
Mohammad A. Kamal
Department of Biochemistry, College of Science, King Saud University, P.O. Box 2455, Riyadh, 11451, Saudi Arabia
Abdulaziz A. Al-Jafari
Department of Biochemistry, College of Science, King Saud University, P.O. Box 2455, Riyadh, 11451, Saudi Arabia
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 5 | Page No.: 767-771







ABSTRACT
The current study addresses the kinetic analyses of the interaction of gallamine triethiodide with bovine retinal acetylcholinesterase (AChE, EC 3.1.1.7). The Michaelis-Menten constant (Ks) of the control for the hydrolysis of acetylthiocholine iodide (ASCh) by AChE was 0.072 mM and it was increased by 32.9-50.3% for the gallamine (0.05-1.5 mM) treated enzyme. The V, was 0.82 μmol/minlmg protein for the control system; it was decreased by 8.4-60.5% in the gallamine treated systems. The Lineweaver-Burk plot, Dixon plot and their secondary replots indicated that the nature of the inhibition was that of neither a linear nor a hyperbolic, conventional mixed type of inhibition system. We refer to this new mode of inhibition as a "non-linear-non-hyperbolic" mixed type inhibition, which is composed of a mixture of competitive and noncompetitive inhibition. The values for KI and KI were estimated to be 0.69 and 1.096 mM, respectively. These findings in the current study indirectly approach towards understanding of the binding sites of the bovine retinal AChE in the light of comparison with reported data for Torpedo, chicken and camel retinal AChE.
PDF Abstract XML References Citation
How to cite this article
Mohammad A. Kamal and Abdulaziz A. Al-Jafari, 2000. Mode of Inhibition of Bovine Retinal Acetylcholinesterase by Gallamine Triethiodide in vitro. Pakistan Journal of Biological Sciences, 3: 767-771.
DOI: 10.3923/pjbs.2000.767.771
URL: https://scialert.net/abstract/?doi=pjbs.2000.767.771
DOI: 10.3923/pjbs.2000.767.771
URL: https://scialert.net/abstract/?doi=pjbs.2000.767.771
INTRODUCTION
Acetylcholinesterase (AChE; EC 3.1.1.7) is a highly active enzyme for the hydrolysis of the neurotransmitter acetylcholine at the cholinergic synapses (turnover number is >104 s–1) and by this way terminates synaptic transmission (Kryger et al., 1998). The hydrolysis reaction proceeds stepwise, initiated by a nucleophilic attack due to the presence of nucleophilic serine of esteratic site of the AChE and carbonyl carbon of the acetylcholine (substrate), acylating the enzyme and liberating choline, which is an important molecule for the assay of this enzyme by the spectrophotometric method. This is followed by a rapid hydrolysis of the acylated enzyme yielding acetic acid and the restoration of the esteratic site of the enzyme. The presence of AChE activity in the retina was reported 55 years ago (Aneissen, 1944). A great deal of information is available on cholinergic drugs and their anticholinesterase effects on retinal functions (Rengstorff and Royston, 1976). Inhibition of AChE has been reported to either speed up or be reduced in the process of dark adaptation (Trussov et al., 1975). The reduction in visual activity and an increase in flicker fusion frequency have often been noted following exposure to anticholinesterases and attributed to changes in accommodative ability (Hutchins et al., 1995).
AChE is known to be a target enzyme for the discovery of ideal drugs for the treatment of a variety of neurological diseases such as Alzheimer =s, myasthenia gravis, Pick= s and Huntington =s (Al-Jafari, 1996). Therefore, a search for selective inhibitors of this enzyme has proven valuable for the improvement of current treatments against several human neurological disorders. Recently, the effect of gallamine triethiodide (GA) on camel retina AChE has been reported (Al-Jafari, 1997). Other names of GA are followings: tricuran, flaxedil, relaxan, retensin, while its chemical nomenclature is 2, 2', 2'-[1,2,3 Benzene-triyltris (oxy)] Iris [N, N, N-triethylethanaminium] triiodide). In the present study, we have assessed the effects of GA on bovine retinal AChE. The GA is a synthetic, non-depolarizing neuromuscular blocking agent and having medical application. It is usually given during surgery along with general anesthesia for muscle relaxation and often used as a "curare principle" for receptor blocking. The purpose of this attempt was to study the effect of GA on the cholinergic system of retinal tissue. Some difference in the mode of action of GA on AChE from camel and bovine retina is anticipated, because the camel eye has features of a diurnal eye and the camel retina presents a centrally elongated macular area in which mainly cone photo-receptors are found, whereas the bovine retinal structure slightly deviates from it. Moreover, there is no tapetum lucidum layer between choroid and retina as found in the eyes of large nocturnal mammals (Duhaiman, 1988).
MATERIALS AND METHODS
All reagents were of analytical grade. Acetylthiocholine (ASCh), gallamine triethiodide and 5,5-dithio-bis (2-nitrobenzoic acid) (DTNB) were purchased from Sigma Chemical Co. (St. Louis, MO, U.S.A.). Bovine serum albumin (BSA) and Triton X-100 were obtained from Fluke Chemika-Bio-Chemika, Switzerland.
Enzyme preparation: Retinas of young cows were obtained from a local abattoir. The whole retina from each eye was rapidly removed, rinsed in pre-chilled 0.9% saline solution, blotted and weighed. In each batch of enzyme preparation, twenty whole retinas were homogenized in pre-chilled 50 mM sodium phosphate buffer (pH 7.4). The extraction of retinal membrane-bound AChE has been described in the previous report (AI-Jafari et al., 1998).
Assay of AChE Activity: AChE activity was determined by the spectrophotometric method of Ellman et al. (1961). The assay mixture contained 0.25 mM ASCh and 0.25 mM DTNB. A 4 min. incubation time was selected for the enzyme assay at 25̊C. after preliminary experiments were performed to ensure that enzyme activity was linear with respect to the reaction time and enzyme concentration employed. To study the effect of gallamine, the enzyme was pre-incubated with gallamine for 4 min. prior to the addition of substrate. A yKs represents the dissociation constant of AChE-ASCh-gallamine complex into the AChE-gallamine complex and ASCh. The inhibition constant K, represents the dissociation constant of the AChE-ASCh-GA complex into the AChE-ASCh complex and free gallamine.
Protein estimation: The protein content of enzyme preparation was estimated with the method of Lowry et al. (1951) using BSA as a standard. Triton X-100 interfered with this assay; therefor, the colored precipitate was removed by centrifugation as described previously (Kamal and Al-Jafari, 1996).
Statistical Analysis: The data were plotted by using the GraFit program (Leatherbarrow, 1992).The values of the correlation coefficient, slope, intercept and their standard errors were obtained by the linear regression analysis of this program.
RESULTS
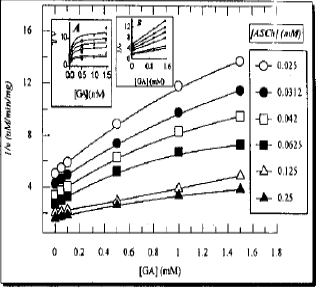
The Lineweaver-Burk plot without regression analysis is presented in Fig. 1. After linear regression analysis all lines became straight and are presented in box of Fig. 1. The 1/v-axis intercept, which is an ordinate of the Lineweaver-Burk plot is shown in a separate plot (Fig. 2). Its reciprocal form without regression analysis is presented in inset A. The comparison indicated that there is no difference in shape of the curve of main plot and its reciprocal form. However, its linear regression analysis showed a straight line (inset B). The slopes of the lines in Lineweaver-Burk plot were also unusual. Therefore, these were analyzed by three different .ways as shown for 1/v - axis intercept (Fig. 3, inset A and El). The Dixon plot was also unusual. Therefore, it was expressed by three different ways in Fig. 4: 1. Main plot after polynomial regression analysis; 2. Original data in inset A and 3. Plot after linear regression analysis of the original data (inset B). The 1/v-axis intercept which is an ordinate of the lines in the Dixon plot, the slope of the lines in the Dixon plot and Kapp were plotted by two different ways: 1. The values before the regression analysis are shown in the main plot of Fig. 5 and 2. the values after the regression analysis- are presented in the inset of Fig. 5.
Data in Table 1 show that GA between 0 and 1.50 mM concentration, increases Ksapp whereas it decreases the Vmss by 8.4 to 60.5%. The type of inhibition produced by GA was assessed by a Lineweaver-Burk plot as well as by Dixon plots. As shown in Fig. 1-5, GA inhibited AChE activity as a mixed type inhibitor. This mixed type inhibition system was composed of competitive and noncompetitive-type inhibition. The Kapp for each ASCh concentration was estimated by regression equation and is presented in Table 2. As shown in Table 2, the K1spp slightly increased between 0.025 to 0.031 mM concentrations of ASCh and it decreased by 3.23% at 0.042 mM ASCh. It increased by 6.79 to 23.25% from 0.063 to 0.25 mM concentration of ASCh. The Vmaxiapp was increased by 19.25 to 215.51% over the entire range of ASCh concentration. The values of K, and K, were calculated from secondary replots of slope and /Vmass, values from the Lineweaver-Burk plot (Fig. 2, 3), original Dixon plot (Fig. 4 inset A) and from the linear regression equation for primary Dixon plots for each ASCh concentration in the Dixon plot at a fixed GA concentration (Fig. 4 inset B). The values of K, was also calculated from Fig. 5. The Ks and yKs values of the bovine retinal AChE were calculated from the mean of the primary double reciprocal plot (Fig. 1 inset) and secondary replots (Fig. 5 inset o, for Ks and for yKs). The values for the kinetic parameters, K, K1, yKs and Vmax were calculated by different methods and are summarized in Table 3.
DISCUSSION
The Lineweaver-Burk plot (Fig. 1) did not follow the simple intersecting linear rules of inhibition, i.e. competitive, noncompetitive or uncompetitive. From these curves, it is reasoned that GA may produces a mixed type of inhibition. In case of a mixed type of inhibition, the curve is generally either hyperbolic or linear. These two types can be distinguished on the basis of the shape of their Dixon plots, If it is curved, it is a hyperbolic mixed type inhibition system. In our case, the Dixon plot was neither linear nor smooth curved (Fig. 4 inset A). We refer to this type of kinetic profile as a "non-linear-non-hyperbolic" (NLNH) mixed type inhibition system. In the present study, the profile seems to be of a mixed-type of inhibition composed of competitive and noncompetitive elements.
The competitive type also has two subtypes, partial and pure, These can be distinguish from the shape of slope1/S-axis, versus the inhibitor concentrations replots.
| Table 1: | Effect of gallamine (GA) on kinetic parameters of bovine retinal AchE |
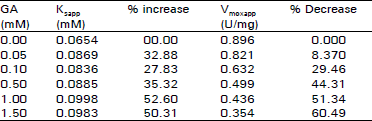 | |
The Ksspp and Vmaxspp were determined by their respective regression equation as well as by the Lineweaver-Burk plot (Fig. 1). The Ksspp is equal to reciprocal of x-intersection while Vmaxapp isequal to receiprocal of y-intersection of each line for each GA concentration in the Lineweaver-Burk plot (each point in this figure, represents the mean of four independent determination). | |
| Table 2: | Kinetic data from Dixon plot for bovine retinal aChE inhibition by gallamine |
 | |
The Klapp and Vmaxiapp were determined by the their respective regression equation and Dixon plot (Fig. 4 inset B). The Klapp is equal to x-intersection while Vmaxiapp is equal to receiprocal of y-intersection of each line for each ASCh concentration in the Dixon plot (each point in this figure, represents the mean of four indepedent determinations). | |
As indicated in Fig. 3, the Slope1/S-axis versus GA plot was linear in the region of two sets of concentrations i.e. lower and higher concentrations (0.0 to 0.10 mM and 0.50 to 1.5 mM), However, the slopes of the plot are different. Consequently, when plotted together, one observes a curve due to the overall slope values. The critical concentration could be readily observed after extrapolation from the linear portion, which was found to be 0.10 mM GA. For the assessment of partial inhibition, the reciprocal form of the slope1/S-axis versus the reciprocal form of the GA concentrations were replotted (Fig. 3), the plot hyperbolic inconsistent with the case of partial competitive inhibition. After linear regression analysis the same plot was replotted in Fig. 3 and the correlation coefficient was estimated to be 0.959. It was therefore, concluded that competitive inhibition, which is a part of NLNH system, is neither partial nor pure.
As in the competitive type of inhibition, the noncompetitive type also has two sub-types, partial and pure. These can be distinguished on the basis of replots of 1/Vmaxapp versus GA concentrations. As described above, again the 1/Vmaxapp versus GA concentration plot was linear in the region of two sets of concentrations (0.0 to 0.10 mM and 0.50 to 1.5 mM) and the slopes of linearly were different. Therefore, when plotted together, it looked like a curve due to the overall slope values. The critical concentration could be readily observed after extrapolation of dotted lines, which was found to be 0.10 mM GA. For the determination of partial inhibition, the reciprocal form of the 1/Vmaxapp, versus the reciprocal form of the GA concentrations were replotted (Fig. 2). Again the plot was not linear, as would be expected in case of partial competitive inhibition. Alternatively, after linear regression analysis, the same plot was replotted in Fig. 2 and the correlation coefficient was determined to be 0.972.
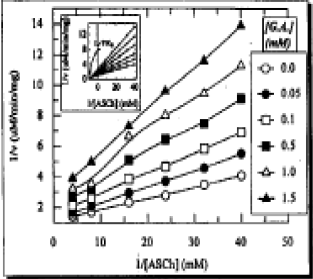 | |
| Fig. 1: | Inhibition of bovine retinal AChE by GA: Lineweaver-Burk plot of 1/v against 1/[IASCH] in the absence (o) and presence of GA concentration. Each point represents the mean of four independent determinations. (Inset) Shows the same Lineweaver-Burk plot after linear regression analysis for the determination of Ksapp and Vmaxapp. The correlation coefficients are above 0.991 |
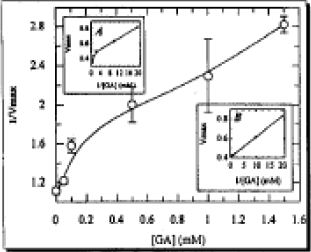 | |
| Fig. 2: | Inhibition of bovine retina AchE by GA: Secondary replot of 1/Vmaxiapp, (∘) from the Lineweaver-Burk plot against GA concentration for the determination of KI. The main plot is without linear regression analysis. Insert plot (A) is reciproal form of the main plot, while (B) is the same as A plot after linear regression analysis, where correlation coefficient was 0.972 |
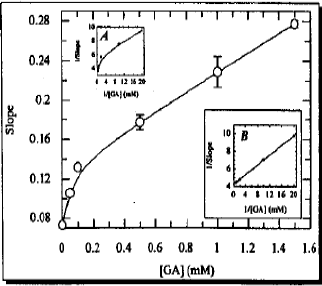 | |
| Fig. 3: | Secondary replot of the slope from the Lineweaver-Burk plot (without linear regression analysis) against GA concentration for exploring the mode of inhibition. Inset (A) is reciprocal of slope versus reciprocal for of the GA concentration while inset (B) is the same as A plot after linear regression analysis, where correlation coefficient was 0.959 |
 | |
| Fig. 4: | Dixon plot for representation of the inhibition of bovine retina AchE by GA at six different fixed concentration of ASCh in presence of 0-1.5 mM GA concentration after polynomial regression analysis. Each point represents the mean of four independent determinations. (Inset A) Shows the original Dixon plot without any regression analysis. (Inset B) Represented the Dixon plot after linear regression analysis for the determination of Ksspp and Vmaxiapp. The correlation coefficient was as follows: (▲), 0.9745; (▵), 0.9678; (■), 0.9633; () 0.975; (•), 0.9777 and (o), 0.9756 |
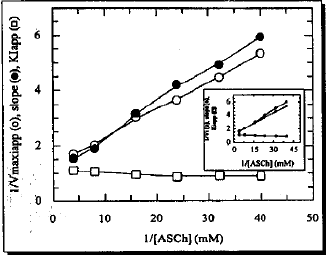 | |
| Fig. 5: | Secondary Dixon plot Replot of 1/Vmaxiapp slope and Kapp against 1/[ASch], (Insert Plot) is after linear regression analysis for the determination of Ks, yKs and K1 respectively. Correlation coefficient was 0.9986, 0.9968 and 0.8644 for 1/Vmaxiapp, slope and Ksspp respectively |
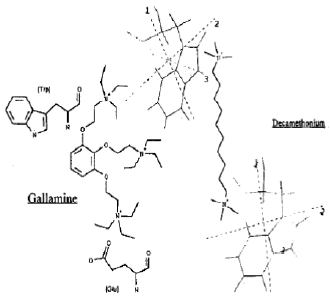 | |
| Fig. 6: | Chemical Structure of gallamine and decamethonium |
| Table 3: | Kinetic constants estimeted by various methods |
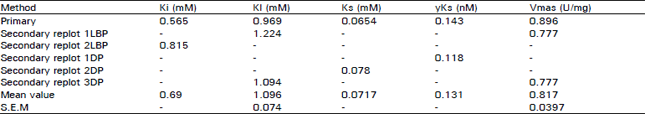 | |
The details of primary plot and secondary replots have been given in thetext whileLBP means Lineawaver-Burk plot and DPmeans Dixon plot. represents replot fo 1/Vmaxapp versus GA concentration (Fig. 2) and2LBP represents replot of slope from Lineweaver-Burk plot versus GA concentration (Fig. 3). 1DPrepresents replot of slope from Dixon plot versus 1/[ASch], 2DPrepresents replot of 1/Vmaxiapp versus 1/[ASch] and3DP represents replot of Ksspp from Dixon plot versus 1/[ASCh] (Fig. 5) | |
Therefore, on the basis of this replot, the noncompetitive component of NLNH system was determined to be neither pure nor partial in nature. These results indicate that GA behaved unusually with bovine retinal AChE and a mixed type of inhibition (NLNH) was confirmed.
Ai discussed above, the same problem was found with the Dixon plot (Fig. 4 inset A), i.e. the 1/v versus GA concentration plot was only linear in the region of two sets of concentrations (0.0 to 0.10 mM, and 0.50 to 1.5 mM), but the slopes were different. Therefore, when plotted together, it looked like a non-smoothly curve due to having two linear regions instead of gradual curvature. The critical transition concentration could be readily observed by extrapolation of dotted lines in a magnified plot (riot shown) and this was 0.10 mM GA.
Regarding the sensitivity of AChE (from various sources) towards GA, it has been reported that a concentration of GA between 0.0001 mM and 0.1 mM has no effect on the rate of hydrolysis of acetylcholine either by membrane-bound or by solubilized Eel AChE. The kinetic parameters specifically Km and Vmax were also not affected (Robaire and Kato, 1975). GA was additionally reported to increase the rate of the catalytic hydrolysis at a low concentration and decrease it at higher concentrations. This, however, was not observed in the current study. Concentrations of GA above 10 mM affected acetylcholine hydrolysis, but complete inhibition was not attained. Concentrations exceeding 10 mM had no further effect on the AChE activity (Roufogalis and Wickson, 1975).
The KI values for GA on Torpedo, chicken and camel retinal AChE were reported to be 0.015 mM, 0.74 mM (Eichler et al., 1994) and 0.160 mM (Al-Jafari, 1997) respectively. In the present study, it was found to be 0.69 mM, close to that of chicken AchE. This observation indicates that, the KI value for bovine retina AChE is 46-fold the value of Torpedo and 4.3-fold higher than camel retinal AChE. This finding demonstrates that bovine retinal AChE has a lower affinity for GA than does Torpedo AChE and camel retinal AChE. Although, its affinity, however, is 4.3 - fold lesser than camel retinal AChE, One other interesting finding is that GA inhibits camel retinal AChE competitively, and inhibits Torpedo and chicken AChE non-competitively. The inhibition of purified bovine erythrocyte AChE by GA was reported to be of a mixed linear type. The competitive inhibition constant (KI) was reported to be 1.94 μM. The uncompetitive inhibition constant (KI) was not determined because it increased the Vmaxiapp rather than decreased it (Dawson et al., 1991). This indicates that AChEs may have different amino acids or the same amino acids but different sequences within its active-site gorge and at the peripheral anionic site in different species, because of its inhibition pattern varies from source to source for GA. It is possible that Tyr-70 and Trp-279 may be absent in the peripheral site of bovine retinal AChE, like in chicken AChE (Eichler et al., 1994). Due to a mixed type inhibition of bovine retinal AChE by GA, the peripheral site may be unsuitable for accommodation of GA. Therefore, GA may be attracted towards the anionic site within the active site gorge due to competitive component of mixed type of inhibition.
Generally, the active site of AChE has three major domains: (1) an esteratic locus, containing serine, (2) an anionic locus 4-5 A° away from the esteratic locus, and (3) a hydrophobic region that is contiguous with or near the esteratic and anionic loci. This region is important for binding an aryl substrate and an active site ligand (Quinn, 1987). The anionic site consists of carboxylated ions derived from the side chain of glutamic or aspartic residues. The ASCh binds with the first two loci, N+ of the choline of ASCh binds to the anionic site, while its acetate binds to the esteratic site. The competitive part of the mixed type of inhibition of GA can be explained in terms of its three - fold structural resemblance of three triethyi ammonium groups (i.e. three quaternary nitrogen atoms with three positive charges) to N+ of the choline of ASCh.
The affinity of GA towards the catalytic anionic site as well as the peripheral site has revealed that GA has some structural complementarity to the active as well as peripheral site of bovine retinal AChE. Because of the positive charge density and benzyl group of GA, it can be concluded that the catalytic anionic site as well as peripheral anionic site of bovine retinal AChE may be located in a more polar environment and their distance seems to be less than 12 A°. Because decamethonium which has 10 methylene groups, corresponding to 12 A° and it has ability to span the two anionic sites of Torpedo AChE (Eichler at al., 1994). The distance between the quaternary groups (top and bottom) of GA may correspond to approximate 9 methylene groups according to its structure (Fig. 6). Therefore, it is possible that one top quaternary group of GA may bind with entrance site of the gorge of bovine retinal AChE enzyme, while bottom group linked with catalytic anionic site due to slightly shorter distance between the two anionic sites as compared to Torpedo AChE.
REFERENCES
- Al-Jafari, A.A., 1996. The nature of the inhibition of camel retina acetylcholinesterase (EC 3.1.1.7) activity by tetrahydroaminoacridine. J. Ocular Pharmacol. Ther., 12: 503-514.
CrossRefDirect Link - Al-Jafari, A.A., 1997. The inhibitory effect of the neuromuscular blocking agent, gallamine triethiodide, on camel retina acetylcholinesterase activity. Toxicol. Lett., 90: 45-51.
CrossRefDirect Link - Al-Jafari, A.A., M.A. Kamal and A.S. Alhomida, 1998. On the inhibition of camel retina acetylcholinesterase activity by cycloheximide in vitro. Cell Biol. Toxicol., 14: 167-174.
CrossRefDirect Link - Dawson, R.M., M.H. Dowling and M. Poretski, 1991. Assessment of the competition between tacrine and gallamine for binding sites on acetylcholinesterase. Neurochem. Int., 19: 125-133.
CrossRefDirect Link - Duhaiman, A.S., 1988. Purification and some properties of camel lens crystallins. Curr. Eye Res., 7: 871-876.
CrossRefDirect Link - Eichler, J., A Anselment, J.L. Sussman, J. Massoulie and I. Silman, 1994. Differential effects of Peripheral site ligands on torpedo and chicken acetylcholinesterase. Mol. Pharmacol., 45: 335-340.
Direct Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Hutchins, J.B., J.M. Bernanke and V.E. Jefferson, 1995. Acetylcholinesterase in the developing ferret retina. Exp. Eye Res., 60: 113-125.
CrossRefDirect Link - Kamal, M.A. and A.A. Al-Jafari, 1996. The preparation and kinetic analysis of multiple forms of human erythrocyte acetylcholinesterase. Prep. Biochem. Biotechnol., 26: 105-119.
CrossRefDirect Link - Kryger, G., I. Silman and J.L. Sussman, 1998. Three-dimensional structure of a complex of E2020 with acetylcholinesterase from Torpedo californica. J. Physiol.-Paris, 92: 191-194.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Quinn, D.M., 1987. Acetylcholinesterase: Enzyme structure, reaction dynamics and virtual transition states. Chem. Rev., 87: 955-979.
CrossRefDirect Link - Trussov, M.S., R.H. Rengstorff and V. Gurvis, 1975. Effects of eserine upon light sensitivity and dark adaptation. Am. J. Optom. Physiol. Optics, 52: 438-441.
Direct Link - Rengstorff, R.H. and M. Royston, 1976. Miotic drugs: A review of ocular, visual and systemic complications. Optom. Vision Sci., 53: 70-80.
Direct Link - Robaire, B. and G. Kato, 1975. Effects of edrophonium, eserine, decamethonium, d-tubocurarine and gallamine on the kinetics of membrane-bound and solubilized eel acetylcholinesterase. Mol. Pharmacol., 11: 722-734.
Direct Link - Roufogalis, B.D. and V.M. Wickson, 1975. Acetylcholinesterase: Specificity of the peripheral anionic site for cholinergic ligands. Mol. Pharmacol., 11: 352-360.
Direct Link