
Sanaa T. El-Sayed
Biochemistry Department, National Research Center, Tahrir St, Dokki, Cairo, Egypt
Ahmed M. Salem
Biochemistry Department, Faculty of Science, Ain Shams University, Egypt
Abeer N. Shehata
Biochemistry Department, National Research Center, Tahrir St, Dokki, Cairo, Egypt
Etidal W. Jwanny
Biochemistry Department, National Research Center, Tahrir St, Dokki, Cairo, Egypt
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 2 | Page No.: 250-256







ABSTRACT
The activity of enzyme hydrolyzing colloidal chitin has been measured on extracts from a number of higher plants belonging to eleven families. Chitinase activity could be detected in dry seeds which increased during germination. Its relationship to different vegetative plant tissues was investigated. Sugar beet leaves (Beta vulgaris) provided the best active chitinase at optimum extracting conditions (extracted with water, at 16°C for 90 min).
Chitinase from leaves of sugar beet was purified by (NH4)2SO4 precipitation (20-60%) and fractionated on Sephadex G-120 followed by Sephadex G-200 column chromatography. A 18.9 fold purification of the enzyme with 14.0 ImU/mg specific activity was achieved. The yield of the purified chitinase was 43.4 mg protein (608 ImU) from 100g dry leaves tissues. The prepared enzyme was showing a single protein band on agarose gel electrophoresis. The purified enzyme had been shown to have a M, 64×103 Dalton on the basis of gel filtration on Sephadex G-200 column. Optimum chitinase activity on chitin was recorded in 0.1M acetate buffer, pH 4.5 at 40°C.
PDF Abstract XML References Citation
How to cite this article
Sanaa T. El-Sayed, Ahmed M. Salem, Abeer N. Shehata and Etidal W. Jwanny, 2000. Chitinase from Leaves of Beta vulgaris and other Higher Plants. Pakistan Journal of Biological Sciences, 3: 250-256.
DOI: 10.3923/pjbs.2000.250.256
URL: https://scialert.net/abstract/?doi=pjbs.2000.250.256
DOI: 10.3923/pjbs.2000.250.256
URL: https://scialert.net/abstract/?doi=pjbs.2000.250.256
INTRODUCTION
Chitinase (EC. 3. 2. 1. 14) is a basic protein that catalyses the hydrolysis of the α-1,4 glycosidic linkages of N-acetyl-D-glycocyamine polymer of chitin randomly. It rapidly hydrolyzed swollen chitin and regenerated chitin but glycol chitin very slowly and can not hydrolyze N,N diacetyl chitobiose (Hirano et al., 1988; Leah et al., 1991). It is present in various seeds and vegetative tissues of higher plants, although there is no chitin, chitosan or chitin like substrate in higher plants (Boller et al., 1983; Hirano et al., 1988). Its activity also increased in germinated seeds (Lai et al., 1993). Also, chitinolytic activity in invertebrates (Kono et al., 1990), vertebrates (Han et al., 1997) and microorganism (Chernin et al., 1997; Porfir’eva et al., 1997) had been investigated.
Chitinases isolated from chick pea pods and leaves, wheat germ and bean leaves have maximum activities at 45, 50 and 37°C respectively (Molano et al., 1979; Boller et al., 1983; Nehra et al., 1994). The Ms of chitinases isolated from cucumber, barely grains, bean leaves and yam seeds were estimated to be 30×103, 33×103, 27×103 and 28×103 Dalton respectively (Boller et al., 1983; Tsukamoto et al., 1984; Jacobsen et al., 1990; Majeau et al., 1990). Chitinase is essential to bioconversion processes of waste shellfish chitin to N-acetylglucosamine (Carroad and Tom, 1978) and it may play a role in early embryo development (De Jong et al., 1992).
Several studies on the isolation and preparation of antifungal enzyme chitinase from microorganisms and plants have been investigated (Broekaert et al., 1988; Leah et al., 1991; Lorito et al., 1993). But the effect of antifungal enzymes from plants are found to be more potent in inhibition of growth of fungi than those from microorganisms (Roberts and Selitrennikoff, 1988; Park et al., 1992).
The present work has been directed towards the selection of suitable, available and inexpensive source for production of chitinase with high activity from a variety of Egyptian higher plant tissues. Purification and characterization of chitinase from leaves of sugar beet (Bete vulgaris L.) also will be carried out.
MATERIALS AND METHODS
Plant materials: Different parts of eleven plant families purchased from local market were used in this study (Table 1). Sugar beet leaves of fully grown plants were collected directly from Kafare El Shckh fields in march before harvesting sugar beet roots as time of collection must be controlled. Healthy seeds were germinated on filter paper moistened with sterile distilled water in dark or light chamber at 30°C for 10 days.
Extraction of chitinase: Dry seeds or germinated seeds or vegetative parts were washed with tap H2O, then with distilled H2O and homogenized. The resulting homogenate was extracted with cold distilled H2O. Insoluble materials were removed by centrifugation at 13×103 g (5°C) for 20 min dialyzed against H2O (5°C) for 2 days and the supernatant was used as crude enzyme preparation.
Purification of chitinase: The enzyme was partially purified by (NH4)2SO4 which was added to the crude enzyme preparation to yield 0-20, 20-60 and 60-100 percent saturation at 5°C. The resulting precipitates was centrifuged, dissolved in H2O then dialyzed against H2O (5°C) for 2 day. Insoluble materials were removed by centrifugation before enzyme assay. Enzyme activity and protein content were determined in each fraction.
Hydroxylapatite chromatography: The enzyme preparation from (NH4)2SO4 20-60 percent fraction was added to hydroxylapatite column and then eluted with sodium phosphate buffer (1.0 to 500 mM, pH 6.8).
Anion exchange chromatography: The prepared enzyme was loaded onto a DEAE-cellulose column previously equilibrated with sodium phosphate buffer (pH 7,0.01 M). After washing, the adsorbed proteins were eluted with a 0.1-0.5 M of NaCl in the same buffer.
Gel filtration orp Sephadex G-120 and G-200: The enzyme preparation from (NH4)2SO4 20-60 percent fraction were pooled and loaded onto a Sephadex G-120 column (0.9×32 cm) equilibrated with phosphate buffer (pH 7,0.01 M). The enzyme was eluted with the same buffer at a flow rate of 15 ml/hr. The final purification of the enzyme was performed by gel filtration on column of Sephadex G-200 (1.3×27 cm). The average of at least 5-6 times determination were carried out for all experiments.
Agarose gel electrophoresis: Confirmation of the purity of the enzyme was performed using agarose gel electrophoresis. It was carried out as described by Carlstrom and Johansson, (1983).
The purified enzyme was kept frozen at -20° before use in the experiments described below.
Substrate: Colloidal chitin was prepared from crab shell chitin according to the method described by Reid and Ogrydziak (1981). This method based on formation of colloidal chitin by treating chitin with 85 percent O-H3PO4 acid.
Enzyme assay: The chitinase activity was assayed calorimetrically. The reaction mixture contained an appreciable enzyme solution and 0.2 percent of colloidal chitin suspended in 0.1M acetate buffer, pH 4.5 in a total volume of 0.5 ml. After incubation at 40°C for 2 hr, N-acetyl glucose amine was determined with the p-dimethyl amino benzaldehyde reagent according to the method of Reissig et al. (1955). Enzyme activity was expressed as International milliunits (ImU). One ImU was defined as the production of 1 nmol of N-acetyl glucose amine per min at 40°C under the given conditions.
Activity determined from the linear portion of the time course was expressed as ImU per g dry tissues wt. and per mg protein. Each value is the mean of at least two to four replicate assays performed.
Protein determination: Protein concentration was measured according to the method of Lowry et al. (1951) using bovine serum albumin as standard. During chromatographic separation, it was estimated by absorbency at 260 and 280 nm (Warburg and Christian, 1941).
Effect of temperature on enzyme activity: The maximum activities of the tested enzyme was determined at different temperatures (25-60°C).
Effect of substrate concentration on enzyme activity: The effect of different substrate concentrations was tested by incubation different concentration of colloidal chitin with the same amount of chitinase enzyme. The enzymatic activities were plotted against substrate concentrations.
Effect of pH on enzyme activity: Small aliquot of the purified enzyme were assayed with three buffering systems, namely acetate (0.1M, pH 3.5-5.5), citrate-phosphate (0.1M, pH 2.5-6.5) and sodium phosphate (4.5-8.0).
Effect of reaction time: The purified enzyme was incubated with the colloidal chitin for different time intervals, then the reaction product was estimated and its amount was calculated as nmole N-acetyl glucose amine per reaction mixture.
Effect of enzyme concentration: The effect of enzyme concentration was tested by incubating different enzyme concentration (10-40 μ g) with the colloidal chitin at the optimum temperature. The relation between enzyme concentration and enzyme were plotted.
Determination of Mt: The Mt of the enzyme was estimated by gel filtration on a column of Sephadex G-200 (1.3×27cm). The column was calibrated with bovine serum albumin (69×103), egg albumin (44×103), myoglobulin (17.2×103) and lysozyme (14.2×103) as a standard proteins.
RESULTS
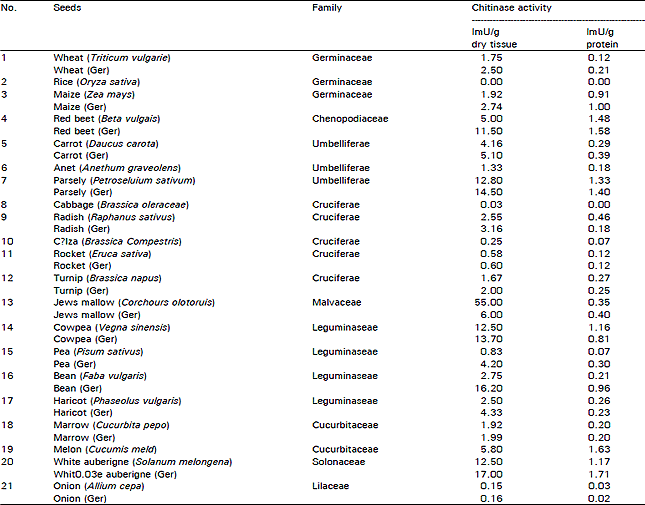
Chitinase level in some higher plants: The experiments have been done to choose the most suitable sources that give the highest activity for chitinase enzyme. Table 1 represents the results of screening for the existence of chitinase activity in different seeds either dry and after germination at 25-30°C for five days in dark. The results indicate that there are different variation in enzyme activities between the different seeds. Those showed high enzymatic activities were white aubergine, cowpea, melon, Jew’s mellow, carrot, red beet and bean.
Four seeds (bean, white aubergine, red beet and cowpea) were germinated for 10 days at 30°C and chitinase activity was daily determined (Fig. 1). It shows the optimum germination time for production of chitinase enzymes. The germinated dry seeds at 30°C in dark showed higher chitinase activity than in light. Chitinase activity of germinated white aubergine seeds increased continuously until the maximum activity was achieved at the 7th day with 2.2 fold increase as compared to the original dry seeds. But after the 7th day of germination the enzyme activity declined by 37 percent. The best day for production of chitinase from germinated bean and red beet seeds was the 5th day with 7.4 and 3.0 fold increase respectively and the activity was decreased by 76 percent at the 10th day. There was an increase in chitinase activity from germinated cowpea seeds until it reached the maximum at the 9th day with 1.2 fold increase then decreased by 33 percent at the 10th day.
Results in Table 2 show that the enzymes activities of the vegetative parts (leave, roots, stem and roots). Varied not only with the species of the plants, but also with the morphological parts of their tissues. The leaves of sugar beet, white aubergine, red beet cabbage, carrot and radish exhibited high activities.
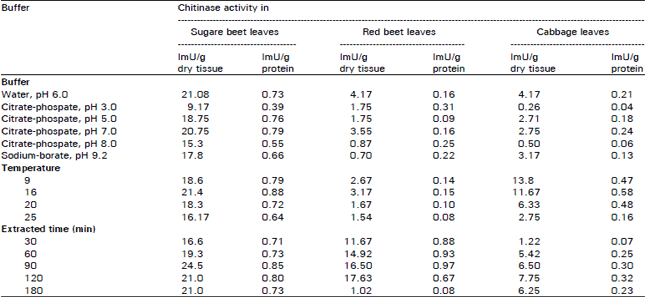
Assessment of optimum conditions for chitinase extraction from sugar beet, red beet and cabbage leaves: Results in Table 2 showed that leaves of sugar beet, cabbage and red beet gave high yield of chitinase. Concerning the optimum conditions for chitinase extraction from these leaves, Table 3 indicated that pH range was 5.0-7.0, temperature range was 9-16°C and time of extraction was 90-120 min.
As the results of the above screening, Egyptian sugar beet leaves were found to contain the largest amount of chitinase. Preliminary experiments also showed that frozen leaves can be stored for 9 months with negligible decrease in chitinase activity. It was chosen for preparing the pure enzyme.
Purification, of chitinase from sugar beet leaves
Protein fractionation by ammonium sulphate: The crude enzyme solution was extracted with distilled. H2O at optimum condition (extraction at 16°C for 90 min) from fresh healthy Egyptian sugar beet leaves. Precipitation of chitinase from the aqueous extract by (NH4)2SO4 (20-60%) showed a good recovery in their activities (75%), while saturation with 0-20 and 60-100 percent (NH4)2SO4 resulted in precipitate with low enzymatic activity and high protein content, therefore these precipitates were discarded.
Trials to purify the chitinase from (NH4)2SO4 fraction (20-60%) by hydroxylapatite and DEAE-cellulose technique showed no and low enzymatic activity respectively.
Gel filtration chromatography: Further purification of the chitinase from ((NH4)2SO4,20-60%) fraction by gel filtration on two successively columns of Sephadex G-120 (0.9×32 cm) and G-200 (1.3×27 cm) give a single peak of chitinase activity coincided exactly with protein peak (Table 4 and Fig. 2). The use of chromatography technique on Sephadex G-120 and G-2003 increased the specific activity of chitinase from 0.74 to 14.0 ImU/mg with 27.4 percent units recovery.
Seventy five mg protein (1050 ImU) of the purified chitinase was produced from 1 Kg fresh Egyptian sugar beet leaves (=173g dry). Lower amount of chitinase (4.7 mg protein per 1 Kg) was prepare from fresh thorn apple leaves than that prepared from sugar beet leaves (Broekaert et al., 1988).
Gel electrophoresis of the purified chitinase: The prepared enzyme was essentially homogenous as a single peak of protein was eluted from Sephadex G-200 column which coincided exactly with the peak of chitinase activity. The purified chitinase migrates as a Single band when applied to agarose gel electrophoresis.
| Table 1: | Determination of chitinase activity in aqueous extracte isolated from some seeds |
 | |
| Table 2: | Determination of chitinase activity in aqueous extracts isolated from some vegetative parts. |
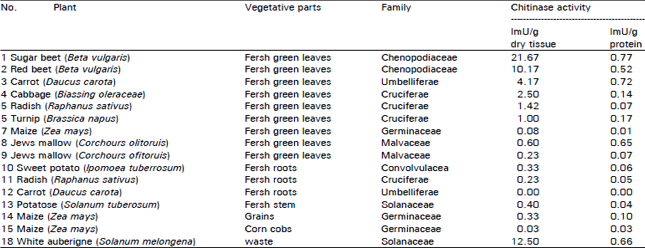 | |
| The average of at leat 3-4 determination of chitinase activity was calculated. One mU =1 nmol of N-acetylglucose amine per min at 40°C | |
| Table 3: | Effect of different pHs, temperature and time on extraction of chitinase from some leaves. |
 | |
| Table 4: | Purifcation scheme for chitinase from sugar beet leaves |
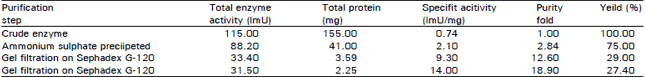 | |
| One ImU = 1 nmol of N-acetylglucose amine per min at 40°C. The average of at least 5-6 times of chitinase was calculated | |
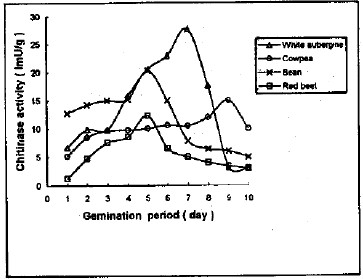 | |
| Fig. 1: | Determination of chitinase activity during germination days of some seeds |
Effect of different temperatures on chitinase activity: Chitinase activity was determined at different temp. within the range 25-60°C. The maximal activity for chitinase was found to be at 40-45°C with optimum at 40°C (Fig. 3). Sharp decline in chitinase activity was noticed at temp. 37 and 55°C with 63 and 68 percent decrease respectively.
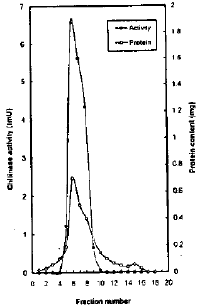 | |
| Fig. 2: | Chitinase activity and protein content obtained by filtration on Sephadex G-200 |
Effect of different pH’s on chitinase activity: The pH depended the enzymatic hydrolysis of colloidal chitin by the purified chitin was determined by replacing the buffer used in the respective standard enzyme assays with 0.1 M acetate buffer (for pH values between 3.5-5.5) or 0.1 M citrate phosphate buffer pH values between 2.5-6.5) or 0.1 M sodium phosphate buffer pH values between 4.5-8.0).
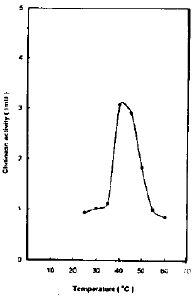 | |
| Fig. 3: | The effect of different temperature on chitinase activity |
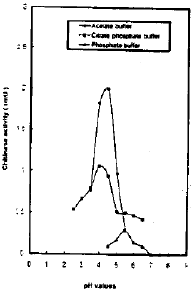 | |
| Fig. 4: | The effect of pH’s on chitinase activity |
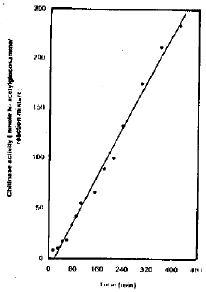 | |
| Fig. 5: | The effect of reaction time on chitinase activity |
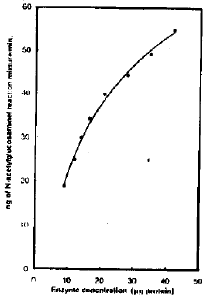 | |
| Fig. 6: | The effect of different concentrations of chitinase on its activity |
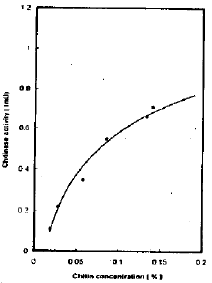 | |
| Fig. 7: | The effect of different colloidal chitin concentration on chitinase activity |
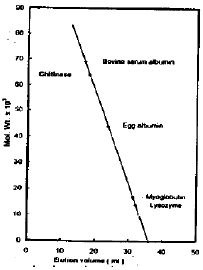 | |
| Fig. 8: | Determination of molecular weight of chitinase by gel filtration on Sephadex G-200 column |
Chitinase activity exhibited maximum activity at pH 4.0-4.5, 5.5 and 4.0-4.5 using 0.1 M acetate, 0.1 M phosphate and 0.1 M citrate phosphate buffers respectively (Fig. 4). The best buffer used having the maximal activity was 0.1M acetate buffer at pH 4.5. This maximal activity decreased to half at pH 5.0 and more than 83 percent at pH 5.5.
Effect of reaction time an chitinase activity: The activities of chitinase were linear function of incubation time up to 420 min. using colloidal chitin as substrate (Fig. 5). It could be concluded that excess products with increasing time of reaction may not interfere with the enzymes activities. This property could be considered as suitable one for using these enzymes in the field of medicine for long time.
Effect of different concentrations of chitinase on its activity: The enzymatic activity of chitinase was increased with increasing in enzyme concentration up to 40 μ g protein per reaction mixture (Fig. 6).
Effect of substrate concentrations on chitinase activity: There is a direct relation between colloidal chitin concentration and the chitinase activity up to 0.11 percent per reaction mixture (Fig. 7).
Determination of molecular weight: The Mt of the prepared chitinase was determined to be 64×103 Dalton (Fig. 8).
DISCUSSION
Chitinase is widespread in higher plants. In this study, screening experiments on different plant parts representing eleven families including seeds (dry and germinated) and vegetative parts were carried out tp determined the mot suitable source for chitinase activity. Result of the screening experiments indicated that the enzymatic activity was varied not only with the species of the tested plants, but also according to the morphology of the examined parts. These results are to great extent in accordance to those found by Powning and lrzykiewicz (1965), Abeles et al. (1971), Hirano et al. (1990), Leah et al. (1991) and Chang et al. (1996). The increase in activity of chitinase during germination may be due to activation of the latent form and for increase in the rate of de novo synthesis of enzymes. These results are consistent with other works which indicated that chitinase enzyme increased greatly during germination of some Japanese seeds (Hirano et al., 1988; Hirano et al., 1990). However, cucumber chitinase is only presented in the mature seeds or in rootlet, but during early stages of germination a very low level was detected (Majeau et al., 1990).
The differences in optimum conditions for extraction of chitinase from sugar beet, red beet and cabbage leaves may be due to relative solubility and stability of each enzyme. Abeles et al. (1971) used distilled H2O as an extraction solution for isolation of chitinase from bean leaves, while Fink et al. (1988) used sodium phosphate buffer (pH 6.0) to isolate this enzyme from oat leaves. In this study, sugar beet leaves proved to be the best source for chitinase. As it is an agridulture waste and a source of pollution, trials were done to use it in production of beneficial enzyme and consequently aid in consuming the pollution.
Pure chitinase was isolated from sugar beet leaves with relatively high recovery of activity ( 27.4%) as compared to that of soybean seeds and cabbage stem (12%) and (18%) isolated by Wadsworth and Zikakis (1984) and Chang et al. (1996) respectively, but lower than those from thorn apple (48%) (Broekaert et al., 1988).
The optimum pH for chitinase activity from sugar beet was measured to be 4.5 which are in accordance to those reported by Wadsworth and Zikakis (1984), Kono et al. (1990) and Zhang et al., (1996) for soybean seeds Japanese eel and carrot roots chitinase respectively. While Molano et al. (1979) and Chang et al. (1996) isolated chitinase from wheat grain and cabbage stem with an optimum pH 6.0. Chitinase isolated from Bacillus cereus showed broad optimum pH from 4.0 to 10.0 (Trachuk et al., 1996).
The optimum temperature for chitinase activity was found to be 40°C which is quite similar to that of the enzyme isolated from tomato leaves and chick pea leaves and pods (Joosten and de wit 1989; Nehra et al., 1994). It is higher than optimum temp. of wheat grain and carrot roots chitinase (30 and 25°C) isolated by Molano et al. (1979) and Zhang et al. (1996) and lower than optimum temp. of cabbage stem and Bacillus cereus chitinase (60°C) isolated by Chang et al. (1996) and Trachuk et al. (1996).
The molecular weight of the purified chitinase was determined to be 64×103. Concerning the M,s of chitinase enzyme isolated from different sources, it was found to be different, as chitinase isolated from bean leaves, soybean seeds, cabbage stem with root and tomato stem tissue have M,s range from 27×103 to 31×101 Dalton (Pegg and Young, 1982; Wadsworth and Zikakis, 1984; Chang et al., 1996). It was found that the M,s of stomach of Japanese eel, Bacillus, cereus, Serratia marcescens and S. peculiarities chitinase were 59×103, 68.52×103, 58×103 and 45×103 Dalton respectively (Kono et al., 1990; Zhang et al., 1996: Porfir’eva et al., 1997).
In conclusion, the screening tests indicated that different parts al plants are rich in chitinase activity specially leaves of sugar beet, cabbage and beet root. Sugar beet leaves were found to be the most convenient inexpensive source for enzyme production and very economic source. Activity of chitinase in the fresh sugar beet leaves remain stable when keeping these leaves at -20°C for months. The purification procedure described here is simple, inexpensive and give an industrially useful enzyme with god specific activity at 40°C in 0.1 M acetate buffer with pH 4.5.
REFERENCES
- Abeles, F.B., R.P. Bosshart, L.E. Forrence and W.H. Habig, 1971. Preparation and purification of glucanase and chitinase from bean leaves. Plant Physiol., 47: 129-134.
Direct Link - Boller, T., A. Gehri, F. Mauch and U. Vogeli, 1983. Chitinase in bean leaves: Induction by ethylene, purification, properties and possible function. Planta, 157: 22-31.
CrossRefDirect Link - Broekaert, W.F., J. van Parijs, A.K. Allen and W.J. Peumans, 1988. Comparison of some molecular, enzymatic and antifungal properties of chitinases from thorn-apple, tobacco and wheat. Physiol. Mol. Plant Pathol., 33: 319-331.
CrossRefDirect Link - Carlstrom, A. and B.G. Johansson, 1983. Agarose gel electrophoresis-immunofixation. Scand. J. Immunol., 17: 23-32.
CrossRefDirect Link - Carroad, P.A. and R.A. Tom, 1978. Bioconversion of shellfish chitin wastes: Process conception and selection of microorganisms. J. Food Sci., 43: 1158-1161.
CrossRefDirect Link - Chang, C.T., Y.L. Hsueh and H.Y. Sung, 1996. Purification and properties of chitinase from cabbage stems with roots. IUBMB Life, 40: 417-425.
CrossRefDirect Link - Chernin, L., A. Gafni, A. Sztejnberg, R. Mozes-Koch and U. Gerson, 1997. Chitinolytic activity of the acaropathogenic fungi Hirsutella thompsonii and Hirsutella necatrix. Can. J. Microbiol., 43: 440-446.
CrossRefDirect Link - De Jong, A.J., J. Cordewener, F.L. Schiavo, M. Terzi, J. Vandekerckhove, A. van Kammen and S.C. de Vries, 1992. A carrot somatic embryo mutant is rescued by chitinase. Plant Cell Online, 4: 425-433.
Direct Link - Fink, W., M. Liefland and K. Mendgen, 1988. Chitinases and β-1,3-glucanases in the apoplastic compartment of oat leaves (Avena sativa L.). Plant Physiol., 88: 270-275.
Direct Link - Han, B.K., W.J. Lee and D.H. Jo, 1997. Survey on the chitinolytic activity from some animals for the industrial utilization. J. Korean Soc. Applied Biol. Chem., 40: 95-100.
Direct Link - Hirano, S., M. Hayashi, K. Murae, H. Tsuchida and T. Nishida, 1988. Chitosan and Derivatives as Activators of Plant Cells in Tissues and Seeds. In: Applied Bioactive Polymeric Materials, Gebelein, C.G., C.E.Carraler Jr. and V.R. Foster (Eds.). Plenum Publishing Co., New York, ISBN: 978-1-4684-5612-7, pp: 45-59.
- Hirano, S., T. Yamamoto, M. Hayashi, T. Nishida and H. lnui, 1990. Chitinase activity in seeds coated with chitosan derivatives. Agric. Biol. Chem., 54: 2719-2720.
CrossRefDirect Link - Jacobsen, S., J.D. Mikkelsen and J. Hejgaard, 1990. Characterization of two antifungal endochitinases from barley grain. Physiol. Plant., 79: 554-562.
CrossRefDirect Link - Joosten, M.H.A.J. and P.J.G.M. de Wit, 1989. Identification of several pathogenesis-related proteins in tomato leaves inoculated with Cladosporium fulvum (syn. Fulvia fulva) as 1,3-β-glucanases and chitinases. Plant Physiol., 89: 945-951.
Direct Link - Kono, M., T. Matsui, C. Shimizu and D. Koga, 1990. Purification and some properties of chitinase from the stomach of Japanese eel, Anguilla japonica. Agric. Biol. Chem., 54: 973-978.
CrossRefDirect Link - Lai, D.M., P.B. Hoj and G.B. Fincher, 1993. Purification and characterization of (1→ 3, 1→ 4)-β-glucan endohydrolases from germinated wheat (Triticum aestivum). Plant Mol. Biol., 22: 847-859.
CrossRefDirect Link - Leah, R., H. Tommerup, I.B. Svendsen and J. Mundy, 1991. Biochemical and molecular characterization of three barley seed proteins with antifungal properties. J. Biol. Chem., 266: 1564-1573.
Direct Link - Lorito, M., A. di Pietro, C.K. Hayes, S.L. Woo and G.E. Harman, 1993. Antifungal, synergistic interaction between chitinolytic enzymes from Trichoderma harzianum and Enterobacter cloacae. Phytopathology, 83: 721-728.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Majeau, N., J. Trudel and A. Asselin, 1990. Diversity of cucumber chitinase isoforms and characterization of one seed basic chitinase with lysozyme activity. Plant Sci., 68: 9-16.
CrossRefDirect Link - Molano, J., I. Polacheck, A. Duran and E. Cabib, 1979. An endochitinase from wheat germ. Activity on nascent and preformed chitin. J. Biol. Chem., 254: 4901-4907.
PubMedDirect Link - Nehra, K.S., L.K. Chugh, S. Dhillon and R. Singh, 1994. Induction, purification and characterization of chitinases from chickpea (Cicer arietinum L.) leaves and pods infected with Ascochyta rabiei. J. Plant Physiol., 144: 7-11.
CrossRefDirect Link - Pegg, G.F. and D.H. Young, 1982. Purification and characterization of chitinase enzymes from healthy and Verticillium albo-atrum-infected tomato plants and from V. albo-atrum. Physiol. Plant Path., 21: 389-409.
CrossRefDirect Link - Powning, R.F. and H. Irzykiewicz, 1965. Studies on the chitinase system in bean and other seeds. Comp. Biochem. Physiol., 14: 127-133.
CrossRefDirect Link - Porfir'eva, O.V., D.V. Yusupova, N.L. Zotkina, R.B. Sokolova and L.A. Gabdrakhmanova, 1997. Chitinolytic complex of Serratia marcescens and peculiarities of its biosynthesis. Microbiology, 66: 289-294.
Direct Link - Reid, J.D. and D.M. Ogrydziak, 1981. Chitinase-overproducing mutant of Serratia marcescens. Applied Environ. Microbiol., 41: 664-669.
Direct Link - Reissig, J.L., J.L. Strominger and L.P. Lelor, 1955. A modified calorimetric method for the estimation of N-acetylamino sugar. J. Biol. Chem., 217: 959-966.
PubMed - Roberts, W.K. and C.P. Selitrennikoff, 1988. Plant and bacterial chitinases differ in antifungal activity. J. Gen. Microbiol., 134: 169-176.
CrossRefDirect Link - Tsukamoto, T., D. Koga, A. Ide, T. Ishibashi, H. Horino-Matsushige, K. Yagishita and T. Imoto, 1984. Purification and some properties of chitinases from yam, Dioscorea opposita THUMB. Agric. Biol. Chem., 48: 931-939.
Direct Link - Wadsworth, S.A. and J.P. Zikakis, 1984. Chitinase from soybean seeds: Purification and some properties of the enzyme system. J. Agric. Food Chem., 32: 1284-1288.
CrossRefDirect Link - Zhang, Y.Y., N.H. Haunerland and Z.K. Punja, 1996. Chitinase profiles in mature carrot (Daucus carota) roots and purification and characterization of a novel isoform. Physiol. Planta, 96: 130-138.
CrossRefDirect Link