
Toshiyuki Matsui
Department of Bioresource Production, Faculty of Agriculture, Kagawa University, Miki-Cho, Kagawa, 761-0795, Japan
Yusuke Kosugi
Department of Bioresource Production, Faculty of Agriculture, Kagawa University, Miki-Cho, Kagawa, 761-0795, Japan
Tomohiro Yanagi
Department of Bioresource Production, Faculty of Agriculture, Kagawa University, Miki-Cho, Kagawa, 761-0795, Japan
Haruo Suzuki
Department of Bioresource Production, Faculty of Agriculture, Kagawa University, Miki-Cho, Kagawa, 761-0795, Japan
Pankaj Kumar Bhowmik
Department of Bioresource Production, Faculty of Agriculture, Kagawa University, Miki-Cho, Kagawa, 761-0795, Japan
Sutevee Sukarakarn
Tropical Vegetable Research Center Kasetsart University, Bangkok 10900, Thailand
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 2 | Page No.: 208-211







ABSTRACT
The genetic relationship of 13 oriental melon (Cucumis melo L.) was determined using random amplified polymorphic DNA (RAPD) analysis with 12 dodecamer oligonucleotide primers. Representative cultivars could be divided into two major groups at 0.756 similarity value. One group included 7 makuwa melons namely, ‘ Nara-1-gou ’, ‘ Tiger-melon ’, ‘ Kinpyou ’, ‘ Kiku-melon ’, ‘ Nashi-makuwa ’, ‘ Shouwa-melon ’ and 4 conomon melons namely, ‘ Numame-shirouri ’, ‘ Katsura-ohoshirouri ’, ‘ Tokyo-ohoshirouri ’, ‘ Kuromonao-ohoshirouri ’. The other major group included 2 makuwa cultivars namely ‘ Ginsen ’ and ‘ Wasegin ’. The RAPD showed that conomon melon belonged to same species as makuwa melon. It seemed that, makuwa melon is an allogamous plant which hybridizes easily with table-melons and conomon melon and it might have hybridized with conomon melon in the distant past.
PDF Abstract XML References Citation
How to cite this article
Toshiyuki Matsui, Yusuke Kosugi, Tomohiro Yanagi, Haruo Suzuki, Pankaj Kumar Bhowmik and Sutevee Sukarakarn, 2002. Classification of Oriental Melon by RAPD Analysis. Pakistan Journal of Biological Sciences, 5: 208-211.
DOI: 10.3923/pjbs.2002.208.211
URL: https://scialert.net/abstract/?doi=pjbs.2002.208.211
DOI: 10.3923/pjbs.2002.208.211
URL: https://scialert.net/abstract/?doi=pjbs.2002.208.211
INTRODUCTION
Oriental melons consisting of Japanese sweet melon (makuwauri) and pickling melon (shirouri or conomon) were introduced from China but both have unknown origin (Shimizu, 1977). Native sweet melon was imported to Japan before the 8th century and it only developed during the 18th century. In order to produce good cultivars by selfing or crossing, Japanese sweet melons ‘ Ginsenmakuwa ’, ‘ Nashimakuwa ’ and ‘ Natsumeuri ’ were also introduced to Japan during the middle of the Meiji period. On the other hand, native pickling melon was imported to our country at the same period with the sweet melon and it was used as a raw material for pickles. In the meantime, oriental melons were classified into several types based on morphological, ecological and physiological differences by Katsumata and Yasui (1964 ), and Hosoki et al. (1990). Asahira and Ohoi (1976) also classified the oriental melons into 4 types based on isozyme banding patterns. However, their classifications were not necessarily clear to differentiate genetic relationship analyzed by conventional methods. In recent years, random amplified polymorphic DNA (RAPD) analysis, proposed by Welsh and McClelland (1990) and Williams et al. (1990), has been established to be a useful tool for genetic relationship and typing. Identifying olive varieties ( Sanz-Coretes et al., 2001), common olive varieties (Wiesman et al., 1998), wild radishes (Yamagishi et al., 1998), apricot varieties (Takeda et al., 1998), sweet potato (Thompson et al., 1997) and cranberry (Novy et al., 1996) have been carried out using decamer nucleotide primers to generate RAPD polymorphism. In this paper, we report the use of RAPD markers to distinguish among 13 oriental melon cultivars and we discuss relationships among cultivars of this species.
MATERIALS AND METHODS
Plant materials: Thirteen Japanese cultivars belonging to 9 oriental sweet melon (makuwa) and 4 oriental pickling melon (conomon) were studied (Table 1). Oriental melons were grown in green houses at the Department of Bioresource Production, Kagawa University, Japan in May to June 1998. Young leaves ( ca. 5 g) were collected in June and stored at -85°C for extraction of DNA.
DNA extraction: Total DNA was extracted from leaf tissue following the CTAB method (Murray and Thompson, 1987) with minor modification. Frozen leaves (1 g) were pulverized in 50 ml of liquid nitrogen and ground with 2 ml of 1X CTAB buffer ( 2% CTAB, 100 mM Tris-HCl pH 8, 1.4 M NaCl, 20 mM EDTA, 1% PVP) in a cold mortar and pestle. After 10 min samples were incubated at 60°C for 60 min, mixed with 2 ml of chloroform-isoamyl alcohol (24:1) followed by incubation at room temperature for 30 min and centrifuged at 2,800 rpm for 15 min. The aqueous phase was added to a new tube and the same procedure was repeated under the same conditions until centrifugation. The combined aqueous phase was added to 1/10 volume of 10% CTAB and mixed with an equal volume of precipitation buffer (1% CTAB, 5 mM Tris-HCl pH 8, 10 mM EDTA). After centrifugation of the sample, the precipitate was dissolved with 2 ml of NaCl-TE buffer (1 M NaCl, 10 mM Tris-HCl pH 8, 1mM EDTA) and added to 2 ml of isopropanol. The nucleic acid precipitation was washed with 2 ml of 70% ethanol, dried under vacuum for 5 min and completely dissolved with 500 μl sterilized water. A 1/10 volume of RNase solution (100 μg /ml) was added to DNA solution, and followed by reaction for 30 min at 50°C. The DNA length and concentration were determined in comparison with standard λ-DNA by electrophoresis with 1.2% agarose gel in TAE buffer, and by spectrophotometry, respectively. The DNA solution was stored at -30°C until needed for use.
Oligonucleotide primers: RAPD primers were obtained from Wako DNA Oligomer (12 mer) Set C (Tokyo, Japan). The sequence of 12 primers are listed in Table 2.
PCR: Amplification reactions were carried out using 25 μl reaction mixtures containing 10 mM Tris-HCl pH 8.3, 50 mM KCl, 1.5 mM MgCl2, 0.01% gelatin, 200 μM each of dNTPs, 200 nM of each primer (400 nM in cases of single primer PCR), 0.1 U of Ampli Taq DNApolymerase (Perkin Elmer Cetus, New Jersey) and 10ng of genomic DNA. Each reaction mixture was overlayed with 50 μl of liquid wax ( Funacoshi, Tokyo) to prevent evaporation. DNA amplification reactions were performed in a thermal cycler (PC-300, Astec, Tokyo) programmed for 1 cycle of 5 min at 94 °C followed by 40 cycles of 30 sec at 94 °C, 15 sec at 42 °C, 1 min at 72 °C, for denaturing, annealing and DNA extension, respectively. The last cycle was followed by incubation at 72 °C for 5 min. Amplification products were analyzed by gel electrophoresis in 1.5% agarose (Wako Co., Tokyo) in TAE buffer.
| Table 1: | Characters of makuwa and conomon groups used in this experiment |
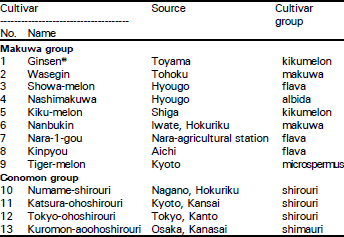 | |
| Katsumata and Yasui (1964) * Takii seed Co. | |
| Table 2: | List of primers which produced amplified DNA fragments |
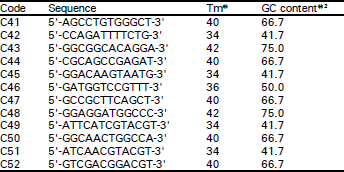 | |
| * The denaturation temperature was calculated using the following equation: Tm (°C) = 4x (G+C) + 2x (A+T) *2 GC content was calculated by number of GC/total basic number | |
After electrophoresis, the gel was stained with ethidium bromide and visualized by illumination with UV light. Molecular sizes of amplification products were estimated using a λ-DNA digested by Hind III/EcoR I and φ_X174 digested by Hae III . All reactions were conducted three times, using different samples, which were collected at three different dates. Only reproducible bands were used for further analysis.
Genetic analysis: Data generated from RAPD analyses were analyzed using the Nei similarity index ( Nei and Li, 1979) which excludes common negative data on the basis of the equation, Similarity= 2Nab/(Na+Nb).
Where Nab= number of scored amplification fragments with the same molecular weight shared between genotype a and b; Na= number of scored amplification fragments in genotype a, and Nb= number of scored amplification fragments in genotype b. A dendrogram was constructed on the basis of the similarity matrix data by unweighted pair group method with arithmetic average (UPGMA) cluster analysis using the software MEGA program (Pensylvania State University, PA).
RESULTS
Screening of the primers: When the 12 primers were used by combining one primer and/or two primers in a single PCR, 78 different combinations could be made as PCR primer (Klein-Lankhorst et al., 1991). Of the 78 different combinations, the primers which produced the characteristic patterns of bands are shown in Table 3. On the other hand, the non-utilized same band was shown at 16%. No bands were detected using the following primers (20%): the single primer was C41, C42, C45, C46, C49 and C51 whereas the two primers were C4142 (combination of C41 and C42), C4145, C4245, C4246, C4251, C4546, C4551, C4651 and C4951.
In order to prevent misextending incorrect nucleotides and incorrect annealed primer, annealing temperature was designed to raise as high as possible within the range of melting temperature ( Tm ) (Fujinaga, 1990) and was selected at 42 °C. GC content is an important index for the selection of primers because the GC content of primer is associated with the Tm and related to the reproducibility of data (Fritsch et al., 1993). The primer C48 which had high (75%) GC content showed a large number of amplified fragments and higher fragment intensity than the primers with lower GC content. The higher GC content produced higher frequencies of RAPDs because of increased total amplified fragments (Takeda et al., 1998). Although the primer C41 and C50 showed higher GC content than the other primers, no polymorphic fragment was detected. In this case, the primer C41 and C50 seemed to produce a stem loop and hairpin loop structures, respectively (Shiba, 1994).
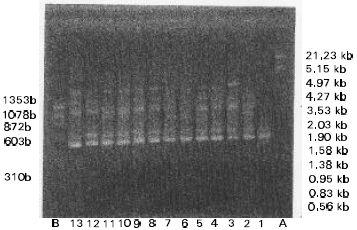 | |
| Fig. 1: | PARD pattern obtained from makuwa and conomon group with the primer C4552. Symbols 1 to 13 are shown in Table 1. A; φ x 174 (Hae III digested), B; λ DNA (Hind III/Ecor I digested). |
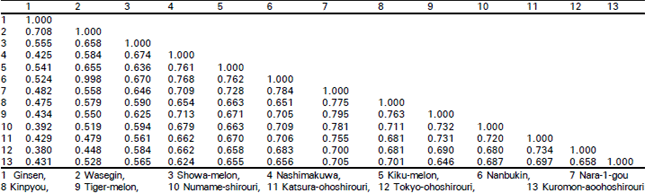
Similarity and dendrogram of makuwa and conomon melons: Fig. 1 shows the RAPD pattern obtained from makuwa and conomon melons with the primer C4552. The amplification pattern of presence/ absence was constructed, followed by DNA typing (pattern). Several unique markers were detected (Table 3). C4351-0.8 ( Kb ), C4351-0.28 and C4950-0.6 were specific to ‘Ginsen’, C4243-1.4, C4449-1.9, C4652-3.8, C4850-0.45 and C52-3.8 were specific to ‘Shouwa-melon’ whereas C4452-0.8 was absent in ‘Ginsen’ and C4547-2.0 was absent in ‘Wasegin’. On the basis of the DNA typing by RAPD markers, a similarity matrix was calculated using the Nei similarity index. Table 4 shows the similarity values of makuwa melon. The similarity values ranged from 0.380 for ‘Tokyo-ohoshirouri’(12) and ‘Ginsen’(1) to 0.784 for ‘Nara-1-gou’(7) and ‘Nanbukin’(6). The higher the value, the closer the relation between the two cultivars.
The dendrogram resulting from the UPGMA cluster analysis is shown in Fig. 2. Results indicate that the makuwa melon could be divided into two major groups at 0.756 similarity value. The first group could be subdivided into two subgroups. One subgroup included 6 cultivars namely, ‘ Nara-1-gou ’(7), ‘Tiger-melon’(9), ‘ Kinpyou ’( 8), ‘ Numame-shirouri ’(10), ‘Katsura-ohoshirouri’(11) ‘Tokyo-ohoshirouri ’(12). The second subgroup included 5 cultivars namely, ‘Kiku-melon’(5),
| Table 3: | Specific distinguishable bands of individual makuwa and conomon group |
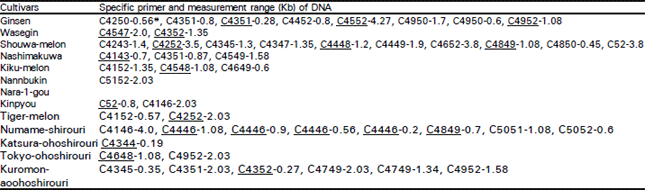 | |
| * Combination of C42 and C50 and nucleotide separated on agarose gel of 0.56 Kb. Under line shows only one absent band. The single primer C41, C42, C45, C46, C49 and C51 did not produce any useful band. | |
| Table 4: | Similarity matrix of makuwa and conomon groups, generated by using Nei’s estimate of similarity |
 | |
 | |
| Fig. 2: | Dendrogram of 13 makuwa and conomon groups by UPGMA cluster analysis |
‘Nashimakuwa’(4), ‘Nanbukin’(6), ‘ Kuromonao-ohoshirouri’(13) and ‘Shouwa-melon ’(3). The other major group included yuki-melon group of ‘ Ginsen ’(1), and ginmakuwa group of ‘ Wasegin ’(2).
DISCUSSION
Of the 12 single primers used in experiment, 7 could not be used as RAPD primers due to non-detection of bands. The combined primers method yielded more bands as compared with single primers methods.
When the results of the RAPD analysis were applied to morphological characters, it was found that one group included makuwa group and conomon group. The other major group included makuwa and kikumelon group. In this group, ‘ Nara-1-gou’ , ‘Kinpyomakuwa’ (flava group) and ‘Tiger-melon’ were subdivided into one section, while the other melons were subdivided into another section. Although ‘ Tiger-melon’ was not clear with crossing parents, it was supposed to be a crossing between microspermus group and makuwa group on the basis of morphological characters, sweet taste and aroma. ‘Katsura-ohoshirouri’ and ‘Tokyo-ohoshirouri’ belonged to conomon group and large fruits. Hosoki et al. (1990) divided into two groups based on the morphological, ecological and physiological characters and conomon group belonged to one major group. In our classification, conomon group did not belong to one group and located into kikumelon and flava group. ‘Numame-shirouri’ shown to be closely related to ‘Kinpyomakuwa’ due to its middle fruit. ‘Kikumelon’ (kikumelon group) and ‘Nambukin’ (makuwa group) showed a very close relationship with ‘Nashimakuwa’ (albida group). ‘ Nambukin ’ domesticated from ‘ Kinmakuwa’ (albida group) has a golden or yellowish peel and less than 10 lengthwise furrows like ‘Kikumelon’. ‘Kikumelon’ has a grayish white peel, and white and sweet flesh like ‘Nashimakuwa’. Although three groups have different morphological characters, their fruits have something in common with small and globular shape.
‘Kuromonao-ohoshirouri’ belonged to conomon group but had a dark greenish peel with light greenish striped pattern. Shouwa-melon’ (flava group) was supposed to be a crossing species between flava group and other melon, and showed a distant relationship to ‘ Nara-1-gou’ and ‘ Kinpyoumakuwa’ in spite of being the same group. ‘Ginsen’ was a crossing species between kikumelon group and flava group, and it was a small and an improved breed. ‘ Ginsen’ showed a very close relationship to ‘ Wasegin’.
ACKNOWLEDGMENT
The authors wish to thank Miss. K. Shimada of Katokichi Co. Ltd. for technical assistance.
REFERENCES
- Murray, M.G. and W.F. Thompson, 1980. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res., 8: 4321-4326.
CrossRefPubMedDirect Link - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Welsh, J. and M. McClelland, 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res., 18: 7213-7218.
CrossRefPubMedDirect Link - Wiesman, Z., N. Avidan, S. Lavee and B. Quebedeaux, 1998. Molecular characterization of common olive varities in Israel and the West bank using randomly amplied polymorphic DNA (RAPD) markers. J. Am. Soc. Hort. Sci., ` 123: 837-841.
Direct Link