose + 0.8% agar and cultured for a month. To induce elongation the induced shoot buds were separated and cultured in MS basal + 0.2 mg/l Kinetin + 3% Sucrose + 0.1% Charcoal. In 20 to 25 days, these isolated buds have grown to a size of 2-3 cm and continued to grow further with occasional multiple branches.
In vitro grown shoots were subcultured to increase the number of propagules in the following ways I) MS basal liquid medium containing 0.1% Charcoal for 15 days. ii) White (1963) solid medium containing 1 mg/l BAP+ 2% Sucrose and iii) in Whites liquid medium supplemented with 1 mg/l BAP + 2% Sucrose. After 45 days, the explants cultured in Whites solid medium, produced 5 to 6 shoots per explant, where as explants cultured in Whites liquid medium suffered necrosis and failed to produce buds.
In vitro rhizogenesis in Annona was found to be very difficult. After multiplication, the shoots (2 to 4 cm) were used for rhizogenesis. Since the shoots taken from the multiplication medium failed to respond to rooting treatment, they were first cultured in MS basal medium containing 2% Sucrose + 1 gm/l activated Charcoal for 15 days and then were used for rhizogenesis. Nair et al. (1984 a) reported rooting of atemoya in MS semi-solid medium. In present study, White’s media were used for root induction in both liquid and solid phases. Presence of 2% Sucrose and 0.1% activated Charcoal was found to be ideal for in vitro rooting.
IAA, IBA, NAA were used for inducing roots from the regenerated shoots at varying concentrations ranging from 0-100 mg/l. IAA and NAA were ineffective in inducing the roots and resulted in callus formation at the base of the shoot even at low concentrations, while, IBA was found to be effective in inducing roots. At lower concentrations of IBA (below 55 mg/l) swelling of the cut ends of the shoots was observed. In Annona cherimola, Encina et al. (1994) reported the use of high concentrations of IBA for rooting. In present study, in vitro grown shoots cultured in liquid White’s medium for 3 days in dark and 7 days in light followed by a transfer to White’s solid medium without hormones produced thin hairy roots. Best response was obtained when shoots were cultured in 100 mg/l IBA, but defoliation occurred. The rhizogenesis obtained in this study is an improved rooting response over the reports of Nair et al. (1984 a,b) in shoots derived from leaf culture and the endosperm culture (Nair et al., 1986). Roots were also induced with 110 mg/l IBA however, defoliation of shoots occurred within 2 days after root induction.
For acclimatization, rooted plantlets were first washed carefully with distilled water to remove agar sticking to the roots and the plantlets were carefully transferred to smaller plastic seed plugs containing a mixture of sterile vermiculate and sand (1:1) and covered to maintain abundant moisture around the plant and incubated at 24±2oC with 18 hours photoperiod. After one week, covers were removed and plants were exposed to external moisture levels gradually. Plants were transferred to bigger plastic pots containing the same mixture and placed in green house. Elongation was observed in transplanted shoots and new leaves emerged after 25 to 30 days. After two weeks, plants were transferred to earthen pots for further hardening.
REFERENCES
- Amin, M.N. and V.S. Jaiswal, 1987. Rapid clonal propagation of guava through in vitro shoot proliferation on nodal explants of mature trees. Plant Cell Tissue Organ Cult., 9: 235-243.
CrossRefDirect Link - Fridborg, G., M. Pedersen, L.E. Landstrom and T. Eriksson, 1978. The effect of activated charcoal on tissue culture; adsorption of metabolites inhibiting morphogenesis. Physiol. Plant., 43: 104-106.
CrossRefDirect Link - George, A.P. and R.J. Nissen, 1987. Propagation of Annona species: A review. Scient. Hortic., 33: 75-85.
CrossRefDirect Link - Lloyd, G. and B. McCown, 1980. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Proc. Int. Plant Propagator's Soc., 30: 421-426.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Schenk, R.U. and A.C. Hildebrandt, 1972. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot., 50: 199-204.
CrossRefDirect Link