
Abdulaziz M. Al-Bahrany
Department of Horticulture, King Faisal University, P.O. Box 420, Al-Hassa 31982, Saudi Arabia
Jameel M. Al-Khayri
Date Palm Research Center, College of Agriculture and Food Sciences,King Faisal University, P.O. Box 420, Al-Hassa 31982, Saudi Arabia
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 10 | Page No.: 1778-1780







ABSTRACT
Cottonseeds (Gossypium hirsutum L.) from lines 54027 and 84033, characterized by enhanced salt tolerance, and their parental line DP62 were studied qualitatively and quantitatively to determine fatty acid composition and proximate analysis. Proportions in fatty acid composition were found to be modified as a result of the salt selection-breeding program. Linoleic acid (18:2) was higher in both salt tolerant lines while palmitic acid (16:0) was lower in line 84027. Significantly different unsaturatedisaturated fatty acid ratios, 3.13, 3.20, and 3.59 were obtained from genotype DP62, 84033 and 84027, respectively. Other parameters studied also reflected genotypic variability, including crude protein, crude oil, crude fiber, ash, nitrogen free extract, and carbohydrates content, This research may contribute to establishing a relationship between the unsaturated/saturated fatty acid ratio of a particular genotype and its ability to maintain physiological and biochemical functions under salt stress.
PDF Abstract XML References Citation
How to cite this article
Abdulaziz M. Al-Bahrany and Jameel M. Al-Khayri, 2000. Genotype Variability in Fatty Acid Composition and Chemical Characteristics of Cotton (Gossypium hirsutum L.). Pakistan Journal of Biological Sciences, 3: 1778-1780.
DOI: 10.3923/pjbs.2000.1778.1780
URL: https://scialert.net/abstract/?doi=pjbs.2000.1778.1780
DOI: 10.3923/pjbs.2000.1778.1780
URL: https://scialert.net/abstract/?doi=pjbs.2000.1778.1780
INTRODUCTION
Mature cotton (Gossypium hirsutum L.) seeds contain oils and proteins at differing concentrations depending on genotype (Cherry et al., 1878a,b, 1981a,b; Lawhon et al., 1976). These compounds are metabolized to provide energy for embryo germination. The content of such compounds and other biochemical and physiological parameters have been shown to differ among salt tolerant plants (Bowman et al., 1996; Caldwell, 1974; Epistein, 1972). Genetic modifications by breeding and agronomic practices can affect both the physical and chemical properties of cottonseed (Cherry and Leffler, 1984); for example, improvement of oil and fatty acids composition (Cherry, 1983).
Salt-tolerant cotton gerrnplasm lines (84027 and 84033) have been developed from the parental line Deltpine 62 (DP62) through a breeding program carried out by Ledbetter (1986). During germination and emergence under varying NaCI conditions, lines 84027 and 84033 showed superior vigor and higher ribosomal-RNA content in relation to line DP62. Furthermore, the selection process for salt tolerance influenced seed germination and growth rate (Al-Bahrany. 1989). The objective of this study was to determine differences elicited by salt selection breeding program between salt tolerant cotton lines 84027 arid 84033 and parental line DP62 in relation 10: 1) Seed composition of fatty acids and unsaturated/saturated ratios and 2) Proximate analysis for moisture, crude protein, crude oil, crude fiber and ash content.
MATERIALS AND METHODS
Plant material: Three cotton germplasms used in this study including two lines selected for salt tolerance (lines 84027 and 84033) and their parental cultivar Deltpine 62 (DP62) eines 84027 and 84033 represent the second selection cycle for germination salt tolerance.
Fatty acids composition: The methyl esters of the fatty acids were prepared directly from seeds and fatty acids content was determined using gas chromatography as described by Flood (1981) with modifications reported by El-Shintinawy and Selim (1995). Grounded clean and dry seeds, 200 mg, were mixed with 4 ml of 0.66 N KOH in methanol then placed in a 20 ml screw capped tube. The tube was flushed with N2, sealed and heated at 100°C for 5 min. The mixture was cooled and 4 ml of boron-trifluoride (BF3) in methanol (14% wlv, BDH Co.) was added. The tube was flushed with N2, capped and heated at 100°C for 5 minutes. The mixture was cooled and 4 ml of each of saturated NaCI; water and petroleum ether (b.p. 40-60°C) were added. The supernatant was removed, washed with water, dried over anhydrous N2SO4, and concentrated by stream of N2 at room temperature to about 1 ml. Methyl ester of dodecanoic acid (2 was added as internal standard. A sample of 0.5 μl dried supernatant was injected into a GLCvariant 6000 with Flame Ionization Detector (HD), 2 m length, 18 inch internal diameter stainless steel column, packed with 15% OV-275. Chrom Placid wash/80-100 mesh stationary phase operated at 175°C. Injection temperature was 230°C. Detector temperature 2513°C with helium carrier gas at a flow rate of 25 ml/min., hydrogen flow 30 ml/min, air flow 300 ml/min. Identification of fatty acid methyl esters was carried out by comparison of their retention times with that of the standard and the quantities were calculated front the areas obtained by the KLB-2220 Recorder Integrator. The unsaturated/saturated fatty acid ratio of each sample was obtained by dividing the summed percentage of unsaturated fatty acids by the summed percentage of saturated fatty acids.
Proximate analysis: Proximate analysis for moisture, crude protein, crude oil, crude fiber, and ash were performed in accordance to the methods described in the Association of Official Analytical Chemist (AOAC, 1984). A sample weighing 5 g of seeds was used for analysis. Total carbohydrates were estimated by subtracting the sum of moisture, crude protein, crude oil, and ash content percentages from one hundred. Nitrogen-free extract (NFE content) was determined by subtracting crude fiber percentage from total carbohydrates percentage.
Statistical analysis: Three samples per genotype were analyzed, and data were subjected to an analysis of variance end means were separated using least significance difference (LSD), where appropriate, at 5% significance level.
RESULTS AND DISCUSSION
Fatty acid composition: The salt selection process was associated with decreasing total saturated fatty acids (myristio, palmitic and stearic) and increasing total unsaturated fatty acids (palmitoleic, oleic and linoleic) (Fig. 1). The principle unsaturated fatty acid of cottonseeds oil in the genotypes tested were linoleic (18:2) followed by oleic (18:1). Palmitic acid (16:0) was the most abundant Saturated fatty acid (Table 1).
| Table 1: | Mean fatty acid compositions and unsaturated/saturated fatty acid ratios from hulled cottonseed of genotypes Deltpine 62, line 84027, and line 84033 |
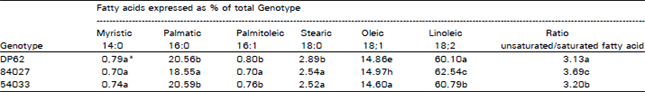 | |
| *Values followed by the same letter in each column are not significantly different at 0.05 level by LSD | |
| Table 2: | Proximate analysis of cottonseed of genotypes Deltpine 62, line 84027, and line 84033 |
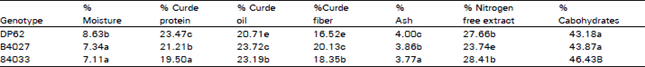 | |
| *Values followed by the same letter in each column are not significantly different at 0.05 level by LSD | |
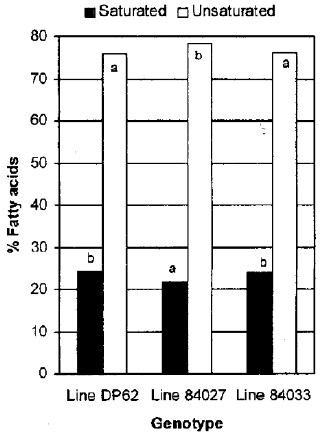 | |
| Fig. 1: | Total saturated and total unsaturated fatty acids exhibited by cotton genotypes Deltpine 62, line 84027, and line 84033 |
These results in agreement with those obtained by other researchers studying various genotypes (Cherry et al., 1981a; Sawan et al., 1989).
Except for myristic acid (14:0), fatty acid composition differed significantly among genotypes (Table 1). Line 84027 had the highest significant concentration of linoleic acid 162.54%) of all genotypes analyzed followed by line 84033, whereas the parental line DP62 lowest concentration of linialeic acid 160.10%) (Table 1). Palmitic acid was lower in line 84027 than either of the other two genotypes. Stearic acid was significantly lower in the salt selected lines in comparison to the parental line. Unsaturated/saturated fatty acid ratios differed according to genotype. Lines 84027 and 84033, characterized by salt tolerance, exhibited higher unsaturated/saturated fatty acid ratio than the parental line (DP62). However, line 84027 had significantly higher ratio (3.59) than line 84033 (3.20) (Table 1). The genotypes that were associated with high unsaturated/saturated fatty acids ratios (lines 84027 and 84033) were previously shown to have greater emergence percentage under salt stress and higher ribosomal-RNA than DP62 (Al-Bahrany, 1989). This suggests that there might be a relationship between unsaturated/saturated fatty acid ratios and salt tolerance. It can be noted that the salt selection process could have resulted in genetic modifications, thus affecting the ratio and composition of unsaturated and saturated fats, despite of the common origin of the two lines.
Proximate analysis: Apparently the selection probess for seed germination salt tolerance resulted in measurable genetic changes and altered chemical composition in all parameters tested (Table 2). The parental line DP62 had a higher protein and ash percentages and lower oil and carbohydrates percentages than the other salt tolerance genotypes (84027 and 84033). The protein content ranged from 19.50 to 23.47%. Protein content in cottonseed of other genotypes ranged from 19.6 to 24.0% (Lawhor et al., 1976) and 25.6 to 27.6% (Cherry et al., 1978a,b, Cherry et al., 1981a,b).
In our study, seed oil content ranged from 20.71 to 23,72%. Salt tolerant line 84027 had the highest increase 13.01 %lin oil content followed by line 84033 (2.48%) in relation to the parental line DP62. In comparison, others have shown that el; content ranged from 18.5 to 25.6% (Lawhon et al., 1976) and 23.2 to 25.7% (Cherry et al. 1978a,b, 1981a,b).
In conclusion, selection procedures for salt tolerance were effective in altering chemical composition in cottonseed. Salt tolerant lines exhibited higher percentage of unsaturated/saturated fatty acid ratios than parental line. This ratio, however, differed between the two salt tolerant lines. Based upon these observations, it can be assumed that fatty acid composition of the two salt tolerant lines were genetical!y modified in different ways by salt breeding program, even though they were selected from a single variety.
REFERENCES
- Bowman, D.T., O.L. May and D.S. Calhoun, 1996. Genetic base of upland cotton cultivars released between 1970 and 1990. Crop Sci., 36: 577-581.
CrossRefDirect Link - Cherry, J.P., J.G. Simmons and R.J. Kohel, 1978. Cottonseed Composition of National Variety Test Cultivars Grown at Different Texas Locations. In: Proceedings of the Beltwide Cotton Production Research Conference, Brown, J.M. (Ed.). National Cotton Council of America, Memphis, Tenn., USA., pp: 47-50.
- Cherry, J.P., M.S. Gray and L.A. Jones, 1981. A review of lecithin chemistry and glandless cottonseed as a potential commercial source. J. Am. Oil Chem. Soc., 58: 903-913.
CrossRefDirect Link - Lawhon, J.T., C.M. Cater and K.F. Mattil, 1976. Evaluation of the food use potential of sixteen varieties of cottonseed. J. Am. Oil Chem. Soc., 54: 75-80.
CrossRefDirect Link - Sawan, Z.M., R.A. Sakr and F.A. Ahmed, 1989. Effect of 1-naphthaleneacetic acid on the seed, protein, oil and fatty acids of Egyptian cotton. J. Am. Oil Chem. Soc., 66: 1472-1474.
CrossRefDirect Link