
Abdul Gafur
Department of Plant Protection, University of Lampung, Bandar Lampung 35145, Indonesia
Subli Mujim
Department of Plant Protection, University of Lampung, Bandar Lampung 35145, Indonesia
Titik Nur Aeny
Department of Plant Protection, University of Lampung, Bandar Lampung 35145, Indonesia
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 11 | Page No.: 1195-1198







ABSTRACT
The present study was aimed at elucidating the morphological variations of Indonesian isolates of Cochliobolus heterostrophus (Drechsler) Drechsler as well as exploring pathogenicity of the isolates. Potential specificity between host varieties and pathogen isolates in the corn–southern leaf blight pathosystem was also exploited. Isolates of C. heterostrophus were collected from ecologically different parts of the country including the islands of Bali, Java, Sulawesi and Sumatera. Variations in morphological and pathogenic characteristics were observed among the isolates although possible host specificity was unlikely to exist between the corn varieties and fungal isolates tested. These results suggested that agricultural ecosystems might have affected morphological and pathological variations within Indonesian isolates of the fungus provided the existence of a common ancestor of the isolates.
PDF Abstract XML References Citation
How to cite this article
Abdul Gafur, Subli Mujim and Titik Nur Aeny, 2002. Morphological and Pathological Variations in the Indonesian Cochliobolus heterostrophus (Pleosporaceae, Pleosporales, Euascomycetes). Pakistan Journal of Biological Sciences, 5: 1195-1198.
DOI: 10.3923/pjbs.2002.1195.1198
URL: https://scialert.net/abstract/?doi=pjbs.2002.1195.1198
DOI: 10.3923/pjbs.2002.1195.1198
URL: https://scialert.net/abstract/?doi=pjbs.2002.1195.1198
INTRODUCTION
Cochliobolus heterostrophus (Drechsler) Drechsler [anamorph: Bipolaris maydis (Nisikado and Miyake) Shoemaker] (Pleosporaceae, Pleosporales) is a pathogen of different diseases on a wide variety of gramineous plants but is the best known as causal agent of southern corn leaf blight. Pathogenic races of the fungus cause southern corn leaf blight disease, an economically important disease in the history of plant pathology. In 1970 alone an estimated one billion US$ loss due to the sudden epidemic of race T of the fungus was reported in USA. Widespread epidemic of the fungus also occurred in other parts of the world including Indonesia.
Because of its economic importance, C. heterostrophus has received considerable interests from researchers around the world. To some extent, it has been used as a model eukaryotic plant pathogen for analyzing mechanisms of pathogenicity (Leach et al., 1982) due to its advantage of being relatively easy to manipulate and therefore preferable for the study of fundamental biology. The fungus has also been employed in a number of scientific investigations including host-specific toxin production (Bronson et al., 1990), sexual reproduction and sexuality (Turgeon et al., 1993; Turgeon et al., 1996), pigmentation (Tanaka et al., 1991; Tanaka et al., 1995; Shimizu et al., 1997), fungicide resistance (Gafur et al., 1998a; Gafur et al., 1998b; Gafur et al., 2001) and mating type determination (Gafur et al., 1997).
Although a number of different investigations on Indonesian isolates of C. heterostrophus have been reported, only a little was known about intra specific morphological and pathological variations within the fungus. Research on the variations of the fungus had scarcely been conducted in Indonesia and only limited information on its morphological and pathological diversities is available. The current scientific understanding of the pathogen could threaten the production of corn, one of the most important food crops in Indonesia. This could be devastating and may contribute to an unexpectedly widespread epidemic in future.
To avoid this drawback, morphological and pathological variations of the species are needed to be clarified. Exploration on the subject was, therefore, a research priority not only for the advancement of science and technology, but also for practical aspects. The current research was aimed at investigating the morphological and pathological variations within the Indonesian isolates of C. heterostrophus, which is crucial to establish the fundamental basis for more comprehensive studies on the fungus. The ultimate goal of this initiative was a better understanding of the host - pathogen interaction in the pathosystem leading to the development of effective disease control strategies. As a number of different corn varieties have been cultivated in Indonesia under different cropping systems and agroecological conditions for an expanded period of time, it is hypothesised that morphological and pathological diversities occur within Indonesian isolates of C. heterostrophus. The present investigation was an effort to challenge that hypothesis.
MATERIALS AND METHODS
Biological materials and media: The research was carried out at College of Agriculture, University of Lampung, Bandar Lampung, Indonesia. The strains used in this study were different isolates of C. heterostrophus collected from ecologically different corn areas in Indonesia (Table 1). HITO7711 and MASHIKI2-2 strains, wild type stock cultures whose mating types had been determined (MAT1-2 and MAT1-1, respectively) (Tanaka et al., 1991), were used as tester strains in crossing experiments (Gafur et al., 1998b). The experiments also involved two different corn varieties, Bisma and C-7. Media used included Sachs’ agar medium for crossing experiments and complete medium for morphological observations and culture maintenance. Sachs’ agar medium contained (g l–1) KNO3, 1.0; MgSO4. 7H2O, 0.5; NaCl, 0.5; Ca(NO3)2, 0.5; Ca3(PO4)2, 0.5; FeCl3, trace and agar, 12.0. The medium was autoclaved at 121°C for 15 min. Complete medium was prepared with distilled water to contain the following components (g l–1): Ca(NO3)2.4H2O, 1.5; MgSO4.7H2O, 0.5; KCl, 0.5; KH2PO4, 0.4; K2HPO4, 0.03; glucose, 10.0; tryptone, 1.0; yeast extract, 1.0 and agar, 15.0. The medium was also autoclaved at 121°C for 15 min.
| Table 1: | Isolates of Colchliobolus heterostrophus used in this research and their origins |
 | |
Crossing experiment: Crossing experiments between each of the isolates and the two sexually known tester strains were conducted to produce asci for morphological observations. Crossing was performed according to the method of Ueyama and Tsuda (1975). Small pieces of mycelia of strains to be crossed were inoculated at the opposite sides of a piece of sterilised rice straw that had previously been put on Sachs' agar medium. The inoculated dishes were incubated at 24°C under intermittent fluorescent light. Mature pseudothecia were harvested after three weeks of incubation.
Morphological observation: Morphological variations in C. heterostrophus collected from different places of Indonesia were observed both macroscopically and microscopically. This mainly included observations on colony colour, conidial size, conidial septa and ascal size. All the fungal isolates used for conidial investigations were grown in petri dishes containing complete medium. Stock cultures of the isolates were maintained on the same medium in slant tubes. Asci were produced after crossing compatible isolates on Sachs’ agar medium as explained above.
Pathogenicity test: Corn plants were exposed to conidial suspensions of each of different isolates of C. heterostrophus. The inoculated plants were covered with plastic bags to prevent fungal conidia from aerial dispersion that may contaminate the neighbouring plants. Disease symptoms, mainly localized on leaf lesions developed following inoculation, were observed on a daily basis. Disease incidence (percent of diseased leaves to the total leaves) and disease severity (by including disease development scores as one parameter) were recorded one, two and three weeks after inoculation. Controlled plants were inoculated with sterile distilled water.
Statistical analysis: All statistical analysis were performed using SAS for Windows. Data sets were analyzed using analysis of variance, ANOVA. Detected differences among the means were further analyzed using multiple comparisons of the Duncan’s multiple range test at a confidence level of P≤ 0.05.
RESULTS AND DISCUSSION
Crossing experiment: Formation of asci inside mature pseudothecia in crosses with any of the tester strains should also indicate that the isolates being crossed belong to the same species. In this study, formation of the fruiting body and sexual spore was evident three weeks after compatible crossings (Fig. 1). Crossing ability has in the past been used as one of the major taxonomic criteria in graminecolous fungi.
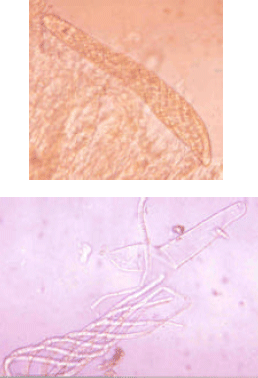 | |
| Fig. 1: | A banana-shaped ascus emerging from pseudothecia (left) and ascospores (right) released from an ascus were observed three weeks after compatible matings |
 | |
| Fig. 2: | Indonesian Cochliobolus heterostrophus collected in the present research on complete medium. The isolates vary considerably not only in colony colour, but also in their growth rate |
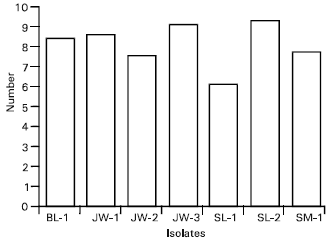 | |
| Fig. 3: | Average number of septa of the Indonesian Cochliobolus heterostrophus. Each value is the mean of 50 counts. |
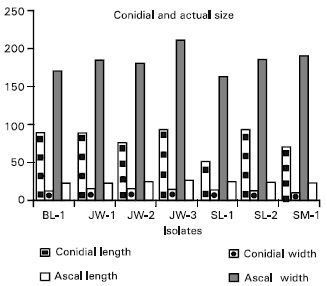 | |
| Fig. 4: | Variations of size of conidia and asci of different isolates of Indonesian Cochliobolus heterostrophus. Each value is the mean of 50 measurements |
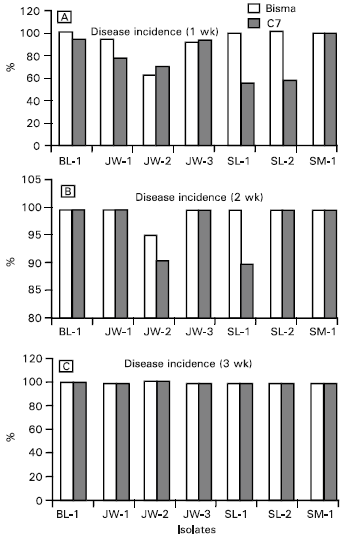 | |
| Fig. 5: | Incidence of the southern corn leaf blight disease caused by Indonesian isolates of Cochliobolus heterostrophus on corn varieties of Bisma and C-7 one (a), two (b) and three (c) weeks after inoculation |
This is particularly true since the middle of 1970’s when researchers were for the first time able to produce sexual stages of the fungi in laboratory (Ueyama and Tsuda, 1975) using methods similar to the one employed in the current research. It was an important contribution to the genetic study of C. heterostrophus, whose perfect stage had rarely been found in the field.
Morphological observation: Phenotypic variations of the Indonesian C. heterostrophus collected in the present research were obvious (Fig. 2). Colony colour of the isolates was slightly different, ranging from dark green to greyish with whitish surface. The growth rate of the fungus on complete medium also varied among isolates. These characteristics are not uncommon to mycologists and have therefore been a subject of research interests for some time. Biochemical pathway of the pigmentation in the fungus was already clarified and a number of genes controlling the trait have also been isolated and characterized (Tanaka et al., 1991; Tanaka et al., 1995; Shimizu et al., 1997). Variations were also detected in the average number of conidial septa (Fig. 3) and sizes of conidia and asci (Fig. 4) among isolates. The average number of septa per conidia ranged from as low as six in SL-1 isolate to more than nine in JW-3 and SL-2, whereas the length of conidia varied from 51,38 to 97,13 μm. Conidia with higher number of septa were generally considered bigger in size. Less variation was observed in the width of conidia. Similarly, variations within the width of asci were less than that of their length. In fact, non significant differences were statistically detected on the width of asci.
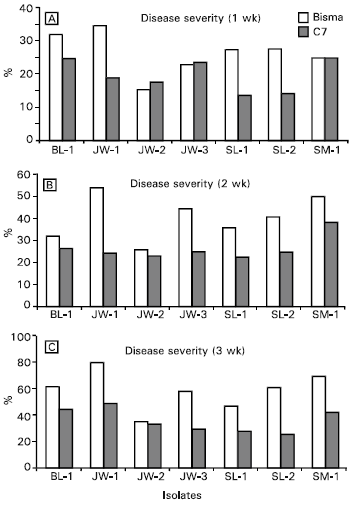 | |
| Fig. 6: | Severity of the southern corn leaf blight disease caused by Indonesian isolates of Cochliobolus heterostrophus on corn varieties of Bisma and C-7 one (a), two (b) and three (c) weeks after inoculation |
Although depending very much on circumstantial conditions, conidial morphology was in the past also used as a principal method for intra specific separation in C. heterostrophus. Many races have been subsequently proposed based on conidial morphology (Alcorn, 1983; Sivanesan, 1987).
Pathogenicity test: Pathogenicity of the isolates was measured in terms of disease incidence (Fig. 5) and disease severity (Fig. 6). Variations in pathogenicity of the Indonesian C. heterostrophus on both corn varieties used, especially within the first week of inoculation, were statistically significant although variations in disease incidence is less than that in disease severity. In fact, three weeks after inoculation, disease symptoms were observed in all leaves of inoculated plants of both varieties. Similarly, variations in disease severity tended to decrease with time on both corn varieties.
Isolate SM-1 appeared to be the most virulent isolate on both varieties, whereas variety Bisma was more resistant to all fungal isolates than variety C-7. No clear-cut relationship could be confirmed between variations in pathogen virulence and host resistance that might lead to possible host specificity. Provided that all the isolates tested have a common ancestor, it could be implied that agroecological conditions might have been contributing to the morphological and pathological variations within the fungus.
As already mentioned, variations in morphological and pathogenicity characteristics of the Indonesian isolates of C. heterostrophus are detected in the present study although host specificity is unlikely to exist between the corn varieties and fungal isolates presently tested. These results, however, should provide a strong basis for investigations on molecular and biochemical host–pathogen interactions in the corn–southern leaf blight pathosystem which certainly are required for developing more effective disease control strategies. The results of the present study also imply that more works have yet to be attempted in order for us to have better understanding of this corn–southern leaf blight pathosystem.
ACKNOWLEDGMENTS
This research work was financially supported by the DCRG-URGE Project (Contract No. 008/DCRG/URGE/2000), DGHE, Ministry of National Education of the Republic of Indonesia, Jakarta, Indonesia. We wish to thank R. Hakim, D. Maharani, A. Oktavia (green house experiments), R. Rinda and W. Widyaningrum (laboratory experiments) for excellent technical assistance.
REFERENCES
- Bronson, C.R., M. Taga and O.C. Yoder, 1990. Genetic control and distorted segregation of the T-toxin production in field isolates of Cochliobolus heterostrophus. Phytopathology, 80: 819-823.
Direct Link - Gafur, A., C. Tanaka, K. Shimizu, S. Ouchi and M. Tsuda, 1998. Molecular analysis and characterization of the Cochliobolus heterostrophus β-tubulin gene and its possible role in conferring resistance to benomyl. J. Gen. Applied Microbiol., 44: 217-223.
CrossRef - Gafur, A., C. Tanaka, K. Shimizu, S. Ouchi and M. Tsuda, 1998. Genetic analysis of Cochliobolus heterostrophus polyoxin-resistant mutants. Mycoscience, 39: 155-159.
CrossRef - Leach, J., B.R. Lang and O.C. Yoder, 1982. Methods for selection of mutants and in vitro culture of Cochliobolus heterostrophus. J. Gen. Microbiol., 128: 1719-1729.
CrossRef - Shimizu, K., C. Tanaka and M. Tsuda, 1997. Cloning of Brn1, a reductase gene involved in melanin biosynthesis in Cochliobolus heterostrophus. J. Gen. Applied Microbiol., 43: 145-150.
PubMed - Tanaka, C., Y. Kubo and M. Tsuda, 1991. Genetic analysis and characterization of Cochliobolus heterostrophus colour mutants. Mycol. Res., 92: 49-56.
Direct Link - Tanaka, C., T. Taniguchi and M. Tsuda, 1995. Genetic analysis of Cochliobolus heterostrophus beige mutants. Mycol. Res., 99: 621-622.
Direct Link - Turgeon, B.G., H. Bohlmann, L.M. Ciuffetti, S.K. Christiansen, G. Yang, Y. Schafer and O.C. Yoder, 1993. Cloning and analysis of the mating-type genes from Cochliobolus heterostrophus. Mol. Gen. Genet., 238: 270-284.
PubMed - Ueyama, A. and M. Tsuda, 1975. Formation in culture of Cochliobolus miyabeanus, the perfect state of Helminthosporium oryzae. Ann. Phytopathol. Soc. Jap., 41: 434-440.
Direct Link