
Leong Sue Yie
Fermentation and Enzyme Technology Laboratory, School of Biological Sciences, University Sains Malaysia, Minden, 11800, Penang, Malaysia
Ibrahim Che Omar
Fermentation and Enzyme Technology Laboratory, School of Biological Sciences, University Sains Malaysia, Minden, 11800, Penang, Malaysia
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 10 | Page No.: 1752-1754







ABSTRACT
Optimization of the solid state fermentation on PKC for benzyldehyde production by a locally isolated fungus, which was identified to be Penicillium diversum revealed that at the temperature of 30̊C, particle size of PKC of 1 mm, water content of 40%, glucose to ammonium sulphate supplementation ratio of 1:1 and inoculum size of 105 spores g–1 PKC, benzyldehyde production of 7.2 mg g–1 PKC was obtained after 5 days fermentation. This is an increase by more than 80% compared to before optimization, The production does not depend on the growth of the fungus.
PDF Abstract XML References Citation
How to cite this article
Leong Sue Yie and Ibrahim Che Omar, 2000. Optimization of a Solid State Fermentation for Benzyldehyde Production by a Locally Isolated, Penicillium diversum Using Palm Kernel Cake (PKC) as Substrate. Pakistan Journal of Biological Sciences, 3: 1752-1754.
DOI: 10.3923/pjbs.2000.1752.1754
URL: https://scialert.net/abstract/?doi=pjbs.2000.1752.1754
DOI: 10.3923/pjbs.2000.1752.1754
URL: https://scialert.net/abstract/?doi=pjbs.2000.1752.1754
INTRODUCTION
The preference by consumers for products from natural resources has initiated work on the use of microorganisms as sources of flavouring compounds. One of these compounds is benzyldehyde, an aromatic aldehyde which is mainly used as flavour compounds to give the cherry and almond flavour. It is the second most important compound in flavour and fragrance industries after vanillin. Natural Benzyldehyde used in food industries is normally extracted from almonds, kernel of apricots, peaches and plums. These natural benzyldehyde extracted from fruits contained undesirable toxic by-products such as hydrocyanic acid. Benzyldehyde can also be synthesised chemically, however the process was found to be less environmental friendly as it requires heavy metals as catalyst, high energy cost and the by-products contain undesirable racemic mixtures (Lawrence and Yuan, 1996). Therefore, the production of natural benzyldehyde through fermentation processes was considered. Production of benzyldehyde by submerged culture systems using various microorganisms have been reported using Lactobacillus plantarum (Groot and De Bont, 1998), Pseuclomonas putida (Simmonds and Robinson, 1998) and Polyporus tuberaster (Kawabe and Morita, 1994). However, solid state fermentation (SSF) process for benzyldehyde production has not been reported. We have consider the SSF process for the main reason of its simplicity, low energy cost and at the same time with the objective to utilise palm kernel cake (PKC) which was used as the substrate. PKC is a solid waste material obtained from the processing of palm oil and the production of PKC in Malaysia escalates rapidly since the eighties (Hutagalung et al., 1982) with the current production to be more than a million ton per year. Currrently, PKC is exported to European countries which are used as animal feeds due to its high protein content and good amino acid profiles. In this study, the culture conditions that affect the production of benzyldehyde in SSF system by a locally isolated fungus were investigated. The fungal isolates for the selection of potential benzyldehyde producers were obtained either from soil samples, rotting fruits, or local fermented foods. The ability for the isolates to produce benzyldehyde on PKC in a SSE system was determined using the fermentation medium consisting of 10 g of PKC of 2.0 mm particle size in a 150 ml conical flask. The substrate was inoculated with 5×105 spores suspended in 5 ml of distilled which was added into the substrate. Incubation was carried out at 30̊C and after 5 days, benzyldehyde production and fungal growth were analysed. This fermentation medium was also used as the basal medium in the optimization experiments. Benzyldehyde that was secreted extracellularly was extracted by adding 10 g of the substrate in 25 ml of 0.2 M acetate buffer containing 0.1% Tween 80. The mixture was vigorously homogenised for 10 min, followed by filtration through Muslin cloth. The filtrate collected was centrifuged for 20 min at 5000 rpm. The supernatant collected after centrifugation was diluted accordingly using distilled water and was used as the source of benzyldehyde. The concentration of benzyldehyde expressed as mg benzyldehyde produced g–1 PKC was determined spectrophotometrically at 245 rim (Lawrence and Yuan, 1996) against a standard curve obtained from the commercially available benzyldehyde (Sigma).
The growth of the fungus was determined by the method described by Swift (1973) based on the spectrophotometric measurement of glucosannine at 530 nm. The amount of glucosamine released from the fungus which was expressed as mg g–1 PKC was determined based on the standard curve of glucosamine (Sigma).
From a total of 150 fungal isolates screened, isolate K5 was selected, with the production of benzyldehyde of 4.0 mg g–1 PKC and growth of 1.0 mg glucosamine g–1 PKC. Based on the morphological characteristics observed under the Scanning Electron Microscope, KS was identified as Penicillium diversum. The fungus posseses 5 symmetrical phialides with 6 metula (Fig. 1). The conidiophores are smooth and the conidia are ellipsoidal. Other benzyldehyde producing fungi that were reported include Bjerkandera adusta, Ischnoderma benzoinum, Dichomitus squalens (Lapadatescu et al., 1997) and Trichosporon beigelli (Casey and Dobb, 1992).
The effect of particle size of PKC from 0.5-2.5 mm on the production of benzyldehyde and growth of Penicillium diversum is shown in Fig. 2. It was observed that the particle size of 1.0 mm resulted the highest benzyldehyde production of about 4.4 mg g–1 PKC after a 5 day cultivation with the growth of 1.8 mg glucosamine g PKC.
 | |
| Fig. 1: | Penicillium diversum showing (A) the conditiophore bearing metula and phialides (meg. 2800 X) and the conidia attached to the metula, (B) ellipsoidal conidia (meg. 5000 X) |
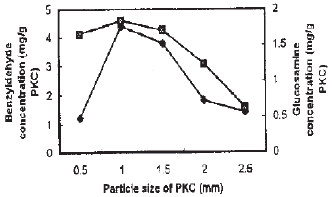 | |
| Fig. 2: | Effect of PKC particle size on benzyldehyde production and growth of penicillium diversum symbols: benzyldehyde concentration (♦) and growth (■) |
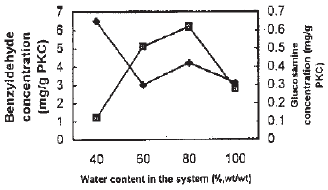 | |
| Fig. 3: | Effect of the ratio of water contcent of PKIC on benzyldehyde producation and growth of symbols: see Fig. 2 |
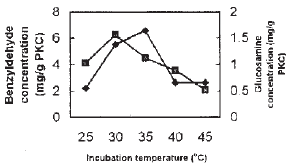 | |
| Fig. 4: | Effect of temperature on benzyldehyde production and growth of Penicillium diversum Symbols: see Fig. 2 |
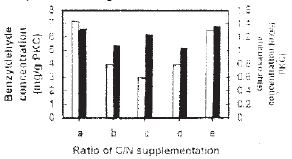 | |
| Fig. 5: | Effect of glucose and ammonium sulphate supplementation ratio on benzyldehyde production and growth of Penicillium diversum The supplementation of glucose and ammonium sulphate was carried out as follows (%) a: 0.25:0.25; b: 0.375:0.125; c: 0.125:0.375; d:0.5:0; e: 0:0.5. Symbols: benzyldehyde concentration (□) and growth (■) |
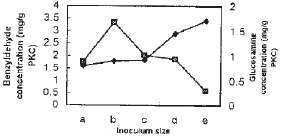 | |
| Fig. 6: | Effect of inoculum size in SSF system on benzyldehyde producation and growth of penicillium diversum The inoculum sizes examined were (number of spores g–1 PKC); a: 104; b: 105; c: 105; d: 107; e: 108 Symbols: see Fig. 2 |
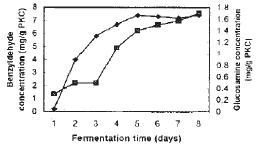 | |
| Fig. 7: | Time course of benzyldehyde producation and growth of penicillium diversum under the optimized culture conditions of the SSF system. Symbols: see Fig. 2 |
Large particle size of PKC of more than 1.0 mm does not improve either the benzyldehyde production or the growth, which may be related to poor accessibility of nutrient from the substrate to the fungus.
Considering the ratio of 1:1 (weight: weight) of water added to the substrate as 100% water content, the effect of water content was determined from 40-80%. As shown in Fig. 3, it was observed that the optimum water content was 40% with the production of benzyldehyde of 6.5 mg g–1 PKC, although the growth of the fungus dropped drastically to only about 0.6 mg glucosamine per g substrate. Higher water content of more than 40% although resulted in good fungal growth, does not seem to improve the productivity of the benzyldehyde.
The optimum temperature for the growth of P. diversum is 30̊C with the growth ranging from 1.5-1.8 mg giucosamine g–1 PKC. However, the optimum temperature for the production of benzyldehyde was observed at 35̊C with the benzyldehyde production of 6.6 mg g–1 PKC (Fig. 4). However, at 35̊C, the growth of the fungus dropped to about 1.1 mg glucosamine g–1 PKC. Higher temperatures were found unfavourable for benzyldehyde production and growth of P. diversum.
Apart from PKC, glucose and ammonium sulphate were also supplemented in the substrate as the additional carbon and nitrogen sources, respectively. The total concentration of glucose and ammonium sulphate added was 0.5% (w/w) and the effect of the ratio between C and N sources were determined at 1:1 (0.25:0.25%), 1:3 (0.125:0.375%) and 3:1 (0.375:0.125%). The addition of either glucose and ammonium sulphate at 0.5% respectively, was also determined. The CN ratio of 1:1 gave the highest production of benzyldehyde of 7.1 mg g–1 PKC, while the growth was 1.4 mg glucosamine g–1 PKC (Fig. 5). An almost similar results was obtained when only 0.5% of ammonium sulphate was added in the substrate. On the other hand, growth was significantly enhanced when only 0,5% of glucose was added in the substrate although benzyldehyde production was reduced to about 5.8 mg g–1 PKC. The result indicated that the fungus was able to utilize high glucose concentration that would be inhibitory in submerged cultures.
Based on the results obtained so far, it is clear that benzyldehyde production is growth independent. The results obtained on the effect of inoculum size again demonstrated that the increase in inoculum size increases the growth, although benzyldehyde production decreases (Fig. 6). The inoculum size in the range of 105-107 spores g PKC resulted in the production of about 7.2 mg g–1 PKC of benzyldehyde.
Based on the cultural conditions of SSF after optimization, consisting of particle size of PKC of 1.0 mm, incubation temperature of 30̊C, water content of 40% (w/w), glucose to ammonium sulphate supplementation ratio of 1:1 and inoculum size of 1×105 spores g–1 PKC, P. diversum was able to produce about 7.2 mg g–1 PKC of benzyldehyde with a growth of 1.4 mg glucosamine g PKC after 5 days (Fig. 7). Comparative studies on benzyldehyde productivity is not quite possible due to the differences in the culture system. However, the production by P. diversum in a SSF system can be considered significant based on the production by submerged systems reported so far ranged from 7-10 mg L–1 (Casey and Dobb, 1992; Kawabe and Morita, 1994, Simmonds and Robinson, 1998). When compared to the conditions before optimization, the benzyldehyde production increased by about 80%, while the growth was improved by about 43%. The results suggested that PKC can be a potential substrate for the production of microbial metabolites through the SSF system. These optimized parameters have been translated for mass production using the deep bed SSF system which was designed and fabricated in the laboratory. The results on the mass production of benzyldehyde by P. diversum will be reported subsequently.
ACKNOWLEDGEMENT
The Research Grant from, National Biotechnology Directorate (NBD), Ministry of Science, Technology and Environment, Malaysia under The Top-Down Biotechnology Project (280/6107/0118) is highly appreciated.
REFERENCES
- Casey, J. and R. Dobb, 1992. Microbial routes to aromatic aldehydes. Enzyme Microbial Technol., 14: 739-747.
CrossRefDirect Link - Kawabe, T. and H. Morita, 1994. Production of benzaldehyde and benzyl alcohol by the mushroom Polyporus tuberaster K2606. J. Agric. Food Chem., 42: 2556-2560.
Direct Link - Lapadatescu, C., G. Feron, C. Vergoignan, A. Djian, A. Durand and P. Bonnarme, 1997. Influence of cell immobilization on the production of benzaldehyde and benzyl alcohol by the white-rot fungi Bjerkandera adusta, Ischnoderma benzoinum and Dichomitus squalens. Applied Microbiol. Biotechnol., 47: 708-714.
CrossRefDirect Link - Lawrence, G.D. and D. Yuan, 1996. Benzaldehyde formation from aspartame in the presence of ascorbic acid and transition metal catalyst. J. Agric. Food Chem., 44: 3461-3466.
Direct Link - Groot, M.N.N. and J.A.M. de Bont, 1998. Conversion of phenylalanine to benzaldehyde initiated by an aminotransferase in Lactobacillus plantarum. Applied Environ. Microbiol., 64: 3009-3013.
Direct Link - Simmonds, J. and G.K. Robinson, 1998. Formation of benzaldehyde by Pseudomonas putida ATCC 12633. Applied Microbial. Biotechnol., 20: 353-358.
CrossRefDirect Link - Swift, M.J., 1973. The estimation of mycelial biomass by determination of the hexosamine content of wood tissue decayed by fungi. Soil Biol. Biochem., 5: 321-332.
CrossRefDirect Link