
Awatif I. El-Nahas
Biological Sciences and Geology Department, Faculty of Education, Ain Shams University, P. N. 11341, Heliopolis (Roxy), Cairo, Egypt.
Sayed. F. Khalifa
Botany Department, Faculty of Science, Ain Shams University, Abassia, Cairo, Egypt
Maged. M. Abou-El-Enain
Biological Sciences and Geology Department, Faculty of Education, Ain Shams University, P. N. 11341, Heliopolis (Roxy), Cairo, Egypt.
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 5 | Page No.: 717-720







ABSTRACT
Comparative karyological studies for 23 taxa of the Solanaceae have been presented and their taxonomic aspects are discussed. It was concluded that the origin of the family is a taxon with basic chronomosome number of x = 7 or 8 which gave rise to other taxa by aneuplodiy and polyploidy. Nictiana elate and Browallia viscose are found to be less evolved than the other species studied. Concerning the total form percentage (TF%) a reverse trend towards increasing the degree of karyotype symmetry with advancing evolution is concluded.
PDF Abstract XML References Citation
How to cite this article
Awatif I. El-Nahas, Sayed. F. Khalifa and Maged. M. Abou-El-Enain, 2000. Karyological Studies on the Solanaceae. Pakistan Journal of Biological Sciences, 3: 717-720.
DOI: 10.3923/pjbs.2000.717.720
URL: https://scialert.net/abstract/?doi=pjbs.2000.717.720
DOI: 10.3923/pjbs.2000.717.720
URL: https://scialert.net/abstract/?doi=pjbs.2000.717.720
INTRODUCTION
The Solanaceae is one of the most important and large plant families, widely distributed throughout tropical and temperate regions (Rendle, 1952; Willis and Shaw, 1966; Lawrence, 1969). D’Arcy (1979) estimated it to contain -83 genera and 2671 species, while in 1991 he estimated 60 genera and about 1746 species for the solanoideae. According to Naik (1984) the family composed of 88 genera and about 2300 species. From the cytological point of view, Gupta (1969) studied some members of this family. Al-Nowaihi and Khalifa (1974), Hunziker (1979), Tetenyi (1987) and Mourad (1988) studied some of the systematic relationships in this family. Gottschalk (1954) and Philomina (1980) studied the interrelationships within the family based on variation of heterochromatin content and chromosome number, respectively. However, the karyological studies on the family are quite little. The present work deals with using the karyotypic analysis data as a mean for studying taxonomy of this family and assessment of their phylogeny.
MATERIALS AND METHODS
The different taxa studied and their sources are given in Table 1. Cytological preparations were carried out on root tips obtained from seeds germinated on moist filter paper in Petri dishes pretreated with 0.05% colchicine solution for 4 hours and fixed in 3:1 (v/v) ethanol : glacial acetic acid for 24 hours. Temporary preparations were made using the Feulgen squash technique and well spread cells were photographed. Some karyotype criteria were measured from 5 to 10 chromosome complements. These are: mean chromosome length in pm, total chromosome length, total short arm length, total long arm length, average arm ratio (Long/short) and TF% (Huziwara, 1962). Karyotype formula is derived from the number of chromosome pairs, chromosome type based on-arm ratio (Levan et al., 1964) and mean chromosome length (MCL). Chromosomes are categorized in the formula according to their mean length values into five types as follows: A = very long chromosomes (above 4.0 μm), B = long chromosomes (above 3.0 to 4.0 μm), C = medium length chromosomes (above 2.0 to 3.0 μm), D = short chromosomes (above 1.0 to 2.0 μm) and E = very short chromosomes (up to 1.0 μm).
RESULTS AND DISCUSSION
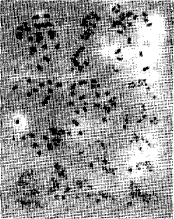
Chromosome complements are illustrated in Figures 1-4 and karyotypes are diagramatically represented by idiograms (Fig. 5). The karyotype analysis of the species studied shows that the basic chromosome number of x = 12 was recorded in 21 taxa representing seven of the eight examined genera. The lowest number (x = 9 + B) was recorded in Nicotiana elate, while x = 11 is found in Browaffia viscose. Except the presence of single B-chromosome in N. elate, the recorded chromosome number here confirms the previous reports. B-chromosome in the present study was single, submedian, shorter than the shortest chromosome in the complement and more or less heterochromatic. 8-chromosomes have previously been considered as evidence either for occurring as aneuploidy (Jackson, 1960) and presence of ancestor with low basic chromosome number for a certain species (Chinnappa and Morton, 1979), yet it can be assumed that N. alata might have been arisen through aneuploid alteration process and is a surviver from an ancestor having a low basic chromosome number which gave rise to the species.
According to Stebbins (1974) hypothesis concerning chromosomal evolution in most woody orders of angiosperms, the present data can be used to propose the phyletic status of the taxa studied based on the variation in basic chromosome number (Fig. 6). Thus, it is assumed that the Solanaceae might have originated from a taxon with basic chromosome number of x = 7 or 8 which gave rise to other taxa by aneuploidy and/or polyploidy. By advancing evolution in different secondary lines, diversification of chromosome complement could be predicted to occur through additional cycles of polyploidy or reduction in chromosome number after polyploidy. The chromosomal modifications in the different taxa were usually associated with structural changes as fragmentation that lead to overall decrease in the chromosome length (Haque and Ghoshal, 1980). Gupta and Gupta (1978) and Moore (1978) suggested that the reduction in chromosome length is associated with increasing evolution and that, the occurrence of polyploidy in a certain taxon is an evidence to be an advanced form. According to the above mentioned opinions it can be concluded that, the two tetraploid species Nicotiana tabacum and Withania somnifera (2n = 4x = 48) are more evolved than the other species studied. Otherwise, Nicotiana elate (i.e. more primitive) than the other species studied.
Measurements of mean chromosome length (MCL) values (Table 1) revealed that, the highest value (2.76 pm) was recorded in Browaffia viscose while the lowest value (1.08 pm) is found in Lycopersicon lycopersicurn cv. Pakmor B. L. The similarity between mean chromosome values for each of D. stramonium (MCL = 1.25 μm) and D. tatula var. inermis (MCL = 1.23 μm) leads to overlook the following views: (a) Wettstein (1895) and Soueges (1907) considered D. stramonium and D. tatula as two species. (b) But, on genetical basis both are shown as one species (Clarke, 1885; Bateson and Sounders, 1902; Avery et al., 1959). (c) Moreover, Al-Nowaihi and Khalifa (1974), on the basis of numerical analysis, accepted the latter view. They also reported that D. tatula tends to join with D. stramonium since the fruit in both is spiny. Similarly, D. tatula var. inermis tends to join with D. stramonium var. inermis (Fruit in both is smooth). Meanwhile, Avery et al. (1959) concluded that the fruit texture, in either case, is due to a recessive gene.
 | |
| Fig. 1: | Browallia viscose. 2, Capsicum annum. 3, C. Frutescense cv. Early Jalpino. 4, C. frutescense cv. Long Green. 5, C. frutescense cv. Long Red. 6, Datum innoxia Mill. 7, D. stramonium. 8, D. stramonium var. inermis staff. 9, D. tatula var, inermis. 10, Lycopersicon lycopersicum. 11, L. lycopersicum cv. Pakmor. 12, L. lycopersicum cv. Strain. |
 | |
| Fig. 2: | 13, Nicotiana acuminate. 14, N. alata 15. N. glauca. 16, N. peniculata. 17, N. tabecum. 18. Physhs alkekengi. 19, P. lxocarpa. 20, Solanum melongena cv. Black Beuty. 21, S. melongena cv. Long purple, 22, S. melor gena cv. Long white. 23, Withen a somnifera |
 | |
| Fig. 3: | Camera Lucida drawings to illustrate the chromosome complements of the taxa numbered as Fig. 1. |
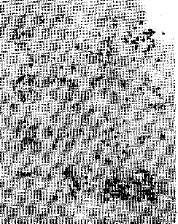 | |
| Fig. 4: | Camera Lucida drawings to illustrate the chromosome complements of the taxa numbered as Fig. 2. |
 | |
| Fig. 5: | Idiograms of the taxa studied, (numbered as Table 1 |
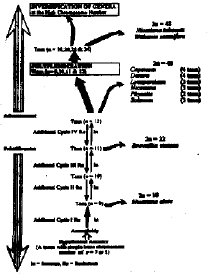 | |
| Fig. 6: | A proposed phyletic status of the taxa studies (Based on basic chromosome numbers). |
| Table 1: | Sources of the taxa studied and their somatic chromosomal criteria |
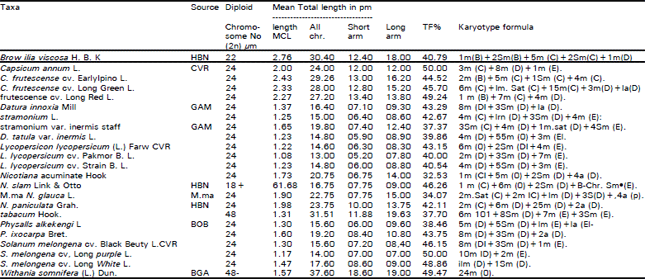 | |
HBN = National Botanical Garden, Belgium; CVR - Centre of Vegetable Research, Dakki, Giza, Egypt; GAM = Garden of Aromatic and Medicinal plants, Delta Barrage, Egypt: M.ma = Western Mediteranean Costal Region of Egypt; BGB = Botanical Garden of Berlin University, Germany. BGA = Botanical Garden of Faculty of Science. Ain Shams Univ., Cairo, Egypt. TF% (Total form percentage) = (Total sum of short arm length/Total sum of chromosome length x 100 (Huziwara, 1962). m = Metacentric or median, Sm = Submetacentric, or submedian, a = acrocentric or sub-terminal centromere, B-chr. = B-chromosome, B = Long chromosome (above 3 to 4 μm), C = Median (above 2.0-3.0 μm). D - Short (above 1 to 2 μm) and E = Very short (up to 1 μm), Sat. = Satellite. | |
The present data as well as those obtained from the above mentioned views, substantiated the assumption that these two species are derived from a unique species, which support their categorization as two varieties under the same species. Within the genus Nicotiana, mean chromosome length values (Table 1) reveals that N. paniculata (MCL = 1,98 μm) and N. glauca (MCL = 1.90 μm) are less evolved than N. acuminate (MCL = 1.73 μm), N. elate (MCL = 1.68 μm) and N. tabacum (MCL = 1.31 μm), whereas the latter species is the most evolved one. Chromosome morphology based on centromere position as indicated by karyotype formula for each of the taxa studied (Table 1) revealed that Browallia viscose and Capsicum fruitescence are more primitive compared to the remaining species due to the presence of long metacentric and submetacentric chromosomes. On the other hand, Physalis alkekengi is more advanced species due to presence of the two very short (E) submetacentric (sm) and acrocentric (a) chromosomes. Mean while the short acrocentric chromosomes recorded in each of Datura innoxia (one pair), Nicotiana paniculata (two pairs), N. glauca (four pairs), N. acuminata (four pairs) and Physalis ixocarpa (two pairs) are evidently advanced states. However, the presence of the medium sized metacentric chromosomes in each of Nicotiana paniculata (two pairs), N. glauca (two pairs) and N. acuminate (one pair) gives the possibility of intermediate evolutionary position for these species.
TF% in the taxa studied indicates that, the highest value (50%) is recorded in each of Capsicum annum and Solanum melanogena cv. long purple, while the lowest value (32.53%) is observed in Nicotiana acuminate. Contrary to the most common evolutionary concept that, the high degree of karyotype symmetry is only found in the primitive species (Levitsky, 1931; Babcock, 1947a, b; Moore, 1978) the present data revealed a reverse trend towards increasing degree of karyotype symmetry with advancing evolution as revealed by the recorded high degree of symmetry (TF% = 49.47) in the karotype of Withania somnifera which has been considered as relatively advanced species based on their diploid chromosome number (2n = 4x = 48) and mean chromosome length value (MCL = 1.75 μm). On the other hand, a lower degree of karyotype symmetry (TF% = 40.79%) is recorded in Browallia viscose which has been considered as a relatively primitive species on the basis of their chromosome number (2n = 2x = 22) and measurement of mean chromosome length value (MCL = 2.76 μm). Based on the argument that, the species studied are possibly derived from a primitive species, Withania somnifera could have retained its ancestral metacentric chromosomes as a result of chromosomal changes during the evolutionary process. The reverse trend of increasing degree of karyotype symmetry with advancing evolution has also been reported in several Gymnospermous plants especially in the family Podocarpaceae (Hair and Beuzenberg, 1958; Stebbins, 1974).
REFERENCES
- Al-Nowaihi, A.S. and S.F. Khalifa, 1974. A numerical classification of Solanceae in Egypt. J. Indian Bot. Soc., 53: 224-248.
Direct Link - Chinnappa, C.C. and J.K. Morton, 1979. The occurrence and behavior of B-chromosomes in Astranthium integrifolium (Compositae). Am. J. Bot., 66: 471-473.
Direct Link - Gupta, S.B., 1969. Duration of mitotic cycle and regulation of DNA replication in Nicotiana plumbaginifolia and a hybrid derivative of N. tabacum showing chromosome instability. Can. J. Genet. Cytol., 11: 133-142.
CrossRefDirect Link - Gupta, R. and P.K. Gupta, 1978. Karyotypic studies in the genus Crotalaria Linn. Cytologia, 43: 357-369.
CrossRefDirect Link - Hair, J.B. and E.J. Beuzenberg, 1958. Chromosomal evolution in the podocarpaceae. Nature, 181: 1584-1586.
CrossRefDirect Link - Haque, M.S. and K.K. Ghoshal, 1980. Karyotypes and chromosome morphology in the genus Salvia Linn. Cytologia, 45: 627-640.
CrossRefDirect Link - Huziwara, Y., 1962. Karyotype analysis in some genera of compositae. VIII. further studies on the chromosomes of aster. Am. J. Bot., 49: 116-119.
Direct Link - Levan, A., K. Fredga and A.A. Sandberg, 1964. Nomenclature for centromeric position on chromosomes. Hereditas, 52: 201-220.
CrossRefDirect Link - Tetenyi, P., 1987. A chemotaxonomic classification of the Solanaceae. Ann. Missouri Bot. Garden, 74: 600-608.
CrossRefDirect Link