
Muhammad Anwer
Government M.D. College, Faisalabad, Pakistan
Irshad Hussain
Environmental Biotechnology Division, NIBGE, P.O. Box 577, Jhang Road Faisalabad, Pakistan
Tom McNeilly
Department of Biological Sciences, Nicholson Building, University of Liverpool, L69 3BX, Liverpool, UK
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 1 | Page No.: 78-81







ABSTRACT
An abandoned lead/zinc mine in Trelogan, Flintshire, North. Wales is the source of contamination of adjacent fertile agricultural land and water courses. In order to evaluate the zinc tolerance of pasture and mine populations of Anthoxanthurn odoratum that have colonized Trelogan mines, the root growth of pasture and mine sampled plants was examined, by charcoal staining method. Statistical analysis showed significant reduction in the root length of pasture and mine populations at various concentration of zinc. Mine populations were found to be more tolerant to zinc as compared with pasture populations of A. odoratum. The significant interaction between zinc concentration x pasture/mine genotypes indicated that different genotypes (clones) responded differently to the increasing zinc concentrations. Some genotypes were found to be more susceptible to zinc than others implying that there may be genetically based variation in zinc tolerance present in the Trelogan mine population of A. odoratum.
PDF Abstract XML References Citation
How to cite this article
Muhammad Anwer, Irshad Hussain and Tom McNeilly, 2000. Assessment of Zinc Tolerance of Pasture and Mine Populations of Anthoxanthum odoratum by Charcoal Staining Method. Pakistan Journal of Biological Sciences, 3: 78-81.
DOI: 10.3923/pjbs.2000.78.81
URL: https://scialert.net/abstract/?doi=pjbs.2000.78.81
DOI: 10.3923/pjbs.2000.78.81
URL: https://scialert.net/abstract/?doi=pjbs.2000.78.81
INTRODUCTION
Environments have been contaminated by heavy metals ever since the original magma of the earth solidified (Bradshaw, 1984). Without human impact excessive concentrations of some heavy metals in soils are the result of natural mineralisation caused by the presence of undisturbed ore bodies near the surface (Ernst, 1990). Many areas of metal contamination have arisen relatively recently. With the industrial development over the last 150 years there has been an increased release of metals during human-related activities. The sources of anthropogenic inputs of metals into the environment are numerous and diverse and include mining, smelting and the combustion of fossil fuels, pollution from car engines, sewage sludge disposal, power lines and from the use of pesticides and fungicides. This has lead to ever-increasing awareness of the potency of heavy metals as environmental pollutants. Elevated levels of the heavy metals are toxic and ultimately cause the death of the plants (Berry and Wallace, 1981). This toxicity leads to retardation and ultimately to complete inhibition of root growth at higher concentrations (Bradshaw and McNeilly, 1981; Symeonidis et al., 1985). The degree of toxicity of the metal depends both on the individual metal and upon the plant population itself (Symeonidis et al., 1985)). Generally, however certain metals are considered more toxic than the others and DeKock (1956) has given the order of toxicity for a range of higher plants as copper > nickel > zinc > lead.
Grime et al. (1988) reported that A. odoratum is found throughout the British Isles, Europe and temperate Asia, Australia and New Zealand. It frequently occurs on heaths, moors, hill grasslands, old pastures, woodlands and meadows on a wide range of soil types from sands to clay in dry and damp habitats. It is also frequently found near lead zinc mine spoil and scree slopes in the limestone dales, river banks, road verges and other waste places. A. odoratum is polymorphic and morphologically variable (Wu and Jain, 1980) and have population differentiation due to soil metal contamination (Antonovics and Bradshaw, 1970). A. odoratum is wind pollinated, protogynous and almost completely outcrossing (Grime et al., 1988). Antonovics (1968) suggested that a small degree of self fertility is also found for A. odoratum at Trelogan mine population. Half lives of this species varied from 0.95 years to 2.17 years depending upon the date of recruitment (Antonovics, 1972). He further observed that turnover of the population on the mine was rapid and only about 3 percent of the individuals lived longer than 5 years. The plant height is usually 150 mm but the flowering shoot has been recorded up to 500 mm (Grime et al., 1988).
Anthoxanthum odoratum plants at the Trelogan lead/zinc mine are known to be lead and zinc tolerant. Jowett (1964) used only one concentration of the metal for the tolerance testing. Some plants may show different responses to different concentrations of those metals found in the mine waste and possibly have some tolerance to metals which are not found in the mine, because mechanisms of Zn tolerance may affect tolerance to other metals, Cu and Ag. Von Insam and Hutchinson (1993) found that plants taken from mainly Ni, Co and As contaminated sites were not tolerant to Cu but showed increased tolerance to Zn and Pb.
MATERIALS AND METHODS
Tillers of twelve mine plants of A. odoratum, collected from the lead/zinc mine area at Trelogan, Flintshire, North Wales, namely T5, T10, T11, T12, T24, T27, C8, C13, C15, C26, Q4 and Q8, grown at the University Botanic Garden at Ness for more than 7 years, were used to check the zinc tolerance of respective genotypes. In addition, three pasture plants Pat1, Pat2 and Pat6, collected at the same time from a pasture closely adjacent to the mine waste and maintained at Ness Botanic Garden, were also used to check their zinc tolerance. All these plants had been already grown in 14 cm pots filled with John Innes compost.
Thirty tillers were taken from each plant and all existing roots were removed at the node at the base of each tiller. Forty five square plastic plates (10 x 10 cm) were used to support 10 section of plastic tubing 5.5 cm in length, each being used to support a single tiller. The basal node of the tiller was suspended in the appropriate test solution in 250 ml plastic beakers.
| Table 1: | Rorison's Nutrient Solution (0.1 strength) |
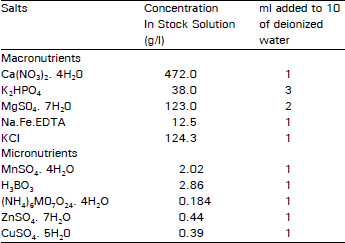 | |
The test solutions were prepared in 0.1 strength Rorison water culture nutrient solution (Hewitt, 1966, Table 1). Two different series of multiple concentrations of zinc sulphate were chosen for pasture and mine populations. Pasture tillers were tested across the sequence 95, 100, 105, 110, 115 and 120 mg l–1 of zinc sulphate (38.35, 40.37, 42.38, 44.40, 46.42, 48.44 mg l–1 of Zn ions respectively) and mine plants were tested across 190, 200, 210, 220, 230, 240 250 and 260 mg l–1 of zinc sulphate (76.70. 80.74, 84.77, 88.81, 92.85, 96.88, 100.92, 104.96 mg l–1 of Zn ions respectively). Pasture plants were grown at lower zinc concentrations than mine plants because preliminary experiments carried out at a range of concentrations indicated that pasture plants did not grow above 120 mg ZnSO4 per litre.
The experiment had three replicates, each having ten tillers per individual plant. Root growth of each tiller was measured 5 days after placement in initial zinc concentration in Rorison culture solution. After this period of exposure to the initial zinc concentration, the tillers were then kept in each increased Zn concentration for 2 days. Before placement in the following concentration, the roots of all the plants were stained using a stirred suspension of finally powdered active charcoal. This staining method allowed a rapid and highly accurate measurement of further root growth. The method did not cause any change in root structure (Schat and ten Bookum, 1992). After each measurement, the roots were restained and the test solution was replaced by one having the next higher zinc concentration at regular intervals of two days until root growth was completely inhibited.
The experiment was carried out in a growth room at temperature 24±2°C, with constant relative humidity of 65 percent and under constant illumination at 50 μmoles m–1 s–1 provided by cold white fluorescent lights. The experiment was arranged in a complete randomised block design.
RESULTS
Zinc tolerance of pasture plants: Zinc concentration, as expected, caused a significant (p<0.001) reduction on the root growth of pasture plants. The difference among the genotypes was also significant (p<0.001). The significant (p<0.001) interaction between zinc concentration x pasture genotypes, indicates that different genotypes (clones) responded differently to the increasing zinc concentrations (Table 2).
| Table 2: | Variance of Zn tolerance of pasture plants of A. ocloraturn |
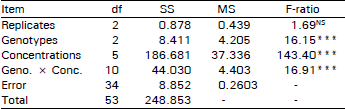 | |
| Table 3: | Analysis of variance of Zn tolerance of mine plants of A. oclaratum |
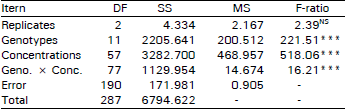 | |
| NS = Non Significant *** = Significant at p<0.001 | |
As far as the three pasture plants were concerned, the significant increased (p<0.05) response was attributed partly to Pat1 in comparison with the other two pasture ecotypes at 95 and 100 mg l–1 ZnSO4, after 5 and 7 days from the start of the test (Fig. 1). Pat1 was more tolerant genotype. However Pat6 was less tolerant than the other two genotypes at a concentration of 95 mg I–1 ZnSO4 but was more tolerant at 105 mg I–1 ZnSO4. This was probably responsible for the significant genotype × concentration interaction. The rooting of all three pasture plants, Pat1, Pat2 and Pat6 was inhibited at 120 mg ZnSO4 I–1. However, increase in root increment per two days in case of all the pasture plants declined with an increase in zinc concentration. The zinc tolerance of these pasture plants appears to decrease in the order Patl > Pat2 > Pat6. Furthermore at 95 mg ZnSO4 per litre the root increment of Pat1 was 2.67 times greater than that of Pat2 and 1.4 times higher to that of Pat6.
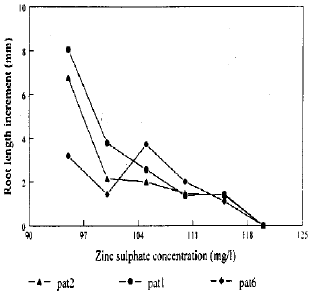 | |
| Fig. 1: | Zinc tolerance of pasture population of A. odoratum |
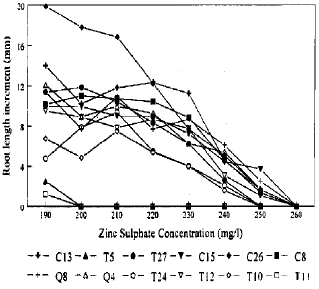 | |
| Fig. 2: | Zinc tolerance of mine population of A. odoratum |
Zinc tolerance of mine plants: Increasing concentrations of zinc significantly (p<0.001) reduced root length of mine plants. The difference among the Trelogan mine clones was also significant at the same probability. The interaction term genotype × concentration of zinc was also significant (p<0.001), indicating that different clones (genotypes) reacted differently to the increasing zinc concentrations (Table 3). Increment of root growth of Trelogan clones (Fig. 2) decreased as the zinc concentration increased. Among the mine plants the least tolerance was shown by clones T5 and T11 (Fig. 2). Root growth of these two genotypes ceased at 200 mg l–1 of ZnSO4, applied 7 days after the start of the test. Root growth of the mine clones T10, T24 and T27 stopped at 250 mg ZnSO4 I–1 while the rooting of genotypes of Q4, 08, C8, C13, C15, C26 and T12 stopped at 260 mg ZnSO4 I–1.
Fig. 1 clearly contrasts the pasture and mine plants of A. odoratum. The clones, T10, T24 and T27 stopped their root at double concentration as compared to the pasture plants. But the root growth of 04, Q8, C8, C13, C15. C26 and T12 ceased at more than double the concentration that stopped root growth of the pasture clones. It is clear that individual genotypes responded differently to the range of zinc concentrations. Some genotypes (e.g., T10) were more susceptible to zinc than others (e.g., C26 and C13, the relatively more zinc tolerant genotypes) implying that there may be genetically based variation in zinc tolerance present in the Trelogan mine population of A. odoratum.
Statistical analysis: The data were analysed using the two way ANOVA from PROC ANOVA with interaction components using the SAS package on the Unix System (SAS, 1989). For separation of means the LSD at 0.05 level was also computed (Steel and Torrie, 1980).
DISCUSSION
Tolerance to zinc was examined in a series of increasing concentrations of zinc based on serial replacement of metal solutions with the next higher concentration, after a specified interval, using the method of Schat and Bookum (1992). The technique used in this experiment gave results in the form of a critical concentration i.e. the concentration required to bring about rooting inhibition produced by the specified level of zinc toxicity.
Due to the sensitivity of pasture plants, the initial root growth of the tillers was carried out at 95 mg ZnSO4 per litre. While initial root growth of tillers of mine plants was carried out at 190 mg ZnSO4 I–1 due to their greater tolerance for zinc. This proved to be an appropriate range of concentration of zinc for these population.
Root growth of the tillers of both pasture and mine plants was reduced significantly (p<0.001) at different higher concentrations of zinc as shown in Table 2 and 3. These results were similar to those of Schat and Bookum (1992) who also observed root reduction in Silene vulgaris, while studying copper tolerance in this species.
Results clearly showed that mine plants tolerate double zinc concentration than the pasture plants. These results parallelled the findings of Symeonidis et al. (1985) and Von lnsam and Hutchinson (1993), although they used different methods for testing the heavy metal tolerance.
Two mine individuals, T5 and T11 stopped rooting at a concentration of 200 mg I–1 ZnSO4, but this may have been due to the tillers being weak and therefore producing short roots which stopped growing at the second zinc concentration. These two individuals may have been less tolerant than the other mine individuals or their physiological condition, when the experiment was started, may have been responsible for poor root growth. Similar MacNair (1983) reported that plants (M. guttatus) were unclassifiable as tolerant or non-tolerant due tb their short roots, no roots or plant condition.
It is worth noticing from the significant zinc concentrations x genotypes interaction term (Table 2 and 3) that genetic variation may exist within and between tolerant and non-tolerant clones of A. odoratum and these results in accordance with the findings of Karataglis (1978) who found that a great variation exists among the genotypes within the population of A. odoratum both for zinc and lead tolerance, using the old method of relative tolerance.
REFERENCES
- Antonovics, J. and A.D. Bradshaw, 1970. Evolution in closely adjacent plant populations VIII. Clinal patterns at a mine boundary. Heredity, 25: 349-362.
CrossRefDirect Link - Antonovics, J., 1968. Evolution in closely adjacent plant populations V. Evolution of self-fertility. Heredity, 23: 219-238.
CrossRefDirect Link - Berry, W.L. and A. Wallace, 1981. Toxicity: The concept and relationship to the dose response curve. J. Plant Nutr., 3: 13-19.
CrossRefDirect Link - DeKock, P.C., 1956. Heavy metal toxicity and iron chlorosis. Ann. Bot., 20: 133-141.
CrossRefDirect Link - Karataglis, S.S., 1978. Studies on heavy metal tolerance in populations of Anthoxanthum odoratimi. Plant Biol., 91: 205-216.
Direct Link - MacNair, M.R., 1983. The genetic control of copper tolerance in the yellow monkey flower, Mimulus guttatus. Heredity, 50: 283-293.
CrossRefDirect Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Symeonidis, L., T. McNeilly and A.D. Bradshaw, 1985. Interpopulation variation in tolerance to cadmium, copper, lead, nickel and zinc in nine populations of Agrostis capillaris L. New Phytol., 101: 317-324.
CrossRefDirect Link - Wu, L. and S. Jain, 1980. Self-fertility and seed set in natural populations of Anthoxanthum odoratum L. Bot. Gazette, 141: 300-304.
CrossRefDirect Link