
A. T. Elsa
Department of Pharmacology, Usmanu Danfodiyo University, Sokoto, Nigeria
U. A. Osunkwo
Department of Veterinary Medicine, Surgery and Theriogenology,
P. A. Onyeyili
Department of Veterinary Medicine, Surgery and Theriogenology,
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 11 | Page No.: 1160-1161







ABSTRACT
The changes in electrical activity of the cerebral cortex during amylobarbitone anaesthesia were studied in rabbits. Electrical activity of the rabbit cerebral cortex recorded (using an oscilloscope), sixty minutes after induction of amylobarbitone (30 mg kg–1, body weight, I.P.) anaesthesia did not significantly differ from control values obtained before anaesthesia or 24 h after recovery from anaesthesia. Amylobarbitone administered @ 60, 90 and 120 mg kg–1 body weight significantly (P<0.05) and dose - dependently decreased the cerebral electrical activity. Therefore, amylobarbitone at the dose of 60 mg kg–1 body weight or higher may cause depression in the electrical activity of the rabbit brain.
PDF Abstract XML References Citation
How to cite this article
A. T. Elsa, U. A. Osunkwo and P. A. Onyeyili, 2002. Changes in Electrical Activity of the Cerebral Cortex During Amylobarbitone Anaesthesia in Rabbits. Pakistan Journal of Biological Sciences, 5: 1160-1161.
DOI: 10.3923/pjbs.2002.1160.1161
URL: https://scialert.net/abstract/?doi=pjbs.2002.1160.1161
DOI: 10.3923/pjbs.2002.1160.1161
URL: https://scialert.net/abstract/?doi=pjbs.2002.1160.1161
INTRODUCTION
The single most important discovery in the exploration of nervous mechanism was that the nerve impulse is identical with an electrical change (Delmonte, 1984; Abbott and Howarth, 1973). This electrical sign of activity has given the investigator a means of studying the function of nervous system in the living organisms through the complexity of structure that the microscope can reveal only in dead tissue (Gasanov, 1987).
Recording instruments have evolved through the capillary electrometer, the Einthoven string galvanometer and electromagnetic recorders to the cathode–ray oscilloscope. The cathode ray oscilloscope device with its inertialess electron beam has proved to be the instrument of choice for recording the exceedingly brief and feeble currents of nerve activity and has become the display unit for modern laboratory computers (Coombs et al., 1955; Mize, 1984).
The measurement of nerve conduction velocity is now a standard electrodiagnostic procedure in clinical neurology (De Jesus et al., 1973). The most commonly used techniques estimate the conduction rates of fast fibers only (Dawson, 1956), but new methods allow for the study of slow conducting sensory (Butchthal and Rosentalek, 1971) and motor fibers (Butchthal and Rosentalek, 1971; Thomas et al., 1985). Several investigations have been performed using motor evoked potential monitoring as a diagnostic tool for detecting the impending damage of central nervous system descending pathways in an early still reversible state during anesthesia and neurosurgery (Thees et al., 1997). In many fields of neurophysiology it is often of interest to observe the firing pattern of single neuron on an oscilloscope. It is convenient at the same time to observe the rate of cell firing and to record it on a chart recorder (Graystone et al., 1969; Gasanov, 1987). General anaesthetic gains access to the brain rapidly inducing a stable level of anaesthesia and providing an accurate index of potency for every class of anaesthetic agents (Firestone et al., 1986). Amylobarbitone is an intermediate acting barbiturate widely used in laboratory animals for organ resection purposes (Goetz et al., 1987; Osunkwo et al., 1989). It was therefore of interest to develop a technique to record acutely the discharges in the cerebral cortex in the amylobarbitone anaesthetized rabbits.
MATERIALS AND METHODS
Twenty New Zealand rabbits of either sex weighing 1.0–2.3 kg and aged 6 to 12 months were used for the study. The rabbits were divided into four groups each of five and housed in research laboratory, Department of Pharmacology, Usmanu Danfodiyo University Sokoto. The rabbits were fed with green leaves and rabbits pellet. Water was provided ad libitum. All the rabbits were groomed with coumaphous (Asuntol 16%, Bayer Pharmaceuticals, Switzerland) and dewormed with Levamisole (Citarin-L, 2.5% solution, Bayer Pharmaceuticals. Switzerland) at a subcutaneous dose of 5 mg kg–1 body weight. The animals were kept for observation for four weeks period of acclimatization before the experiment was commenced.
Rabbits in group A, received amylobarbitone (30 mg kg–1, body weight, I.P.) and the electrical activity of the cerebral cortex was measured after 60 min of anaesthesia induction by placing the oscilloscope (Hitachi Denshi Ltd., Tokyo, Japan) probe on the cerebral cortex through a hole, drilled in the skull with spinal needle. The electrical signals of the cortex were fed into the digital recorder (Model WR7700, Graphtec Corporation, Japan) and recorded on the chart paper.
Animals in groups B, C and D were treated with amylobarbitone intraperitoneally at a dose of 60, 90 and 120 mg kg–1, body weight respectively. A dorsal craniotomy was performed on all the rabbits in three groups. The head was clipped, shaved and disinfected. A craniocaudal midline skin incision was made on the skull. The scalp was cut along the midline and retracted with the temporal muscle in order to reveal nearly all the frontal and parietal bones and as much of the temporal bone as possible. All the subcutaneous tissues were carefully scrapped away. The skull was cut with a scalpel blade along with the tissues attaching parietal, maxillary and frontal and occipital bones. The part of the cut skull was reflected cranially to expose the cerebral cortex. The probe of the oscilloscope was placed on the cortex in situ. The signals of the electrical activity of the cortex was fed into the digital recorder and recorded on the chart paper. The reflected bone of the skull was placed into its anatomical position. The skin was sutured with silk in simple interrupted suture pattern. The surgical site was dressed with gentian violet. Postoperatively, 400,000 IU. procaine penicillin was injected intramuscularly once daily for three days. On postoperative day one, the interrupted skin sutures were removed and the signals of the electrical activity of the cortex were recorded. The reflected part of the skull and skin was returned and re-sutured. Gentian violet was applied on the surgical site to prevent flies and other insects and to enhance the healing process. Data obtained from the study was presented as mean±standard deviation and differences between means were analyzed using analysis of variance (ANOVA).
RESULTS
The results of the oscilloscope recordings of the electrical activity of the cerebral cortex at four different doses of amylobarbittone (30, 60, 90 and 120 mg kg–1) indicated that the electrical activity of the rabbits cerebral cortex was recorded 60 min after induction of anaesthesia (Table 1) and 30 mg kg–1 amylobarbitone differed non-significantly from the control values recorded before anaesthesia or values obtained 24 h after recovery from anaesthesia.
| Table 1: | Electrical activity of the cerebral cortex during treatment with amylobarbitone |
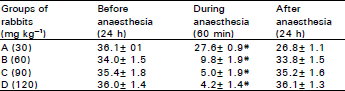 | |
| * P<0.05 when compared with values before anaesthesia and 60 min. during anaesthesia using analysis of variance. | |
Although amylobarbitone at 60 mg kg–1 body weight, significantly (P<0.05) decreased cerebral cortex electrical activity after 60 min of anaesthesia, induction of higher doses of amylobarbitone (90 or 120 mg kg–1 body weight) significantly (P<0.05) and dose dependently decreased the cerebral electrical activity of rabbit brain.
DISCUSSION
The oscilloscope was used to study the electrical activity of the rabbit brain because it has proved to be the instrument of choice for recording the exceeding brief and feeble currents of nerves and has become the display unit for modern laboratory works (Graystone et al., 1969; Evans, 1985). When amylobarbitone was administered at a lower dose, all the rabbits so treated showed insomnia with non significant decrease in electrical activity of the cerebral cortex, 60 min after induction of anaesthesia. The slight decrease in the electrical activity of cortex suggests that the central effect of amylobarbitone did not largely affect the cortex to produce significant changes in the rabbit brain.
However, amylobarbitone when administered at a higher dose, caused a significant decrease in electrical activity of the cerebral cortex of rabbits. Also much higher doses of amylobarbitone, significantly and dose-dependently further reduced the cerebral electrical activity. These observations can be explained by the fact that changes in electrical activity of the cortex at the normal metabolic state could be affected by the degree of depolarization of the nerves produced by the action of anaesthetic agent. As reported by Delmonte (1984) and Chen and Towe (1984), if the nerve is impaired by a narcotic agent its excitability will be reduced. It therefore follows that amylobarbitone may produce central effects consistent with an inhibition of these brain areas causing narcosis and subsequent decrease in electrical activity of cortex. Thus a possible cortico-inhibitory mechanism of central origin might be operating during amylobarbitone anaesthesia.
Another possible explanation for the mechanism of amylobarbitone produced reduction in the electrical activity of the cerebral cortex, is that many nerve cells are affected by the anaesthetic agent that could be responsible for adding their activity in such a way as to produce waves of amplitude having their major electrical characteristics in common (Thomas et al., 1985; Gasanov, 1987). The exact neuronal mechanisms at the membrane level are still to be defined to suit all the available data but there is now general agreement that neocortex potentials are generated by postsynaptic activity and that their rhythms are paced from the thalamus (Brazier, 1961; Nwankwoala and Ezeamuzie, 1995). However measurement of the electrical activity of the cerebral cortex of rabbits using an oscilloscope at 24 h before and after anaesthesia did not produce any significant change. This is probably due to the unavailability of anaesthetic agent in the body system. It is also possible that at least parts of the inhibitory actions at amylobarbitone on the cerebral cortex could be explained by cortical mechanism (Steriade and Linas, 1988). For example the electrical activity of the cortex was reduced by amylobarbitone at both lower and higher doses but remains remarkably unchanged in unanesthetized rabbit. These findings suggest that dose of amylobarbitone equal to or greater than 60 mg kg–1 body weight was adequate to cause decrease in the electrical activity of the rabbits’ brain.
ACKNOWLEDGMENTS
The authors gratefully acknowledge Usmanu Danfodiyo University Sokoto for partial funding of this work.
REFERENCES
- Abbott, B.C. and J.V. Howarth, 1973. Heat studies in excitable tissues. Physiol. Rev., 53: 120-158.
Direct Link - Butchthal, F. and A. Rosentalek, 1971. Sensory condition from digit to palm and palm to wrist in the carpal tunnel syndrome. J. Nurol. Neurosurg. Psychiatry, 34: 243-252.
CrossRef - Chen, Z. and A.L. Towe, 1984. Cerebral response to medullary pyramids stimulation in the rabbits. Brain Behav. Evol., 25: 175-186.
CrossRef - Delmonte, M.M., 1984. Electro cortical activity and related phenomena associated with meditation practice: A literature review. Int. Neurosci., 24: 217-231.
Direct Link - De Jesus, J.P.V., J.H. Petrusewicz and R.L. Barchi, 1973. The effect of cold on nerve conduction of human slow and fast nerve fibers. Neurology, 23: 1182-1189.
Direct Link - Gasanov, U.G., 1987. Amplitudinal principle for the structural functional classification of cortical neurons. Neurosci. Behav. Physiol., 17: 185-192.
CrossRef - Graystone, P., N. Owen and H. Mclennan, 1969. A simple rate meter and oscilloscope intensity modulator for single cell recording. Can. J. Physiol. Pharmacol., 47: 398-400.
PubMed - Steriade, M. and R.R. Linas, 1988. The functional states of the thalamus and the associated neuronal interplay. Physiol. Rev., 68: 649-742.
Direct Link