
Rubina Arshad
Nuclear Institute for Agriculture and Biology (NIAB), P.O. Box 128, Jhang Road, Faisalabad, Pakistan
Muhammad Salih Ahmad
Nuclear Institute for Agriculture and Biology (NIAB), P.O. Box 128, Jhang Road, Fap
Isalabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 5 | Page No.: 862-865







ABSTRACT
Penicillin G acylase (PGA) is one of the most widely used enzymes at an industrial scale. It is used to hydrolyze penicillin G to produce 6-aminopenicillanic acid (6-APA), a required intermediate in the production of semi-synthetic penicillins. Escherichia coli is the most abundantly used organism for PGA production. Present study was focused on the kinetics of growth and PGA production of three local E. coli isolates, FH6-NIAB, FMu2-NIAB and FD25-NIAB. Optimal enzyme production was reached after 18 hr. For longer incubations, the enzymatic activity decreased slightly. PGA activity was detectable after about 6 hours of incubation.
For the selected isolates, PGA production was determined for various concentrations of glucose and phenylacetic acid in the medium. PGA production by E. coli isolates was completely repressed by glucose concentration above 5 g/l. Production of PGA by E. coli isolates was completely dependent upon the presence of PAA in the medium and optimal enzyme activity was obtained between 1 g/l and 2 g/l of PAA. Enzyme activity was found to be highest when PAA was used as an inducer. PGA production by our local E. coli isolates was completely inducible.
PDF Abstract XML References Citation
How to cite this article
Rubina Arshad and Muhammad Salih Ahmad, 2000. Kinetic Study of Growth and Penicillin G Acylase Production by Escherichia coli. Pakistan Journal of Biological Sciences, 3: 862-865.
DOI: 10.3923/pjbs.2000.862.865
URL: https://scialert.net/abstract/?doi=pjbs.2000.862.865
DOI: 10.3923/pjbs.2000.862.865
URL: https://scialert.net/abstract/?doi=pjbs.2000.862.865
INTRODUCTION
Penicillin acylase (PA) is an enzyme of considerable commercial and medical interest. It is used in the production of 6-aminopenicillanic acid (6-APA), the nucleus used for industrial semi-synthesis of novel antibiotics. The crystal structure of PA from E. coli has been determined (McVey et al., 1997). Although the biological function of the enzyme remains something of a mystery, the 3D structure has revealed much to understand this enzyme.
It is estimated that 88 percent of the 6-APA obtained enzymatically results from penicillin G acylase activity; the rest results from penicillin V acylase activity (Vandamme, 1988). This is mainly because of the development of hyper-producing strains for PGA. PGA enzyme from E. coli ATCC 11105, Bacillus megaterium ATCC 14945, and from mutants of these two strains are the most commonly used in industry. Production of PGA by E. coli ATCC 11105 is repressed by glucose, induced by phenylacetic acid and abolished by growth at temperatures higher than 30°C (Wojskowicz, 1981).
As first noted by Kaufmann et al. (1960), a most important constituent of the medium is phenylacetic acid, which is metabolized during the course of bacterial growth and fermentation to induce enzyme synthesis, increasing its production by 5-10 fold. Vojtisek and Slezak (1975b) showed absolute enzyme repression by glucose to be independent of glucose concentration within the range of 0.1-1.0 percent.
Klein and Wagner (1980) found glucose, fructose and lactate to strongly repress acylase production with the industrial strain of E. coli ATCC 11105. With the constituent hybrid strain, however, these carbon sources increased enzyme levels. The effect of glucose on phenylacetic acid was further studied by Goluh and Bei'Kind (1977), who found that glucose repressed synthesis of components of PAA transport system and inhibited their activity. Thus E. coli growing on glucose did not take up PAA and hence did not synthesize acylase.
Robas et al. (1993) have defined the conditions for optimal expression of the PGA genes in recombinant E. coli strains; an initial glucose concentration of 5 g/l is sufficient to get optimal expression, however, PAA concentrations above 2 g/l have decreased stimulatory effect on any E. coli strain tested. The most important constituent of fermentation medium is PAA, which is metabolized during the course of bacterial growth, increasing enzyme production by 5-10 fold. Enzyme production is greatest at incubation temperature 24 and 30°C (Savidge and Cole, 1975).
The present study was conducted to evaluate the production of penicillin G acylase from E. coli isolates, to compare the effect of glucose and phenylacetic acid (PAA) on enzyme production and the kinetics of induction by PAA.
MATERIALS AND METHODS
E. coli strains and culture conditions: The microorganisms used in this study were Escherichia coli. E. coli FH6-NIAB, FMu2-NIAB and FD25-NIAB were isolated in our laboratory at NIAB from faeces of horse, mule and donkey, respectively. These particular strains were maintained on nutrient agar (Difco) slopes incubated at 37°C for 18 hr and stored at 5°C.
The inoculum was prepared in corn-steep liquor (CSL) medium. CSL 10 g/l (pH 7.0) supplemented or not with phenylacetic acid and/or glucose; 20 ml of CSL medium was dispensed in 250 ml Erlenmeyer flasks. Medium was sterilized at 121°C for 15 minutes. Inoculated broth was placed on a rotary shaker and incubated overnight at 26°C. Then, 5 ml of the cell suspension was used to inoculate 50 ml culture medium in 300 ml flasks. Culture medium contained: Bacto-peptone (Difco) 20 g/l, NaCl 5 g/l supplemented or not with PAA and/or glucose. Incubation was carried out on a rotary shaker at 26°C for 18 hr.
Glucose and phenylacetic acid concentrations: To study the influence of glucose concentration in the medium on PGA production, the same concentration of glucose was added to both the inoculum and the culture medium. The initial glucose concentrations varied between 0 to 50 g/l. Assays were performed at 0, 5, 10, 15, 20, 25, 30, 35, 40, 45 and 50 g/l glucose, in the presence of 2 g/l PAA. Effect of PAA concentration on PGA production by E. coli was studied. Initial PAA concentration varied between 0 to 5 g/l. Assays were performed at 0, 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 4.0 and 5.0 g/l PAA in the presence of 5 g/l glucose.
Determination of Growth: After 18 hours of incubation, pH and relative cell growths of the various cultures were estimated by measuring the absorbance (O.D.) at 660 nm.
Determination of PGA production: Cells were harvested by centrifugation for 20 min at 4000 g. Cells were washed with 0.05 M phosphate buffer (pH 7.5) and re-suspended in the same buffer. PGA activity was determined by spectrophotometric p-dimethylaminobenzaldehyde method (Saba et al., 1998).
Protein estimation: Protein contents of E. coli FH6-NIAB, FMu2-NIAB and FD25-NIAB were estimated as described earlier (Ahmad et al., 1999).
RESULTS AND DISCUSSION
Effect of glucose: After overnight incubation, the glucose concentration above 5 g/l strongly inhibited the cell growth of these isolates studied. The pH of the medium at the end of culture was basic in the absence of glucose and at glucose concentration of 5 g/l. However, pH of the medium decreased (acidic) with the increase in glucose concentration from 10 g/l to 50 g/l, due to acetate production (Fig. 1a).
Initial glucose concentration above 5 g/l inhibits cell growth (Fig. 1b), protein contents (Fig. 1c) and PGA production (Fig. 1d). Better results in terms of PGA production and productivity were obtained in complex medium with glucose concentration below 5 g/l (Fig. 1d). Glucose apparently repressed PGA synthesis in E. coli. PGA production in E. coli isolates was completely repressed and decreased by glucose concentration above 5 g/l. In the presence of glucose, the observed decrease in PGA activity was not simply the result of the growth decrease. Protein contents also showed similar results. With the increase in glucose concentration above 5 g/l protein contents of isolates decreased (Fig. 1c). It can be concluded that glucose concentration above 5 g/l is not beneficial for PGA production in the culture medium as it delays the onset of synthesis, which might be attributed to catabolic repression. Illanes et al. (1994) have reported that when glucose was omitted from the complex medium, both penicillin acylase production and productivity from Bacillus megaterium were significantly higher, and only a slight decrease in specific growth rates was observed.
Effect of PAA: The effect of PAA concentration in the medium on E. coli isolates was tested in the presence of 5 g/l glucose. The pH of cultures grown decreased with the increase in PAA concentration (g/l) in the medium (Fig. 2a). In the absence of PAA, pH of culture was basic however, with PAA at 4 g/l, the pH of culture was acidic. Figure 2b indicates that at initial PAA concentration 1.5 g/l, cell density was more and at 2.0 g/l to 5.0 g/l, the cell number decreased. Increasing PAA concentration was found to increase the E. coli final cell number. For all isolates, optimal production was obtained for PAA concentration at 1.5 g/l. As shown in Fig. 2c, the value of 0.5 g/l PAA was a clear optimum in terms of protein contents of different E. coli cultures studied. At higher values they decreased considerably as was the case for cell growth rate and productivity.
PGA induction was studied in terms of the level of the addition of PAA to the culture medium. Results for the effect of PAA concentration are presented in Fig. 2d. When the effect of PAA was under study, rates were calculated on the basis of 6-APA production (mg 6-APA/hr/mg protein).Induction of 0.5 g/l to 2.0 g/l PAA stimulated PGA activity. An optimal enzyme production was obtained for a PAA concentration around 1.5 g/l to 2.0 g/l. Thus the level of PAA affected the synthesis and production of PGA, but the effect on cell growth and protein contents was not significant with some decrease at levels higher than 2.0 g/l. From the overall results, it is concluded that PAA and glucose have additive stimulatory effects on our local E. coli isolates and PGA production reached levels as high as 4.7 to 7.0 mg 6-APA/hr/mg protein (Fig. 2d).
Kinetics of growth and PGA production: The kinetics of growth and PGA production was also studied for E. coli isolates grown in a medium supplemented by 2.0 g/l PAA in the presence of 5 g/l glucose.
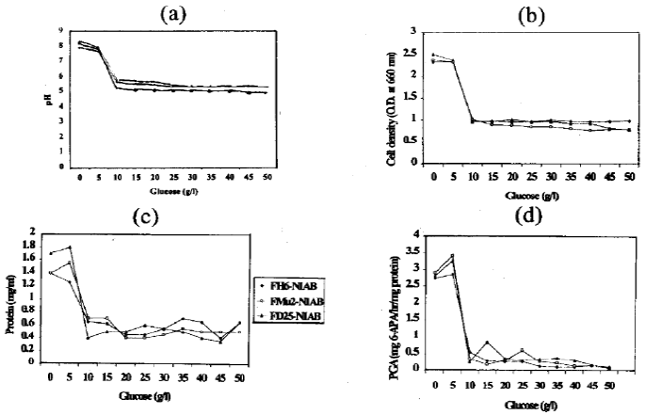 | |
| Fig. 1(a-d): | The effect of glucose concentration in the medium on PGA production by E. coli isolates. (a) pH of the medium at the end of the cultures, (b) cell densities of the cultures at 660 nm, (c) protein contents (mg/ml) of the cultures at 300 nm and (d) total PGA production of various cultures expressed in mg 6-APA/hr/mg protein |
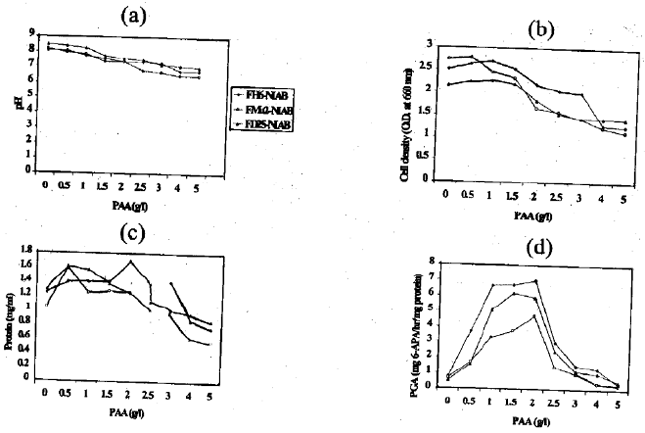 | |
| Fig. 2(a-d): | The effect of PAA concentration in the medium on PGA production by E. coli isolates tested in the presence of 5 g/l glucose concentration. (a) pH, (b) cell density at 660 nm, (c) protein contents at 300 nm and (d) total PGA production |
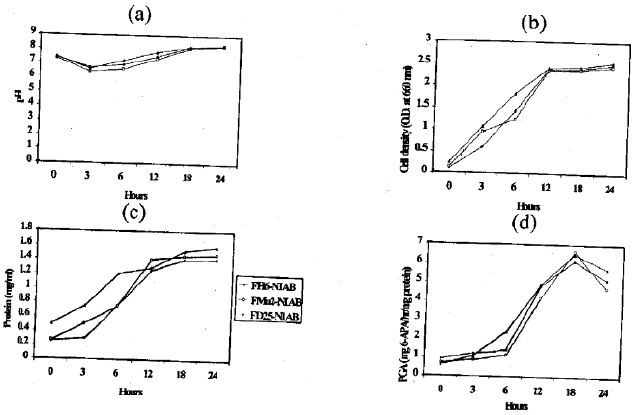 | |
| Fig. 3(a-d): | Kinetics of PGA production by E. coli isolates cultivated in a medium supplemented by 2 g/l PAA in the presence of 5 g/l glucose. (a) pH, (b) cell density at 660 nm, (c) protein contents at 300 nm and (d) total PGA production |
The growth rate, pH of culture, protein contents and final yield of PGA from our local E. coli isolates were compared at different time Intervals. The cells were grown for a maximum incubation period of 24 hr and were harvested after every three hour. Figure 3 shows that no marked difference of behaviour of the three isolates could be observed all along the cultures for any condition tested.
The pH of the medium at the start of cultures was basic. However, after 3 hr of incubation pH of the cultures decreased and was found to be slightly acidic (Fig. 3a). With the increase in incubation period, the pH of cultures showed an increasing trend and after 24 hr this pH was again basic. The rate of culture growth was determined by taking samples at various time intervals and measuring the density at 660 nm. Figure 3b shows that these E. coli isolates have similar initial growth rates and they showed a marked increase in cell growth shortly after 6 hours of incubation. Cells increased in number with increase in incubation period. Optimal cell growth was observed after 18 hours of incubation. Similarly, protein contents increased with longer periods of incubation (Fig. 3c).
The kinetics of PGA production in the medium supplemented by PAA (2.0 g/l) at a glucose concentration of 5 g/l is represented in Fig. 3d. The enzyme activity was detectable after 6 hours of incubation. Optimal enzyme production was reproducibly reached after 18 hr. For longer incubations, the enzymatic activity decreased slightly, probably because of proteolysis. Cell growth, protein contents and enzyme production kinetics were similar in all cases and the values reported in Fig. 3 represent the higher activities obtained at 18 hours of incubation.
Our results demonstrate that an initial glucose concentration of 5 g/l is sufficient to get optimal PGA production. The effect of glucose on PGA production decreases for glucose concentrations above 5 g/l. PGA was induced by PAA, the optimum conditions being 1.5-2.0 g/l when added at the onset of fermentation. PAA can be metabolized by E. coli and used as the sole carbon source, which makes induction a complex kinetic event. For any E. coli isolate tested however, PAA concentrations above 2.0 g/l have a decreased stimulatory effect. A similar observation, made for E. coli ATCC 11105, was previously attributed to a catabolic repression by PAA (Vojtisek and Slezak, 1975a; Wojskowicz, 1981).
The results of our study were compared with others reported in the literature. As in other cases, PGA induction by our E. coli isolates was competitively inhibited by PAA and glucose concentrations above 2.0 g/l and 5 g/l, respectively. This is encouraging for further studies on strain improvement by media formulation.
REFERENCES
- Ahmad, M.S., M. Saba and R. Arshad, 1999. Escherichia coli producing penicillin g acylase from dung of horse, donkey and mule. Pak. J. Zool., 31: 141-145.
Direct Link - Illanes, A., F. Acevedo, J.C. Gentina, I. Reyes and R. Torres et al., 1994. Production of penicillin acylase from Bacillus megaterium in complex and defined media. Process Biochem., 29: 263-270.
CrossRefDirect Link - McVey, C.E., S.P. Tolley, S.H. Done, J.A. Brannigan and K.S. Wilson, 1997. A new crystal form of penicillin acylase from Escherichia coli. Acta Crystallogr. Section D: Struct. Biol., 53: 777-779.
CrossRefDirect Link - Robas, N., H. Zouheiry, G. Branlant and C. Branlant, 1993. Improved penicillin amidase production using a genetically engineered mutant of Escherichia coli ATCC 11105. Biotechnol. Bioeng., 41: 14-24.
CrossRefDirect Link - Vojtisek, V. and J. Slezak, 1975. Penicillinamidohydrolase in Escherichia coli: II. Synthesis of the enzyme, kinetics and specificity of its induction and the effect of O2. Folia Microbial., 20: 289-297.
CrossRefDirect Link - Vojtisek, V. and J. Slezak, 1975. Penicillinamidohydrolase in Escherichia coli: III. Catabolite repression, diauxie, effect of cAMP and nature of the enzyme induction. Folia Microbiol., 20: 298-306.
CrossRefDirect Link - Wojskowicz, J., 1981. Regulation of penicillin amidase activity in Escherichia coli. Acta Microbial. Pol., 30: 13-23.
PubMedDirect Link