
Ali H. Sayyed
Department of Biological Sciences, Imperial College of Science, Technology and Medicine, Silwood Park, Ascot, Berkshire, SL57PY, UK
Denis J. Wright
Department of Biological Sciences, Imperial College of Science, Technology and Medicine, Silwood Park, Ascot, Berkshire, SL57PY, UK
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 12 | Page No.: 1330-1344







ABSTRACT
The bacterium Bacillus thuringiensis is the main source of insecticidal proteins in insect resistant plants. However, biochemical and genetic studies have shown that insect resistance to B. thuringiensis (Bt) toxins can occur and with the advent of Bt transgenic crops this is a major concern. Several insect species have shown resistance to these toxins in the laboratory but the diamondback moth, Plutella xylostella is the only species which has evolved resistance under field conditions to date. Many studies have been done to elucidate the mode of action of the toxins and the mechanisms and genetics of resistance. In this article Bt toxins, their mode of action, mechanisms and genetics of resistance and management strategies for delaying resistance are reviewed. The emphasis is placed on examining the presently recommended high dose/refuge strategy.
PDF Abstract XML References Citation
How to cite this article
Ali H. Sayyed and Denis J. Wright, 2002. Genetic Diversity of Bt Resistance: Implications for Resistance Management. Pakistan Journal of Biological Sciences, 5: 1330-1344.
DOI: 10.3923/pjbs.2002.1330.1344
URL: https://scialert.net/abstract/?doi=pjbs.2002.1330.1344
DOI: 10.3923/pjbs.2002.1330.1344
URL: https://scialert.net/abstract/?doi=pjbs.2002.1330.1344
INTRODUCTION
Since the widespread introduction of synthetic insecticides in the late 1940s, a steady stream of new compounds has been developed. Due to their efficacy and specificity these insecticides are much more effective than the control methods that preceded them. However, due to the specific mode of action and ease of metabolism of many of these pesticides, pest populations can negate them by utilizing alleles which provide resistance and pesticide resistance has doggedly followed the introduction of new pesticides (Roush and Tabashnik, 1990).
Since the first commercial Bacillus thuringiensis (Bt) formulations appeared in the late 1930’s, they have been used widely as niche products for pest control. It had been presumed that resistance to Bt toxins was unlikely because of its unique mode of action (Bauer, 1995). However, this was subsequently shown not to be the case and the Indian mealmoth, Plodia interpunctella Hüb. (Lepidoptera:Pyralidae) was the first insect reported to have developed resistance to Bt toxins, albeit under laboratory conditions. While various other insect species have been subsequently shown to develop resistance under laboratory conditions (Table 1), the diamondback moth, Plutella xylostella L. (Lepidoptera:Plutellidae) remains the only insect species which has developed resistance in the field. The development of resistance to Bt toxins is seriously threatening their life expectancy as pest control agents, particularly with the introduction of commercially grown Transgenic crops expressing insecticidal Bt proteins which increase the risk of resistance by providing a constant selection pressure (Wright et al., 1997; Tabashnik et al., 1998). As many major pests have already developed resistance to chemicals, the fact that Bt makes up to 98% of all biopesticides and that the demand for Bt crops is increasing makes the impact of resistance potentially dramatic.
100 years of Bacillus thuringiensis discovery: Bacillus thuringiensis represents a major group of microbes used for insect biocontrol (Macintosh et al., 1991). It is a gram-positive soil bacterium distinguished from other bacilli by its production of parasporal crystal proteins (Yamamoto and Powell, 1993). Bacillus thuringiensis is quite closely related to B. cereus (Carlson et al., 1994) and classical biochemical and morphological methods of classifying bacteria have consistently failed in distinguish B. thuringiensis from B. cereus. Chromosomal DNA hybridization, phospholipids and fatty acid analysis and genomic restriction digest analysis support a single-species hypothesis (Schnepf et al., 1998). Cry toxin synthesis is also transmissible from B. thuringiensis to B. cereus via conjugation (Gonzalez et al., 1982).
Bacillus thuringiensis was first discovered as a potential microbial agent for insect control in 1901 when the Japanese bacteriologist Ishiwata (1901) isolated this bacterium from diseased larvae of Silkworm, Bombyx mori L. (Lepidoptera:Bombycidae) and named it “Sottokin” mean “sudden death Bacillus”. A decade later, Ernst Berliner isolated a similar organism from a diseased granary population of the flour moth Anagasta kuehniella Zeller (Lepidoptera: Pyralidae) from Thuringia, Germany (Berliner, 1911, 1915). Aoki and Chigasaki (1916) reported that its activity was due to a toxin present in sporulated cultures and that the toxin was not an exotoxin. In 1927, Matte re-isolated the organism from the same host, as did Berliner (Heimpel and Angus, 1960) and both Berliner and Matte observed, in addition to the spore, a second body which they called a “Restkorper” in the developing sporangia (Beegle and Yamamoto, 1992).
The first attempts to use Bt as a biopesticide using B. thuringiensis var. kurstaki (Btk), were carried out in the late 1920s and early 1930s against the European corn borer, Ostrinia nubilalis Hüb. (Lepidoptera: Pyralidae) in South East Europe (Van Frankenhuyzen, 1993). The first commercial product, Sporein®, became available in 1938 in France, primarily for control of P. interpunctella (Weiser, 1986). Work by Steinhaus (1951) restimulated interest in the commercial exploitation of Bt for control of lepidopteran pests of field crops and by 1957 the product Thuricide® (Btk) was available (Beegle and Yamamoto, 1992). The availability of commercial products initiated a period of “intermittent field testing” throughout the 1960s with inconsistent results (Mott et al., 1961). This resulted in two significant developments. First, the discovery of HD-1 a Btk isolate, which proved to be more potent than the isolates in existing commercial Bt products (Dulmage, 1970). The use of HD-1 population was also encouraged by its broad spectrum of activity against more than 100 lepidopteran species (Navon, 1993). Second, the establishment of an international system for standardizing the potency of commercial products. The first generally accepted standard was prepared in France from a fermentation of H-type Bt and called “E-61” (Burges, 1967). This was later replaced by HD-1 as the North American standard based on larvae of the cabbage looper, Trichoplusia ni Hüb (Lepidoptera:Noctuidae) (Beegle et al., 1986). The activity of Bt products is now routinely expressed as the number of International Units (IU) per unit product. Bonnefoi and Debarjac (1963) named as isolate as B. thuringiensis var. aizawai (Bta), which was particularly active against larvae of the wax moth, Galleria mellonella L. (Lepidoptera: Pyralidae) and Spodoptera spp. (Lepidoptera:Noctuidae). Since then several Bt vars. have been found and used against different pests (Beegle and Yamamoto, 1992).
Insecticidal toxins produced by the Bacillus thuringiensis: Bacillus thuringiensis typically produces several types of toxin, four of which are significant (Dulmage, 1970) α-exotoxins (heat-labile exotoxins), β-exotoxins (fly-factor or heat-stable exotoxins), δ-endotoxins (Crystal toxins) (Heimpel, 1August 16, 2002967) and “louse-factor” (Gingrich et al., 1974).
α-exotoxins: Toumanoff (1954) was the first to report that Bt produced a heat-labile insecticidal exotoxin which was toxic to G. mellonella larvae. While Smirnoff (1964) found a similar substance in Thuricide® filtrates, which was toxic to Lepidoptera, Coleoptera, Diptera, Orthoptera and Hymenoptera. Heimpel (1967) coined the term α-exotoxin for such Bt heat-labile toxins. Krieg (1986) estimated the size of α-exotoxins at 45-50 KDa by gel filtration.
β-exotoxins: These were first discovered by Connel and Richards (1959) and named “β-exotoxin” by Heimpel (1967) and since then defined as an adenine-nucleotide and ATP analogue and given the name “thuringiensin”. These low molecular weight toxins are thermostable and have a broad spectrum of activity killing various lepidopteran dipteran, hymenopterans isopteran nematodes and mites. Gingrich et al. (1992) reported the existence of more than one type of heat tolerant exotoxin. Levinson et al. (1990) confirmed the existence of a second heat tolerant exotoxin and named II β-exotoxin, which is more specific than the type I β-exotoxin and very active against the Colorado potato beetle, Leptinotarsa decemlineata Say. (Coleoptera:Chrysomelidae). The β-exotoxin preferentially inhibits biosynthesis of RNA. (Beegle and Yamamoto, 1992). The products based on these β-exotoxins are also used effectively against several species of red spider mites as well as against larvae of houseflies and blowflies (Beegle and Yamamoto, 1992). The β-exotoxin can cause teratogenic effects and disrupt larval or pupal moulting. In mites, exotoxin has been reported as having a gonadotropic and morphogenetic action similar to that of juvenoids (Petrova, 1987).
A new variant of β-exotoxin has been described from B. thuringiensis var. israelensis by Weiser et al. (1992) and Horak et al. (1996). They found activity of water-soluble metabolites of B. thuringiensis var. israelensis toxic to aquatic molluscs and Trematoda. The toxin was termed “M-exotoxin” for molluscs-active exotoxin.
Enterotoxins: Bacillus thuringiensis isolates have been found to produce B cereus-diarrhoeal-type enterotoxins (Carlson et al., 1994). Bacillus cereus enterotoxins are responsible for symptoms of food poisoning following ingestion of B. cereus. Damgaard et al. (1996) isolated enterotoxin producing strains of B. thuringiensis from various foods. Similarly Te Gizit et al. (1997) reported two enterotoxins producing strains that has previously identified as B. cereus and implicated in incidents of food poisoning.
Vegetative insecticidal proteins: A new class of insecticidal toxins, vegetative insecticidal proteins have been isolated from B. thuringiensis. Vip3A is a noval protein with a wide spectrum of activities against lepidopteran insects and was first reported by Estruch et al. (1996). For example Donovan et al. (2001) demonstrated that an important component of B. thuringiensis insecticidal activity against Spodoptera exigua (Lepidoptera: Noctuidae) is the synthesis of Vip3A protein by B. thuringiensis cells after ingestion of spores and crystal proteins by insect larvae. These proteins had no homology to known proteins and were expressed in vegetative growth stage (Estruch et al., 1996). The midgut epithelium cells of the susceptible insects are the primary target for the Vip3A insecticidal proteins and their subsequent lysis appears to be the primary mechanism of lethality (Yu et al., 1997).
δ-endotoxins (crystal proteins): During sporulation Bt produces parasporal crystalline inclusions bodies of a wide range of morphological types (Meadows et al., 1992). The proteins comprising these crystals account for 20-30% of the total bacterial protein at sporulation (Boucias and Pendland, 1998). These can be classified into six general types: bipyramidal, rhomboid, spherical, rectangular, irregular pointed and irregular spherical (Chilcott and Wigley, 1994). The bipyramidal crystals show a greater frequency of toxicity than all other types and the majority of isolates with lepidopteran activity contain such inclusions (Attathom et al., 1995). The crystal (Cry) proteins that form bipyramidal crystals are typically 130 KDA in size and several closely related proteins may be present in a single crystal (Yamamoto and Powell, 1993). Bipyramidal crystals synthesized in the host cells are typically about 1.1 μm long and 0.5 μm wide (Oeda et al., 1989). Irregular spherical crystals can be mosquitocidal (Ohba et al., 1995), while rhomboid crystals are active against some coleopteran species (Burtseva et al., 1995). In addition to producing Cry toxins, several Bt strains also produce cytolytic endotoxins. These include the Cyt1A toxins from Bt israelensis and Bt morrisoni PG14 and Cyt2A from Bt kyushuensis. Unlike the Cry endotoxins, the Cyt endotoxins display broad unspecific activity in vitro and vivo. Crickmore et al. (1998) defined Cry proteins as “a parasporal inclusion (Crystal) protein from Bt that exhibit some experimentally verifiable toxic effect to a target organism, or any protein that has obvious sequence similarity to a known Cry protein”.
The inclusion bodies act as gut poisons to species belonging to six orders of insect: Lepidoptera, Diptera, Coleoptera, Hymenoptera, Homoptera and Mallophaga (Höfte and Whitely, 1989; Feitelson et al.,1992) as well as to Nematoda and Protozoa (Feitelson et al., 1992). A total of 89 different crystal proteins have been cloned (De Maagd et al., 2001). The full list of delta-endotoxin is maintained by Dr N. Crickmore, University of Sussex UK and can be found at http://www.biols.susx.ac.uk/home/Neil_Crickmore/Bt/.
Mode of action of Cry proteins: Bacillus thuringiensis crystals ingested by a susceptible insect larva liberate 130-140 KDa protoxin molecules that are solubilized in the high pH (8.0-10.0) environment of the lepidopteran and dipteran midgut (Koller et al., 1992), or below pH 4 and above pH 10 in the case of the coleopteran-active toxin, Cry3A (Koller et al., 1992). Insecticidal activity decreases rapidly following exposure of protoxins to conditions below pH 2 or above pH 11 (Tran et al., 2001). By the action of midgut proteases these protoxins are processed into active, toxic fragments (approximately 55-70 KDa) (Schnepf et al., 1998), which either act at the midgut membrane or pass through the gut into the haemocoel where it exerts its effect (Fast, 1981). The major proteases in the lepidopteran midgut are trypsin-like (Milne and Kaplan, 1993) or chymotrypsin-like (Novillo et al., 1997). The conversion of Cry1A-type protoxin involves the removal of approximately 600 amino acid residues from the C-terminal end of the molecule, producing an active 67 KDa toxin (Aronson et al., 1991; Visser et al., 1993; Bravo et al., 2002). The earliest physiological symptom of protoxin is an increase in glucose uptake by gut epithelial cells due to a stimulation of respiration (Faust et al., 1974).
Differences in the extent of solubilization of protoxin can sometimes explain differences in the degree of the toxicity between Cry proteins (Du et al., 1994). A reduction in solubility is also thought to be one potential mechanism for insect resistance (McGaughey and Whalon, 1992). However, the host specificity of Bt toxins depends largely upon their ability to bind to a specific midgut receptors (Estada and Ferre, 1994; Feldmann et al., 1995). Rie et al. (1989) reported the occurrence of multiple receptors for Cry1A toxins. For example, receptor 1 binds all Cry1A toxins, receptor 2 binds Cry1Ab and Cry1Ac, while receptor 3 binds Cry1Ac only (Yamamoto and Powell, 1993). Cry1Ab and Cry1Ac share the same binding site in T. ni and O. nubilalis, whereas the Cry1Aa binds to a different site (Estada and Ferré, 1994).
In Lymantria dispar (Lepidoptera:Lymantriidae), Cry1Aa shares a binding site with Cry1Ac and in Manduca sexta (Lepidoptera: Sphingidae) it also share binding site with Cry1Ab (Hofmann et al., 1988). However, Cry1Ab shares a common site with Cry1Aa, Cry1Ac and Cry1F in P. xylostella (Ballester et al., 1994). Such heterogeneity, combined with differences in affinity and concentration of binding sites, may to a large extent account for the observed diversity in toxicity spectra. Additional factors which can play a role in determining specificity are protoxin stability (Arvidson et al., 1989), differential solubilization of crystals (Aronson et al., 1991) and subsequent proteolytic processing (Milne et al., 1995).
Different domains of the Cry protein molecule are responsible for the steps of receptor recognition and pore formation. ( Van Frankenhuyzen, 1993). Elucidation of the three dimensional structure of Cry3A (coleopteran-specific), Cry1Aa (lepidopteran-specific) and Cyt2A (mosquito specific) showed that the toxic fragments of Cry proteins are composed of three distinct structural domains. Domain I (a 7-α-helical bundle) has the ability to form pores or channels in the insect epithelial membrane (Thompson et al., 1995). Domain-II (a triple-β-sheet structure) may be responsible for receptor recognition, which has a fold similar to that of the plant lectin jacalin and jacalin is known to bind carbohydrates via exposed loops at the apex of its β-prism fold. (Schnepf et al., 1998). Domain III (a β-sandwich) in which the C-terminal end is buried, may protect the toxic fragment from further degradation during proteolytic processing (Smith and Ellar, 1994). The “Jelly-roll” configuration of Domain III is a relatively common structural feature that has been observed in a number of other proteins (Yamamoto and Powell, 1993). The β-sandwich structure of domain III is thought to play a key role in the biochemistry of the protoxin molecule, protecting the molecule from proteolysis within the gut (Li et al., 1991). Also, the domain-exchange studies have found that the toxicity of a toxin to the insect host followed the movement of domain III, which is prerequisite step in receptor binding (Li et al., 2001).
In contrast to Cry3A and Cry1Aa, Cyt2A consists of a single domain in which two outer layers of α-helix wrap around a mixed β-sheet. Cyt1A is believed to have a similar structure (Schnepf et al., 1998). The β-sheet structure of Cyt2A suggests a pore based on a β-barrel (Li et al., 1996). Chow et al. (1989) have observed that Cyt1A aggregates on the surface of the target cell. While Gazit et al. (1997) found that Cyt1A self-assembles within the membrane and also identified two α-helices (A and C), which are involved in both membrane interaction and intermolecular assembly.
The membrane bound Cry protein toxic fragment induces the formation of a pore, either aspecific or K+- specific, in the midgut epithelial cell membrane. Pores in the plasma membrane disrupt the actively maintained osmotic balance, causing the cells to swell and burst by colloid-osmotic-lysis (Knowles and Ellar, 1987; Visser et al., 1993) due to an alteration in ion flux and inhibition of amino acid transport across the brush border membrane (BBM) (Giordana et al., 1993). Direct permeability assays on cell lines (Knowles and Ellar, 1987) and BBM vesicles (BBMV) (Carrol and Ellar, 1993) showed that a non-selective Cry protein-induced pore was formed of about 0.6 mm radius, permeable to cations, anions and uncharged molecules up to the size of sucrose. A three-dimensional model of the Cry3A protein supports the hypothesis that the toxin causes the formation of pores or channels in the lipid bilayer (Li et al., 1991).
Binding of the toxin is a two stage process involving reversible (Hofmann et al., 1988) and irreversible phases (Mohan et al., 1995). Irreversible binding is thought to involve tight binding between the Cry protein and receptor and insertion of the former into the apical membrane or both. For example, a truncated Cry1Ab molecule containing only Domain II and Domain III can still bind to midgut receptors but only reversibly, supporting the notion that irreversible binding requires the insertion of Domain I (Flores et al., 1997). The changes in the gut result in vegetative propagation of normal gut microbial flora or of introduced microorganisms, which cause the larvae to stop feeding due to “extreme digestive discomfort” within as little as 2 minutes. This eventually results in a septicaemia which can contribute to or cause death of the larvae (Fast, 1981). Death caused by the bacterial septicemia usually occurs 2-3 days post-ingestion (Bauer, 1995). A proposed mode of action of Bt toxins has been shown (Fig. 1).
Bacillus thuringiensis and transgenic crops: Since the first transgenic plants appeared in the early 1980s (Horsch et al., 1984), there has been very rapid progress directed at using this new technology for the practical ends of crop improvement. Protection of crops from insect pests was quickly seized upon as a major goal of plant genetic engineering (Hilder and Boulter, 1999).
The development of transgenic crop plants expressing Cry proteins (toxins) from Bt has provided new options in the integrated pest management of a wide range of insect pest species, being compatible with biological methods of pest control due to the selective toxicity of the Bt toxins and their methods of delivery to the target pest (Wearing and Hokkanen, 1994). Cotton, maize and potato varieties engineered with Cry genes are fully approved for commercial use in various countries. The number of countries growing transgenic crops commercially has increased from 1 in 1992 to 13 in 1999 (Shelton et al., 2002).
The more obvious advantages of engineered insect resistance in plants are protection of target tissue, weather independent protection, greater stability of Cry toxins, compared with conventional spray application of Bt products and, thus, lack of dependence on application timing for treatment success and protection is possible for the entire season and only those insects which feed on transgenic crops will be exposed (Ely, 1993).
The fast increase in the prevailing acreage of transgenic plants by more than 25 fold from 1.7 million ha in 1996 to 44.2 million ha in 2000 is expected to increase and global market is projected to move from less than US $ 500 million in 1996 to US $25 billion in 2010 (James, 1997; Frutos et al., 1999; Shelton et al., 2002).
However, a major concern is the potential vulnerability of Bt crops to the evolution of resistance by insect pests (Tabashnik et al., 1997a; Wolfenbarger and Phifer, 2000). Large-scale cultivation of Bt transgenic crops will certainly impose selection pressure for pre-existing Bt-resistant insects to increase their numbers. Transgenic plant resistance with Bt genes may thus be a short-lived phenomenon rapidly overcome by insects in much the same way that insecticide toxicity has often been overcome (van Emden, 1999).
Bacillus thuringiensis and non-target insects: Short term risks to natural enemies is a function of the intrinsic susceptibility of the organism and the level of exposure to the toxin (Jepson et al., 1994). Laboratory tests against various invertebrates indicate that Bt has limited impacts on non-target organisms. However, given the wide host range of many Bt strains, non-target impacts of Bt application can occur. For example larvae of the monarch butterfly, Danaus plexippus are susceptible to Cry1Ab (Losey et al., 1999; Jesse and Obrycki, 2000; Hellmich et al., 2001). However, experiments investigating the effects of conventional Bt sprays on other non-target lepidopterans have shown that it is difficult to generalize about susceptibility to Bt and susceptibility must be dealt with on a species by species basis (Peacock et al., 1998).
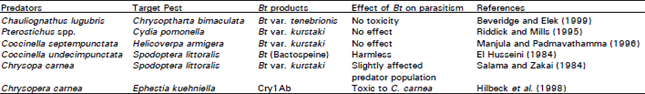
Insect natural enemies are important control agents of insect pests. They are often generalists and will attack several insect pests. Control measure which reduce natural enemy populations may, therefore, result in outbreaks in non-target pests. Bacillus thuringiensis has rarely been found to be toxic to natural enemies (Johnson and Gould, 1992, Hoy, 1998). However some laboratory studies have reported negative effects of Bt toxins on natural enemies. For example Cry1Ab increased mortality of the predatory Lacewing, Chrysoperal carnea Stephens (Neuroptera: Chrysopidae) when its larvae were provided with Cry1Ab fed prey (Hilbeck et al., 1998a) as well as Cry1Ac directly in an artificial diet (Hilbeck et al., 1998b). However other studies with Chrysopa spp., including use of exotoxin-containing products (Zuo et al., 1994), Bt kurstaki (Salama and Zaki, 1984) and Bt tenebrionis (Langenbruch, 1992) did not report significant effects. Some examples of effects of Bt on insect predators are given in Table 1.
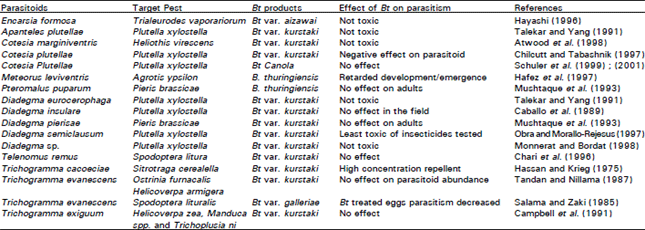
As with predators a range of effects on parasitoids have been recorded, with several studies finding no impact. For example Schuler et al. (1999, 2001) found no detrimental effects of a Bt transgenic oilseed rape line expressing Cry1Ac on the ability of the parasitoid Cotesia plutellae (Hymenoptera:Braconidae) to control P. xylostella. However, treatment of hosts with Bt may adversely affect larval parasitoids by increasing mortality of larval parasitoids within hosts which survived the treatment and by reducing the body size of adult parasitoids (Monnerat and Bordat, 1998). If Bt kills the host too quickly the parasitoid does not have sufficient resources to develop. For example, in a parasitoid of spruce budworm, Apanteles fumiferanae (Hymenoptera:Braconidae) populations were reduced by 50-60% because of lack of parasitoid emergence before host death (Nealis and Van Frankenhuyzen, 1990). Similarly, A. glomeratus, A. melanoscelus and A. litae showed reduced adult life-span after Bt kurstaki application against the host (Salama et al., 1996). The effects of Bt on parasitoids are summarized in Table 2.
Effect of Bacillus thuringiensis on humans: Bacillus thuringiensis has been used for over 60 years for insect pest control but there have been very few reports of clinical infection caused by the organism (Siegel, 2001). However, it has been suggested that the low number of reported cases may be an underestimate due to inadequate diagnostic laboratory facilities, failure to identify Bacillus isolates to species, the mixed microbiological nature of some clinical specimens and the rejection of clinically significant isolates as contaminants. For example, Jackson et al. (1995) reported that Bt isolates recovered from patients with burn wounds were initially incorrectly identified as B. cereus. There are two incidence of reported allergic reaction have been found (McClintock et al., 1995). In the first case it was concluded the individual was most likely, suffering from a previously diagnosed disease while the second case involved an individual with a previous history of life threatening food allergies.
Proven cases that Bt causes clinical disease in mammals remain extremely rare and the risk to public health from Bt is considered to be extremely small (Drobniewski, 1993). Similarly, all mammalian toxicity testing of individual Cry toxins expressed in Bt plants has proved negative. Since Bt affects insects through unique receptor sites in the cell membranes of the insect gut, there are no known receptor sites in mammalian species which could be affected. Thus, there is a reasonable certainty that no harm will result to mammals from dietary exposure to residues of Bt (Anonymous, 1997). It has also been shown that Cry proteins rapidly degrade in vitro, usually within 30 s. Cry proteins range in size from approximately 60 to 120 KDa (Schnepf et al., 1998). These proteins degrade in simulated digestion models to polypeptides of less than 2 KDa (less than 10 amino acids in length). These in vitro models are significantly less robust than the gastrointestinal systems of humans or other animals, which suggests that the Cry proteins will be rapidly and extensively degraded upon consumption (Betz et al., 2000).
Resistance to Bacillus thuringiensis: Resistance is a complex genetic, evolutionary and ecological phenomenon (Metcalfe, 1989) and generally occurs by selection of rare individuals in a population that can survive the insecticide. It is a function of survival of the fittest. Homozygous resistant genes are rare in untreated populations compared with heterozygotes and homozygous susceptible (Dent, 2000). Widespread application of pesticides propagates resistant alleles through preferential survival either by selection or random mutation. Continued selection pressure in the same direction will lead to these alleles being essential in the population for offspring production (Sutherst and Comins, 1979).
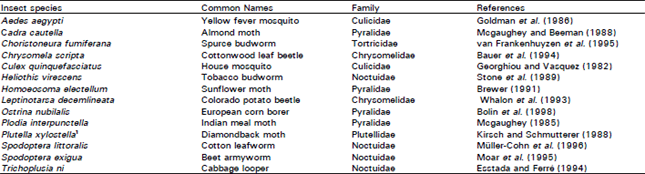
Commercial Bt formulations had been used in the field for more than three decades before resistance appeared (Tabashnik, 1994). The eventual development of Bt resistance in populations of P. xylostella was perhaps inevitable, however, given its biology, its history of insecticide resistance and the very high frequency of Bt applications applied to high value crucifer crops to control this pest in the late 1980s. In the laboratory, a total of 14 species have developed resistance to Bt Cry toxins (Table 3) (Ferré and van Rie, 2002).
Insects with a large number of generations per annum will tend to show a more rapid development of resistance than insects with only one generation in a growing season (Georghiou and Taylor, 1986). In a population with a low influx of genes resistance may become rapidly fixed while in populations with a continuous influx of novel genes resistance may be continuously diluted (Peferoen, 1997). The way insecticides are applied also has a dramatic effect on the rate of resistance development. Topical application of Bt compared with expression of a Bt Cry protein in a plant imposes different selection pressure on an insect population.
Bacillus thuringiensis resistance mechanisms: The mechanism by which an insect evolves resistance to a particular toxin is unavoidably related to the toxin’s mode of action (Gill et al., 1992; Marrone and Macintosh, 1993). Although several different resistance mechanisms have been proposed (Gill et al., 1992), the most likely ones to date involve changes in Bt receptors or solubilization-activation of the crystal proteins (Ferré and van Rie, 2002). Receptor mediated mechanism may include loss of Cry toxin binding sites, increase in non-specific binding not related to toxicity. While solubilization and proteinase mediated resistance mechanism could involve changes in gut pH or in proteinases involved in protoxin activation.
Receptor-mediated mechanisms: Studies on various lepidopteran species have suggested that toxin binding to midgut receptors is responsible for toxin sensitivity or specificity among several different species (Ballester et al., 1994). In this type of resistance, the protoxin is solubilized and activated in the resistant population as it is in susceptible insects. However, the activated toxin can no longer recognize a suitable binding site or epitope on the receptor (Rie et al., 1990; Ferré et al., 1991). The first case of resistance associated with altered midgut binding of a toxin was reported in P. interpunctella (Rie et al., 1990). The resistant strain of P. interpunctella showed reduction in binding affinity but no change in the number of binding sites of Cry1Ab. This indicated that alteration in the binding site for Cry1Ab was preventing effective binding of the toxin.
The examination of Cry toxin binding in P. xylostella revealed loss in specific binding to Cry1Ab, suggesting that resistance was also due to a change in the Cry1Ab binding site (Ferré et al., 1991). A Hawaiian colony of P. xylostella that was resistant to all three Cry1A toxins was found to exhibit reduced binding of Cry1Ac in BBMV assays (Tabashnik et al., 1994a) and rapid reversal of resistance was associated with the restoration of the binding properties of Cry1Ac (Tabashnik et al., 1994b). Similarly, a population of P. xylostella from Malaysia highly resistant to Cry1Ac and moderately to Cry1Ab exhibited reduced binding to both toxins in BBMV assays and reversal of resistance was associated with the restoration of binding properties of the toxins (Sayyed et al., 2000a).
Solubilization/proteinase-mediated mechanisms: Proteolytic activation of Cry1 toxins involves the removal of peptide sequences from both the N- and C- termini of the protoxin (Bravo et al., 2002). Resistance could thus involve a decrease in proteolytic properties of the insect midgut (Marrone and Macintosh, 1993).
Since proteolytic enzymes are involved in the dissolution and activation of Bt protoxins (Dai and Gill, 1993; Milne and Kaplan, 1993), the first evidence for protease involvement in Bt resistance was found in a Bt resistant strain of P. interpunctella that displayed a slower processing of protoxins than in the susceptible strain and activation of Cry1 protoxins with midgut enzymes resulted in proteins of intermediate size of 60 KDa proteins (Oppert et al., 1994). In this strain of P. interpunctella, the major serine proteinase was absent. Since these proteinases are involved in the activation of Bt protoxin (Oppert et al., 1996), lack of critical Bt activating enzymes could contribute to toxin resistance. Similarly, two other resistant strains of P. interpunctella were found to have a genetically-linked reduced protoxin activation compared with a susceptible strain due to the absence of major trypsin-like gut enzyme (Oppert et al., 1997). In the CP73-3 colony of Heliothis virescens (Lepidoptera:Noctuidae) slower activation of Cry1Ab protoxin and faster degradation of Cry1Ab toxin in midgut extract was observed by Forcada et al. (1996). While Inagaki et al. (1992) found that complete degradation of Cry1Ab by proteases derived from the resistant Spodoptera litura (Lepidoptera:Noctuidae) was the likely cause of resistance. Similarly, Keller et al. (1996) suggested that reduced sensitivity of fifth-instar larvae of S. littoralis to Cry1C could be attributed to increased degradation of the toxin by proteases in the less susceptible larvae. Milne et al. (1995) reported that a protein complex present in the midgut of spruce budworm, Choristoneura fumiferana (Lepidoptera:Tortricide) could inactivate Cry1Aa by precipitation followed by proteolysis, thus accounting for resistance to this toxin.
| Table 1: | The effects of Bacillus thuringiensis on parasitoids of different insect pest species |
 | |
| Table 2: | Effect of Bacillus thuringiensis on predators of different insect pest species |
 | |
| Table 3: | Species of insects selected in the laboratory or field1 for resistance to Bacillus thuringiensis |
 | |
| Table 4: | Cross-resistance pattern to Bacillus thuringiensis and its toxins in different insect pest species |
 | |
However administration of serine protease inhibitors with protoxin in a susceptible strain of P. xylostella did not lead to a decrease in toxicity, which suggested that the proteolytic enzymes involved in the activation of endotoxins in this population belonged to a different group of proteases (Tabashnik et al., 1992a).
Cross-resistance patterns: Cross-resistance patterns can help to identify resistance mechanisms (Roush and Tabashnik, 1990). reported for a number of toxins (Table 4). Some insect species Generally the incidence of cross-resistance is low but it has been can be readily selected for resistance to several different Bt toxins (Mcgaughey and Johnson, 1994). For example, it has been shown that P. interpunctella can be selected for resistance to Cry1Aa, Cry1Ab, Cry1Ca and Cry1Da and possibly other Cry toxins contained in B. thuringiensis var. aizawai (Bta) (McGaughey and Johnson, 1994). Cross-resistance between Bt toxins has also been reported in H. virescens (Gould et al., 1992; Gould et al., 1995) and P. xylostella (Tabashnik et al., 1994c; Wright et al., 1997; Sayyed et al., 2000a, Sayyed and Wright, 2001a). Cross-resistance among Cry1A toxins is not surprising, owing to their structural and functional similarities (Shelton et al., 1993) and studies have shown that these toxins may bind to the same receptor in most of the insect species tested (Ballester et al., 1999).
Several studies have reported an absence of cross-resistance between synthetic insecticides and Bt. For example Bt resistant P. xylostella showed no cross-resistance to phenthoate, fenvalerate, chlrofluazuron or abamectin (Iqbal et al., 1996; Sarnthoy et al., 1997). However, resistance to Btk in the sheep lice, Bovicola (Damalinia) ovis Schrank (Phthiraptera: Trichodectidae) was shown to be inversely related to pyrethroid resistance (Drummond et al., 1995).
Cross-resistance patterns and their underlying physiological mechanism are very complex and somewhat unpredictable, even in closely related groups of toxins and insects (Table 4; Tabashnik, 1994). Where resistance is due to reduced binding, then cross-resistance can be predicted from competitive binding studies with susceptible insects, however when the resistance is unrelated to binding, then this can produce unexpected results such as the broad cross-resistance as in H. virescens (Gould et al., 1995) and P. xylostella (Sayyed et al., 2000a, Sayyed and Wright, 2001a).
Multiple mechanisms of resistance: These evolve as a consequence of serial exposure to different toxins or combinations of toxins (Wright et al., 1997; Frutos et al., 1999). For example, Gould et al. (1992) reported that a strain of H. virescens selected for Cry1Ac also showed resistance to Cry1Ab and Cry2Aa. Binding analyses showed that there was no change in the binding characteristics of Cry1Ab and Cry1Ac and although the resistant strain seemed to have fewer receptors for Cry1Ac, these were of high affinity. There was no significant decrease in the affinity or number of binding sites which could explain the level of resistance which was therefore considered to occur post-binding (Moar et al., 1995). In a S. littoralis population, experiments suggested the low level of activity of Cry1A toxins was due to an inability to carry out the pore formation step (Escriche et al., 1998). Similarly, a P. xylostella population (SERD3) from Malaysia resistant to both Btk and Bta showed decreased binding to Cry1Ab but not to Cry1Aa, Cry1Ac or Cry1Ca, suggesting that reduced toxin binding alone could not account for the resistance observed (Wright et al., 1997). In another population (SERD5), collected from the same location, reduced activation of protoxin is thought to be a major mechanism (Sayyed et al., 2001b).
Genetics of Bacillus thuringiensis resistance: The genetic basis of resistance is complex (Heckel, 1994). It includes the initial frequency of resistance alleles, natural variability, mode of inheritance (degree of dominance, sex linkage) and fitness costs associated with resistance.
Initial frequency of resistance alleles: Although such information is useful for assessing the evolution of resistance, it is not easily available (Frutos et al., 1999) and the assessment of the initial frequency of resistance alleles is almost never conducted prior to development of resistance (Tabashnik et al., 1990; Gould et al., 1997).
The initial frequency of resistance alleles influences the rate at which resistance will evolve (Alstad and Andow, 1995; Gould et al., 1997). Change in allele frequency is the key indicator of the effectiveness of an insecticide. The initial allelic frequency is generally assumed to range from 10–6 to 10–3 (Roush and McKenzie, 1987; Gould et al., 1998). A direct approach, making use of a homozygous recessive resistant strain, has estimated the frequency of a major Bt resistance allele in a field population of H. virescens to be 1.5 x 10–3 (Gould et al., 1997). By applying the same approach in P. xylostella, Tabashnik et al. (1997a) estimated the frequency of a recessive Bt resistance allele in a susceptible laboratory population to be 1.2 x 10–1. While Andow and Alstad (1996) used a F2 screening procedure to estimate the frequency of Cry1Ab resistance alleles as <1.3 x 10–2 in a laboratory population of O. nubilis. Using a slightly modified procedure Andow et al. (2000) and Bentur et al. (2000) estimated the initial frequency of resistance in an Iowa population of Ostrinia nubilalis (Lepidoptera:Pyrallidae) to Bt corn <3.9 x 10–3 and <3.6 x 10–3 respectively. Using a new approach, Tabashnik et al. (2000) estimated initial frequency of resistance in a field population of P. gossypiella in Arizona to be 1.6 x 10–1.
Natural variation in susceptibility to Bt: Intra specific variation in susceptibility to Bt between different geographical populations has been reported for various insect species. For example, a population of rice stripped stemborer, Chilo suppressalis Walk. (Lepidoptera: Pyralidae) from Southeast Asia was susceptible to Cry1Ca (Lee et al., 1997) whereas a population from Southern France was not (Fiuza et al., 1996). Similar results were reported for O. nubilalis collected from different places of USA (Huang et al., 1997). However, the spruce budworm, Choristoneura fumiferana Clem (Lepidoptera: Tortricidae) showed limited geographical variation in susceptibility to Bt although intra-population variability showed a 2 to 30 fold difference between the highest and lowest percent mortalities at a single dose (Van Frankenhuyzen et al., 1995). In general, the susceptibility of Cry proteins can vary among different populations of a given insect species (Cabrera et al., 2001). This could affect the standardisation of potency of B. thuringiensis based products and make estimation of resistance levels in populations exposed to Cry toxins more difficult.
Another way to estimate the variability in resistance genes is to measure heritability (h2). Tabashnik (1994) estimated heritability of resistance to Btk and Cry1A toxins and showed that compared with eight other insects species, P. interpunctella had relatively high h2 values, indicating low phenotypic variation. However, relatively higher h2 values were found in two populations of P. xylostella from Malaysia (Iqbal et al., 1996; Wright et al., 1997; Sayyed et al., 2000a reflecting a low phenotypic variation probably resulting from continuous exposure to Bt.
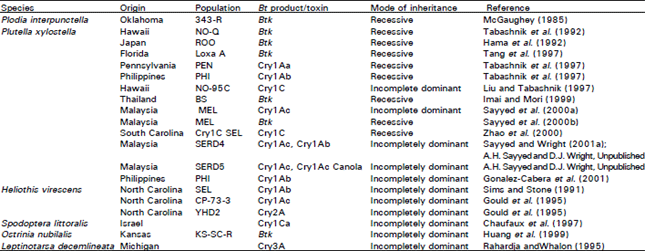
Mode of inheritance of resistance: Information on the mode of inheritance can improve resistance monitoring, risk assessment, modelling and resistance management (Tabashnik et al., 1992b). The inheritance of Bt resistance in insects has commonly been assumed to be recessive in nature. The mode of inheritance of resistance in various insect populations is summarised in Table 5.
The single backcross method has been used to determine the number of loci involved in resistance. This has shown that the backcross data fitted fairly well to a single locus model (Ferré and van Rie, 2002). However resistance to Cry1Ca and Cry1Ac in Florida and Melaka populations of P. xylostella (Zhao et al., 2000; Sayyed et al., 2000a, Sayyed and Wright, 2001a), to Cry1Ab in a North Carolina population of H. virescens (Sims and Stone, 1991), to Cry1Ca in a population of S. littoralis from Israel (Chaufaux et al., 1997) and to Cry3Aa in a Cry3A-selected laboratory population of L. decemlineata (Rahardja and Whalon, 1995) was found to be controlled by the combined interaction of resistance alleles from the various loci.
It has been shown consistently that the resistance to B. thuringiensis is autosomally inherited. However, in some cases the sex of the resistant parent has a significant influence on inheritance of resistance. For example, resistance to Cry1C in a S. littoralis population (Chaufaux et al., 1997), to Cry1Ab in BL and to Cry1Ac in SERD4 populations P. xylostella had some maternal influence (Real et al., 1995; Sayyed and Wright, 2001a). In contrast, a Cry1Ab-selected population of P. xylostella exhibited some paternal influence.
At present, the evidence suggests that partial or completely recessive modes of inheritance (Table 5) are consistently associated with modification of binding sites and therefore altered specific binding (Liu and Tabashnik, 1997a; Gould et al., 1997). Whereas more dominant alleles seem to be associated with other resistance mechanisms, conferring more broad spectrum resistance (Tabashnik et al., 1998). For example, in the SERD5 population of P. xylostella resistance was incompletely dominant (Sayyed and Wright, unpublished data) and reduced protoxin activation is suggested to be the major mechanism of resistance to Cry1Ac (Sayyed et al., 2001b).
| Table 5: | Inheritance of resistance to Bt products and Cry toxins in different insect pest populations |
 | |
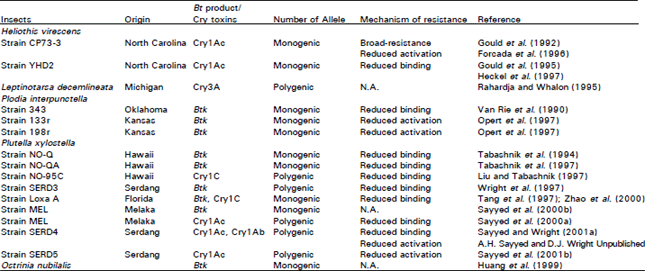
| Table 6: | Mechanism of resistance to Bt products and Cry toxins in different insect pest populations |
 | |
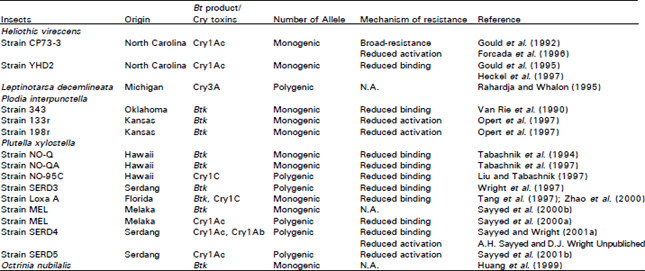
| Table 7: | Fitness costs and reversion of resistance to Bt products and Cry toxins in different insect pest populations |
 | |
Number of resistance alleles: Theoretical models describing the evolution of resistance in various insect population have been developed based on a monogenic mode of inheritance (Ferré and van Rie, 2002). However, results obtained by using different methods indicate that insects may have more than one factors conferring resistance to Cry toxins. For example, in backcross experiments results showed that Bt resistance populations did not fit a monofactorial model (Rahardja and Whalon, 1995; Liu and Tabashnik, 1997a; Sayyed et al., 2000a; Sayyed and Wright, 2001a) (Table 6). The results of different approaches used to determine the number of resistance alleles are summarised (Table 6).
Genetic analysis using isozyme polymorphism has also been used to identify the number of factors conferring resistance. For example, a backcross design with 10 marker loci in the YHD2 strain of H. virescens revealed the existence of a major locus named BtR-4 on linkage group 9, responsible for as much as 80% of the total resistance to Cry1Ac (Heckel et al., 1997).
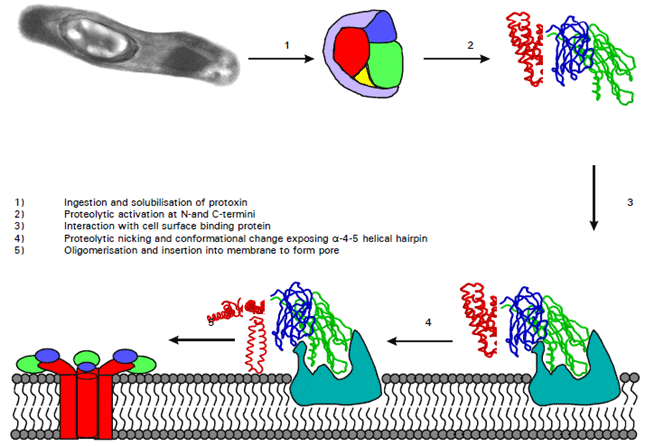 | |
| Fig. 1: | Proposed mechanism of action of Bt toxin (After De Maagd et al., 2001) |
Using the isozyme-linkage method, a strong correlation was found between Cry1A resistance and two mannose-6-phosphate isomerase isozymes in the PHI strain of P. xylostella (Herrero et al., 2001).
Reversion of resistance and fitness costs: Instability of resistance is the tendency for the frequency of resistance genotypes to decrease in a population beyond effects directly attributable immigration or emigration (Tabashnik et al., 1994a). Resistance to insecticides is often accompanied by fitness costs, such as a decreased rate of development, fecundity, survival or mating competitiveness relative to susceptible insects (Roush and McKenzie, 1987). In the majority of cases, resistance to Bt and individual Cry toxins has been found to be unstable. A rapid decline in resistance has been found in three selected strains of P. xylostella from Hawaii with 2800 to 90-fold resistance (Tabashnik et al., 1994a). A fast rate of decline of resistance has also been reported for Btk resistant P. xylostella populations from Japan and Malaysia (Hama et al., 1992; Sayyed et al., 2000b). Such declines in resistance are most likely caused by fitness costs associated with resistance. However, P. interpunctella resistance did not decline in one strain (343-R) even after 29 generations on untreated diet. Similarly, in a P. xylostella population from Hawaii resistance in one isofemale line of the selected strain NO-Y remained stable after 32 generations without exposure (Tabashnik et al., 1995). It has been shown that the decline in resistance in moderately resistant strains of P; xylostella (Tabashnik et al., 1994a), P. interpunctella (McGaughey and Beeman, 1988) and H. virescens (Sims and Stone, 1991) was much slower than in populations with higher levels of resistance. The results of reversion of resistance studies are summarized in Table 7.
High fitness costs associated with evolution of Bt resistance have been reported (Tabashnik 1994; Groeters et al., 1994). However, in other cases few if any fitness costs have been found (McGaughey, 1985; Liu et al., 1996; Tang et al., 1997). For example, there were no differences in fitness between Bt-susceptible and resistant population of H. virescens, when larvae were fed diets that did not contain Bt (Gould and Anderson, 1991). However, there were increases in both development time and mortality for Bt-resistant H. virescens larvae forced to consume Btk. Relative to Bt-susceptible L. decemlineata, larval development was delayed and egg production was decreased in a Bt-resistant population feeding on potato plants not treated with Bt var. tenebrionis (Trisyono and Whalon, 1997; Alyokhin and Ferro, 1999). Also, more Bt-susceptible than Bt-resistant L. decemlineata survived after overwintering in diapause (Alyokhin and Ferro, 1999). While a resistant population of P. gossypiella feeding on transgenic Bt-cotton showed a longer development time than Bt-susceptible bollworms (Liu et al., 1999).
In contrast, two populations of P. xylostella from Malaysia showed shorter development times and greater pupal weight compared with unselected sub-populations in the presence of Cry1Ac (Sayyed and Wright, 2001b). Likewise, H. virescens was reported to show enhanced growth in the presence of Cy1Ab toxin (Gould et al., 1995). Liu et al. (1996) suggested that prolonged selection in the field reduced fitness costs either in substitution of alternate alleles conferring resistance or by selection of fitness modifiers at loci not directly related to resistance. Fitness costs are often difficult to describe owing to their compost nature and dependence on various physiological aspects as well as mechanisms of resistance (Frutos et al.,1999; Sayyed and Wright, 2001b). In addition, genetic drift or mutation can cause instability (Tabashnik et al., 1994a). However, these mechanisms may not be expressed in larger field populations. In addition, diverging opinions exist regarding the impact of fitness cost on delay of resistance. Some consider that fitness costs directly caused by resistance alleles will have important effect in the field (Tabashnik et al., 1994b), whereas other consider that even strong fitness costs will have a minimal impact on evolution of resistance (Roush, 1997). Liu et al. (1996) suggested that stability is not necessarily a fixed trait even for a particular population.
Implications of genetic diversity for resistance management: The main purpose of resistance management is to diminish the selection of initially rare individuals carrying resistance alleles and hence to keep the frequency of resistance alleles sufficiently low for insect control (Schnepf et al., 1998). Strategy development generally relies on theoretical assumptions and on computer models (Tabashnik, 1994; Roush, 1997, 1998). However, a thorough understanding of the genetics of resistance to Cry toxins is essential for the development and maintenance of resistance management strategies (Tabashnik et al., 1991, 1998). It had been widely assumed that resistance to Bt in insects is inherited as a recessive trait (McGaughey, 1985; Tabashnik et al., 1992a; Ferré et al., 1995). However, as we have seen incompletely dominant, polygenic modes of inheritance of resistance to Cry1Ac were present in three field populations of P. xylostella from Malaysia. Some degree of dominance has also been reported in two other field-derived Cry toxin resistant populations of P. xylostella from the Philippines and Hawaii and in laboratory selected populations of at least five other insects species (Table 5). Thus, the monogenic recessive model of Bt resistance in P. xylostella (Tabashnik et al., 1992a) may not necessarily be the most common in the field, with many resistant insect populations having more complex genetics of resistance. There is also evidence that a Cry toxin resistance mechanism (reduced activation) other than reduced binding is of major importance in P. xylostella (Sayyed et al., 2001b).
Various strategies to manage resistance to Bt toxins have been proposed (Tabashnik, 1994; Gould, 1998; Frutos et al., 1999), all of which rely to a greater or lesser extent on resistance being recessive, that the frequency of resistance alleles is low in the field compared with susceptible alleles and that the frequency of resistance alleles will decrease when the selection pressure is released. For example, if resistance is recessive, F1 offspring produced by mating between susceptible and resistant adults are killed by eating Bt plants. If mating is random, initially rare homozygous resistant adults emerging from Bt plants are likely to mate with the more abundant homozygous susceptible adults emerging from non-Bt plants, producing F1 progeny that cannot survive on Bt plants expressing high doses of toxin (Fishhoff, 1996; Anonymous, 1997). Mathematical models and data from laboratory and greenhouse studies indicate that resistance can be delayed substantially when these assumptions are valid (Gould, 1998). However, it is difficult to predict the exact concentration of Cry toxin for incomplete mode of inheritance of resistance. Whereas, Gould et al. (1994) proposed 25 times the concentration needed to kill 99 percent of susceptible insects. Population genetics theory (Tabashnik and Croft, 1982; Gould, 1986; Mallet and Porter, 1992; Alstad and Andow, 1996) and laboratory experiments (Liu and Tabashnik, 1997; Roush, 1998) predict that this approach will substantially delay evolution of resistance, if it is appropriately implemented and its assumptions are met.
The principal methods proposed for Bt resistance management are: (a) mixtures, mosaics or rotations of transgenic plants; (b) time or tissue-specific expression of toxin; (c) low doses of toxin in combination with natural enemies; (d) co-expression of different cry genes; and (e) high expression (dose) with refugia, which is the strategy recommended currently (Shelton et al., 2000; Tang et al., 2001). However, if non-recessive inheritance and multiple mechanisms of resistance to Cry toxins are, in fact, relatively common in field populations of insects, rethinking of resistance management strategies may be required (Tabashnik et al., 1998).
The refuge/high-dose strategy entails high risk because it could greatly accelerate resistance if certain assumptions are not valid. For example, non-random (assortative) mating and movement of adults may lead to failure of this strategy. Resistant larvae of P. gossypiella on Bt cotton have been observed to take long duration to develop, compared with susceptible larvae on non-Bt cotton (Table 7). While Cry1Ac-SEL SERD4 and Cry1Ac-SEL MEL sub-populations of P. xylostella had a shorter development time on Cry1Ac-treated leaves compared with an unselected population on untreated leaves (Table 7). In the above examples assortative mating could occur if local populations of insects were sufficiently synchronised and mating occurred prior to dispersal. While this is perhaps unlikely to be the case with P. xylostella, where overlapping generations is common, such developmental asynchrony could be important in cotton crop pests with non-overlapping generations. If dispersal occurs after mating, immigration of resistant individuals could increase the frequency of heterozygotes in the refugia (Caprio and Tabashnik, 1992).
The refuge/high-dose strategy will also fail if the plants do not produce, or fail to maintain, a dose sufficiently high to kill most heterozygotes. For example in Bt cotton fields in Australia, the damage due to H. armigera was similar to that in refugia (Forrester and Pyke, 1997). Efficacy against Helicoverpa spp. typically declines through the boll maturation period, to the point where survival of larvae is little different to that in refugia although growth rates of survivors on the INGARD Bt crops are still dramatically reduced (Fitt, 2000). Clearly this pattern is not consistent with a high dose strategy and the changing efficacy of Bt cotton imposes additional risks for resistance management. In Australia, Bt cotton expressing two independent Cry toxins (Cry1 Ac and Cry2A) shows much more consistent efficacy compared with Bt cotton expressing Cry1Ac alone and will greatly enhance the sustainability of resistance management (Roush, 1996). Other possibilities for Cry toxins are also being investigated (Llewellyn and Higgins, 1998, Hanzlik and Gordon, 1998).
The refuge in the refuge/high-dose strategy should ideally be maintained free of any treatment with pesticides to ensure the presence of a sufficient number of susceptible adults (Gould, 1998). In fact, Anonymous (1997) recommend the planting of 20% non-Bt plants of cotton that can be treated with a non-Bt foliar insecticide, or a 4% refuge of non-Bt plants that is left untreated. However, Shelton et al. (2000) have shown that the insect population in the sprayed refuge had a significantly lower average mortality at the diagnostic dose for resistance compared with the insects in the unsprayed refuge. Insects collected from the Bt plants would have a resistant genotype for Bt resistance and significantly greater numbers of Bt-resistant larvae were found on the Bt plants when the refuge was sprayed compared with when it was not sprayed. However, if the 20% non-Bt refuge is managed using an IPM strategy in such way that the population of beneficial insects increases this could result in a significant decrease in the pest population (Roush, 1996; Riggin-Bucci and Gould, 1997). Therefore, the use of chemical sprays can be reduced, increasing the effectiveness of the refuge (Gould, 1998).
The multiple toxin strategy is usually considered as an adjunct to the high-dose strategy. Instead of having a high concentration of one toxin, plants express high concentration of two or more toxins. The multiple toxin strategy coupled with a refugia could provide benefits even if a high dose (25 times the LD99 of susceptible insects) is not reached (Gould et al., 1994). However, if the two toxins are each expressed at levels that only kill 50-80% of the insects, this strategy may not be highly effective in slowing the evolution of resistance (Gould et al., 1994).
Cross-resistance between Cry toxins (McGaughey, 1994) and multiple resistance have important implications for Bt resistance management. In the presence of cross-resistance, a two-toxin resistance management strategy (Bt plants expressing two Cry toxins) can fail quickly. However, in the absence of cross-resistance there is a ten-fold advantage of using this strategy (Roush, 1998). The potential of Cyt1A in preventing evolution of resistance in mosquito and P. xylostella larvae exposed to Bti and Btk (Wirth et al., 1997; Sayyed et al., 2001a) and resistance to Cry3A in C. scripta (Federici and Bauer, 1998) has also been reported.
In crops such as cotton, with multiple pests, because of unique biology of each pest it is hard to design a resistance management strategy that is appropriate for all the pest species. A toxin may be highly active against a given pest (e.g., Cry1Ac vs H. virescens) but less active against another present on the same crop (e.g., Cry1Ac vs H. zea or H. armigera). In such a case, the second pest may be exposed to a moderate or weak dose on a Bt crop and may evolve resistance. This leads to use of Bt plants as a component of a comprehensive IPM approach (Hoy, 1998). The use of Bt-based formulations within an IPM program was shown to be effective for controlling pests (Trumble et al., 1994; Meade and Hare, 1995). IPM can delay resistance by providing multiple sources of mortality (Denoholm and Rowland, 1992). For example, the use resistant cabbage cultivars will not only slower the development of P. xylostella but the synergistic interaction can also lead to more successful control by Bt as well as an improved efficacy of natural enemies (Schuler and van Emden, 2000).
As there is no single answer or strategy to delay resistance, only sound and flexible methods of pest control will provide sustainability. The refuge/high-dose strategy is currently the most promising approach, however the strategy is based on assumptions that as we have discussed may not always apply. Continued studies on the genetic diversity of Bt resistance in different insect populations are required and the susceptibility of pest populations must continue to be monitored to evaluate the success of resistance management plans.
REFERENCES
- Betz, F.S., B.G. Hammond and R.L. Fuchs, 2000. Safety and advantages of Bacillus thuringiensis-protected plants to control insect pests. Regular Toxicol. Pharmacol., 34: 156-173.
CrossRefDirect Link - Burtseva, L.I., V.A. Burlak, G.V. Kalmikova, H. de Barjac and M.M. Lecadet, 1995. Bacillus thuringiensis novosibirsk (Serovar H24a24c), a new subspecies from the West Siberian plain. J. Invertebrate Pathol., 66: 92-93.
CrossRefDirect Link - Crickmore, N., D.R. Zeigler, J. Feitelson, E. Schnepf and J. Van Rie et al., 1998. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev., 62: 807-813.
Direct Link - Ferre, J. and J. Van-Rie, 2002. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol., 47: 501-533.
PubMed - Alyokhin, A.V. and D.N. Ferro, 1999. Relative fitness of colorado potato beetle (Coleoptera: Chrysomelidae) resistant and susceptible to the Bacillus thuringiensis Cry3A toxin. J. Econ. Entomol., 92: 510-515.
CrossRefDirect Link - Alstad, D.N. and D.A. Andow, 1995. Managing the evolution of insect resistance to transgenic plants. Science, 268: 1894-1896.
CrossRefDirect Link - Aronson, A.I., E.S. Han, W. McGaughey and D. Johnson, 1991. The solubility of inclusion proteins from Bacillus thuringiensis is dependent upon protoxin composition and is a factor in toxicity to insects. Applied Environ. Microbiol., 57: 981-986.
PubMedDirect Link - Arvidson, H., P.F. Dunn, S. Strnad and A.I. Aronson, 1989. Specificity of Bacillus thuringiensis for lepidopteran larvae: Factors involved in vivo and in the structure of a purified protoxin. Mol. Microbiol., 3: 1533-1543.
CrossRefPubMedDirect Link - Attathom, T., W. Chongrattanameteekul, J. Chanpaisang and R. Siriyan, 1995. Morphological diversity and toxicity of delta-endotoxin produced by various strains of Bacillus thuringiensis. Bull. Entomol. Res., 85: 167-173.
CrossRefDirect Link - Ballester, V., B. Escriche, J.L. Mensua, G.W. Riethmacher and J. Ferre, 1994. Lack of cross‐resistance to other Bacillus thuringiensis crystal proteins in a population of Plutella xylostella highly resistant to cryia(b). Biocontrol Sci. Technol., 4: 437-443.
CrossRefDirect Link - Ballester, V., F. Granero, B.E. Tabashnik, T. Malvar and J. Ferre, 1999. Integrative model for binding of Bacillus thuringiensis toxins in susceptible and resistant larvae of the diamondback moth (Plutella xylostella). Applied Environ. Microbiol., 65: 1413-1419.
PubMedDirect Link - Bauer, L.S., 1995. Resistance: A threat to the insecticidal crystal proteins of Bacillus thuringiensis. Florida Entomol., 78: 414-443.
Direct Link - Beegle, C.C. and T. Yamamoto, 1992. Invitation paper (C.P. Alexander Fund): History of Bacillus thuringiensis berliner research and development. Can. Entomol., 124: 587-616.
CrossRefDirect Link - Beegle, C.C., T.L. Couch, R.T. Alls, P.L. Versoi and B.L. Lee, 1986. Standardization of HD-1-S-1980: U.S. standard for assay of lepidopterous-active Bacillus thuringiensis. Am. Entomol., 32: 44-45.
CrossRefDirect Link - Bentur, J.S., D.A. Andow, M.B. Cohen, A.M. Romena and F. Gould, 2000. Frequency of alleles conferring resistance to a Bacillus thuringiensis toxin in a Philippine population of Scirpophaga incertulas (Lepidoptera: Pyralidae). J. Econ. Entomol., 93: 1515-1521.
CrossRefPubMedDirect Link - Bravo, A., R. Miranda, I. Gomez and M. Soberon, 2002. Pore formation activity of Cry1Ab toxin from Bacillus thuringiensis in an improved membrane vesicle preparation from Manduca sexta midgut cell microvilli. Biochim. Biophys. Acta (BBA)-Biomembranes, 1562: 63-69.
CrossRefPubMedDirect Link - De Maagd, R.A., A. Bravo and N. Crickmore, 2001. How Bacillus thuringiensis has evolved specific toxins to colonize the insect world. Tren. Genet., 17: 193-199.
PubMed - Estruch, J.J., G.W. Warren, M.A. Mullins, G.J. Nye, J.A. Craig and M.G. Koziel, 1996. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA., 93: 5389-5394.
PubMed - Ferre, J., M.D. Real, J.V. Rie, S. Jansens and M. Peferoen, 1991. Resistance to the Bacillus thuringiensis bioinsecticide in a field population of Plutella xylostellaL. is due to change in the midgut receptor. Proc. Natl. Acad. Sci. USA., 88: 5119-5123.
Direct Link - Gill, S.S., E.A. Cowles and F.V. Pictrantonio, 1992. The mode of action of Bacillus thuringiensis endotoxins. Ann. Rev. Entomol., 37: 615-634.
Direct Link - Gonzalez-Cabrera, J., S. Herrero, A.H. Sayyed, B. Escriche and Y.B. Liu et al., 2001. Variation in susceptibility to Bacillus thuringiensis toxins among unselected strains of Plutella xylostella. Applied Environ. Microbiol., 67: 4610-4613.
CrossRefPubMedDirect Link - Gould, F., 1998. Sustainability of transgenic insecticidal cultivars: integrating pest genetics and ecology. Ann. Rev. Entomol., 43: 701-726.
CrossRefDirect Link - Gould, F., A.M. Ramirez, A. Anderson, J. Ferre, F.J. Silva and W.J. Moar, 1992. Broad-spectrum resistance to Bacillus thuringiensis toxins in Heliothis virescens. Proc. Natl. Acad. Sci. USA., 80: 7986-7990.
Direct Link - Hilbeck, A., W.J. Moar, M.P. Carey, A. Filippini and F. Bigler, 1998. Toxicity of Bacillus thuringiensis Cry1Ab toxin to the predator Chrysoperla carnea (Neuroptera: Chrysopidae). Environ. Entomol., 27: 1255-1263.
Direct Link - Hilder, V.A. and D. Boulter, 1999. Genetic engineering of crop plants for insect resistance-a critical review. Crop Prot., 18: 177-191.
CrossRefDirect Link - Hofte, H. and H.R. Whiteley, 1989. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev., 53: 242-255.
Direct Link - Jackson, S.G., R.B.G. Brand, R. Ahmed and S. Kasatiya, 1995. Bacillus cereus and Bacillus thuringiensis isolated in a gastroenteritis outbreak investigation. Lett. Applied Microbiol., 21: 103-105.
PubMed - Knowles, B.H. and D.J. Ellar, 1987. Colloid-osmotic lysis is a general feature of the mechanism of action of Bacillus thuringiensis δ-endotoxins with different insect specificity. Biochim. Biophys. Acta Gen. Subj., 924: 509-518.
CrossRefDirect Link - Llewellyn, D.J. and T.J. Higgins, 1998. Biotechnological Approaches to Crop Protection Novel Sources of Insect Tolerance Genes other than Bt-Toxins. In: Pest Management Future Challenges, Zalucki, M.P., R.A.I. Drew and G.G. White (Eds.). University of Queensland Press, Brisbane, Australia, pp: 223-229.
- Li, J., T.J. Carroll and D.J. Ellar, 1991. Crystal structure of insecticidal δ-endotoxin from Bacillus thuringiensis at 2.5 A resolution. Nature, 353: 815-821.
CrossRefDirect Link - Liu, Y.B. and B.E. Tabashnik, 1997. Experimental evidence that refuges delay insect adaptation to Bacillus thuringiensis. Proc. R. Soc. Lond. B: Biol. Sci., 264: 605-610.
CrossRefDirect Link - Liu, Y.B., B.E. Tabashnik and M. Pusztai-Carey, 1996. Field-evolved resistance to Bacillus thuringiensis toxin CryIC in diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol., 89: 798-804.
CrossRefDirect Link - Liu, Y.B., B.E. Tabashnik, T.J. Dennehy, A.L. Patin and A.C. Barlett, 1999. Development time and resistance to Bt crops. Nature, 400: 519-520.
CrossRefDirect Link - Losey, J.E., L.S. Rayor and M.E. Carter, 1999. Transgenic pollen harms monarch larvae. Nature, 399: 214-214.
CrossRefPubMedDirect Link - Macintosh, S.C., T.B. Stone, R.S. Jokerst and R.L. Fuchs, 1991. Binding of Bacillus thuringiensis proteins to a laboratory-selected line of Heliothis virescens. Proc. Natl. Acad. Sci. USA., 88: 8930-8933.
PubMedDirect Link - Mallet, J. and P. Porter, 1992. Preventing insect adaptation to insect-resistant crops: Are seed mixtures or refugia the best strategy? Proc. R. Soc. Lond. B: Biol. Sci., 250: 165-169.
CrossRefDirect Link - Manjula, K. and K. Padmavathamma, 1996. Effect of microbial insecticides on the control of Maruca testulalis and on the predators of redgram pest complex. Entomon, 21: 269-271.
Direct Link - McClintock, J.T., C.R. Schaffer and R.D. Sjoblad, 1995. A comparative review of the mammalian toxicity of Bacillus thuringiensis-based pesticides. Pestic. Sci., 45: 95-105.
CrossRefDirect Link - McGaughey, W.H., 1985. Insect resistance to the biological insecticide Bacillus thuringiensis. Science, 229: 193-195.
CrossRefPubMedDirect Link - McGaughey, W.H., 1994. Implications of cross‐resistance among Bacillus thuringiensis toxins in resistance management. J. Biocontrol Sci. Technol., 4: 427-435.
CrossRefDirect Link - McGaughey, W.H. and R.W. Beeman, 1988. Resistance to Bacillus thuringiensis in colonies of indianmeal moth and almond moth (Lepidoptera: Pyralidae). J. Econ. Entomol., 81: 28-33.
CrossRefDirect Link - McGaughey, W.H. and M.E. Whalon, 1992. Managing insect resistance to Bacillus thuringiensis toxins. Science, 258: 1451-1455.
CrossRefPubMedDirect Link - McGaughey, W.H. and D.E. Johnson, 1994. Influence of crystal protein composition of Bacillus thuringiensis strains on cross-resistance in indianmeal moths (Lepidoptera: Pyralidae). J. Econ. Entomol., 87: 535-540.
CrossRefDirect Link - Meade, T. and J.D. Hare, 1995. Integration of host plant resistance and Bacillus thuringiensis insecticides in the management of lepidopterous pests of celery. J. Econ. Entomol., 88: 1787-1794.
CrossRefDirect Link - Meadows, M.P., D.J. Ellis, J. Butt, P. Jarrett and H.D. Burges, 1992. Distribution, frequency and diversity of Bacillus thuringiensis in an animal feed mill. Applied Environ. Microbiol., 58: 1344-1350.
Direct Link - Metcalfe, R.L., 1989. Insect resistance to insecticides. Pestic. Sci., 26: 333-358.
CrossRefDirect Link - Milne, R. and H. Kaplan, 1993. Purification and characterization of a trypsin-like digestive enzyme from spruce budworm (Choristoneura fumiferana) responsible for the activation of δ-endotoxin from Bacillus thuringiensis. Insect Biochem. Mol. Biol., 23: 663-673.
CrossRefPubMedDirect Link - Milne, R.E., A.S.D. Pang and H. Kaplan, 1995. A protein complex from Choristoneura fumiferana gut-juice involved in the precipitation of δ-endotoxin from Bacillus thuringiensis subsp, sotto. Insect Biochem. Mol. Biol., 25: 1101-1114.
CrossRefDirect Link - Moar, W.J., M. Pusztai-Carey, H. van Faassen, D. Bosh and R. Frutos et al., 1995. Development of Bacillus thuringiensis CryIC resistance by Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae). Applied Environ. Microbiol., 61: 2086-2092.
Direct Link - Monnerat, R.G. and D. Bordat, 1998. Influence of HD1 (Bacillus thuringiensis ssp. kurstaki) on the developmental stages of Diadegma sp. (Hym., Ichneumonidae) parasitoid of Plutella xylostella (Lep., Yponomeutidae). J. Applied Entomol., 122: 49-51.
CrossRefDirect Link - Muller-Cohn, J., J. Chaufaux, C. Buisson, N. Gilois, V. Sanchis and D. Lereclus, 1996. Spodoptera littoralis (Lepidoptera: Noctuidae) resistance to CryIC and cross-resistance to other Bacillus thuringiensis crystal toxins. J. Econ. Entomol., 89: 791-797.
CrossRefDirect Link - Nealis, V. and K. van Frankenhuyzen, 1990. Interactions between Bacillus thuringiensis berliner and Apanteles fumiferanae vier. (Hymenoptera: Braconidae), a parasitoid of the spruce budworm, Choristoneura fumiferana (clem.) (Lepidoptera: Tortricidae). Can. Entomol., 122: 585-594.
CrossRefDirect Link - Mohan, R.F., E. Alcantara, M.K. Lee, X.J. Chen, A. Curtiss and D.H. Dean, 1995. Single amino acid changes in domain II of Bacillus thuringiensis CryIAb δ-endotoxin affect irreversible binding to Manduca sexta midgut membrane vesicles. J. Bacteriol., 177: 2276-2282.
Direct Link - Roush, R.T. and J.A. McKenzie, 1987. Ecological genetics of insecticide and acaricide resistance. Ann. Rev. Entomol., 32: 361-380.
CrossRefDirect Link - Salama, H.S., F.N. Zaki and M.M. Sabbour, 1996. Effect of Bacillus thuringiensis endotoxin on Apanteles litae Nixon andBracon instabilis Marsh (Hymenoptera: Braconidae), two parasitoids of the potato tuber moth Phthorimia operculella Zeller (Lepidoptera: Gelishiidae). J. Applied Entomol., 120: 565-568.
- Sayyed, A.H., R. Haward, S. Herrero, J. Ferre and D.J. Wright, 2000. Genetic and Biochemical approach for characterization of resistance to Bacillus thuringiensis toxin Cry1Ac in a field population of the Diamondback moth. Applied Environ. Microbiol., 66: 1509-1516.
Direct Link - Sayyed, A.H., J. Ferre and D.J. Wright, 2000. Mode of inheritance and stability of resistance to Bacillus thuringiensis to Plutella xylostella populations from Malysia. Pest. Manage. Sci., 56: 743-748.
Direct Link - Sayyed A.H. and D.J. Wright, 2001. Cross-resistance and inheritance of resistance to Cry1Ac toxin in diamondback moth, Plutella xylostella from lowland Malaysia. Pest. Manage. Sci., 57: 413-421.
PubMed - Sayyed, A.H. and D.J. Wright, 2001. Fitness costs and stability of resistance to Bacillus thuringiensis in a field population of the diamondback moth Plutella xylostella L. Ecol. Entomol., 26: 502-508.
Direct Link - Tabashnik, B.E., 1994. Evolution of resistance to Bacillus thuringiensis. Ann. Rev. Entomol., 39: 47-79.
CrossRefDirect Link - Andow, D.A., D.M. Olson, R.L. Hellmich, D.N. Alstad and W.D. Hutchison, 2000. Frequency of resistance to Bacillus thuringiensis toxin Cry1Ab in an Iowa population of European corn borer (Lepidoptera: Cramidae). J. Econ. Entomol., 93: 26-30.
CrossRefPubMedDirect Link - Talekar, N.S. and J.C. Yang, 1991. Characteristic of parasitism of diamondback moth by two larval parasites. Biocontrol, 36: 95-104.
CrossRefDirect Link - Van Frankenhuyzen, K., C.W. Nystom and B.E. Tabashnik, 1995. Variation in tolerance to Bacillus thuringiensis among and within populations of the Spruce budworm (Lepidoptera: Tortricidae) in Ontario. J. Econ. Entomol., 88: 97-105.
PubMedDirect Link - Visser, B., D. Bosch and G. Honee, 1993. Domain Function Studies of Bacillus thuringiensis Crystal Proteins a Genetic Approach. In: Bacillus thuringiensis and Environmental Biopesticide theory and Practice, Entwistle, P.F., J.S. Cory, M.J. Bailey and S. Higgs (Eds.). John Wiley and Sons, Chichester, UK.
- Wirth, M.C., A. Delecluse, B.A. Federici and W.E. Walton, 1998. Variable cross-resistance to Cry11B from Bacillus thuringiensis subsp. Jegathesan in Culex quinquefasciatus (Diptera:Culicidae) resistant to single or multiple toxins of Bacillus thuringiensis ssp. Israelensis. Applied Environ. Microbiol., 64: 4174-4179.
- Wright, D.J., M. Iqbal, F. Granero and J. Ferre, 1997. A change in a single midgut receptor in the diamondback moth Plutella xylostella is only in part responsible for the field resistance to Bacillus thuringiensis subsp Kurstaki and B. thuringiensisssp. aizawai. Applied Environ. Microbiol., 63: 1814-1819.
- Tabashnik, B.E., A.L. Patin, T.J. Dennehy, Y.B. Liu, Y. Carriere, M.A. Sims and L. Antilla, 2000. Frequency of resistance to Bacillus thuringiensis in field populations of pink bollworm. Proc. Nat. Acad. Sci., 97: 12980-12984.
Direct Link - Liu, Y. and B.E. Tabashnik, 1997. Inheritance of resistance to the Bacillus thuringiensis toxin Cry1C in the diamondback moth. Applied Environ. Microbiol., 63: 2218-2223.
Direct Link - Heimpel, A.M. and T.A. Angus, 1960. Bacterial insecticides. Bacteriol. Rev., 24: 266-288.
Direct Link