
Subhash Chandra Ghosh
Faculty of Agriculture, Kagawa University, Miki cho, Kagawa Ken 761-0795, Japan
Koh-ichiro Asanuma
Faculty of Agriculture, Kagawa University, Miki cho, Kagawa Ken 761-0795, Japan
Akihito Kusutani
Faculty of Agriculture, Kagawa University, Miki cho, Kagawa Ken 761-0795, Japan
Masanori Toyota
Faculty of Agriculture, Kagawa University, Miki cho, Kagawa Ken 761-0795, Japan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 10 | Page No.: 1654-1657







ABSTRACT
The effects of different nitrogen levels (N0 = control, N1 = 101 Kg ha–1 and N2 = 202 kg ha–1) on net photosynthesis (PN), stomatal conductance (SC), transpiration rate (TR), water use efficiency (WUE) and leaf chlorophyll (LC, SPAD value) were studied in field grown potatoes (Solanurn tuberosum L. cvs. May Queen and Dejima). PN increased by the application of N to a smaller extent over control. In all cases, trend of PN initially increased and reached the peak at the time of tuber initiation and then gradually decreased. SC also increased in the N treated plant. The TR increased sequentially as fertilizer N level increased and WUE also increased by N treatment. Nitrogen fertilizer effect was more pronounced in case of LC. Higher N treated plant showed the higher levels of LC at all stages of growth. PN also showed highly significant positive correlation with SC, TR, WUE and LC. Between the cultivar PN, SC, TR and LC is comparatively higher in Dejima than May Queen. In conclusion, PN increased through the increase of SC resulted by the increase of TR, WUE and substantially increased the LC.
PDF Abstract XML References Citation
How to cite this article
Subhash Chandra Ghosh, Koh-ichiro Asanuma, Akihito Kusutani and Masanori Toyota, 2000. Gas Exchange Properties of Potato Plants under Different Nitrogen Regimes. Pakistan Journal of Biological Sciences, 3: 1654-1657.
DOI: 10.3923/pjbs.2000.1654.1657
URL: https://scialert.net/abstract/?doi=pjbs.2000.1654.1657
DOI: 10.3923/pjbs.2000.1654.1657
URL: https://scialert.net/abstract/?doi=pjbs.2000.1654.1657
INTRODUCTION
Nitrogen plays a vital roles in the growth and development of the crops as it required in large amounts in the non-legume crops. Therefore, judiciary supply of N fertilizer is important for proper growth and economic benefits of the users and also for the higher photosynthate production. Many factors known to affect the photosynthetic capacity, such as CO2 supply, temperature, leaf water status and illumination (EL-Sharkway and Hesketh, 1964; Gaastra, 1959; Hesketh and Moss, 1963) may act directly as limiting variables to photosynthesis. In leaves of higher plants photosynthesis is regulated strongly by the environment interacting with the genetic potential of the plants.
The influence of mineral nutrition on photosynthesis of plants has been reviewed by Natr (1972) wherein examples of increased rates of photosynthesis with increasing amounts of nutrients are cited. Andreeva et al. (1971) also reported that photosynthetic rates of potatoes increased with nutrient solution strength up to an optimum concentration. Increased photosynthetic rates with increasing amounts of applied N were, however, not found in field grown wheat (Thomas and Thorne, 1975; Pearman et al., 1979; Gregory et al., 1981). Low N supply reduces markedly the rate of CO2 assimilation by sugar beet leaves (Nevins and Loomis, 1970). Firman and Allen (1988) reported photosynthetic rates of leaves of the field potato plants grown without applied N reached levels as high as those receiving N. Dwelle et al. (1981a, b) have found significant differences in photosynthetic rates among potato clones. According to Mihhorpe (1963) the rate of photosynthesis of potato crops after tubers are formed is controlled by the growth of the tubers, the chief sink of photosynthetic products.
Still there are controversy remains between the N treatment and the photosynthetic rates of potato plants. Therefore, we studied the other physiological properties together with photosynthesis for better understanding of the effects.
The Chlorophyll meter (SPAD) provides a simple, quick and nondestructive methods for estimating leaf chlorophyll content (Watanabe et al., 1980). The SPAD meter has been widely used to predict the need for additional N fertilizer for many crops (Turner and Jund, 1991; Arcia et al., 1996; Peng et al., 1996; Piekielek et al., 1995; Fox et al., 1994). For many species, more than 95% of the plant dry weight is derived from photosynthetically fixed CO2 (Zelitch, 1975).
Cieply (1976) concluded that assimilation rates can be used as a physiological criterion for rapid selection in potato breeding. Therefore, photosynthesis study is important for plant growth and higher yield. Many studies have been done on photosynthesis but combined studies on net photosynthesis, stomatal conductance, transpiration rate, water use efficiency and leaf chlorophyll have not been done too much in the field grown potatoes under different N levels. For that, a better understanding of the effect of variable N fertilizer on photosynthesis and other physiological properties and their interrelationship is important.
MATERIALS AND METHODS
This is the companion paper and the experimental details are described earlier (Ghosh et al., 2000). Treatment consisted of three N levels (N0 = control, N1 = 101 Kg ha–1 and N2 = 202 kg ha–1) and two potato cultivars arranged in a split-plot design.
The leaf photosynthesis and related gas exchange characteristics was measured throughout the growth period from the youngest fully expanded leaf (third or fourth leaf from the apex) by using a portable photosynthetic system, the LI-COR Primer 6200 (LI-COR Inc. USA) on sunny days between 10.00 am to 2.00 pm. Water use efficiency was calculated as the ratio of photosynthesis to transpiration rate (Paez and Gonzalez, 1995).
Leaf chlorophyll was measured by using Minolta SPAD-502 meter (Minolta Co. Ltd., Japan) to the same leaf after the completion of photosynthesis measuremoms. All the statistical analyses were performed by the software MSTAT-C package (Michigan State University, USA).
RESULTS
Net photosynthesis (PN): The two cultivars showed a similar trend of increase of PN by the N treated plants (Table 1). Higher levels of N treated plants sometimes showed higher PN than control and the N1 treated plants sometimes possessed the higher levels of PN. But there was no consistent level of increase was observed between the N1 and N2 treatment. PN initially increased and reached the peak at 24 DAE (days after emergence) and then gradually decreased. Cultivar Dejima showed slightly higher levels of PN than May Queen and the rate of decrease was relatively slower than May Queen at the later stage of growth.
| Table 1: | Effect of different N levels on net photosynthesis (μ mol m–2 s–1) of two potato cultivars |
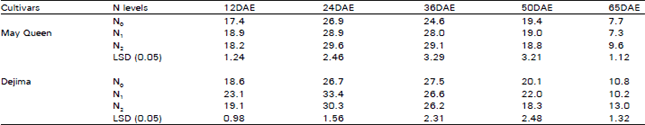 | |
| DAE: days after emergence, + Lsd (0.05): Least siginificant difference at 5% probability level | |
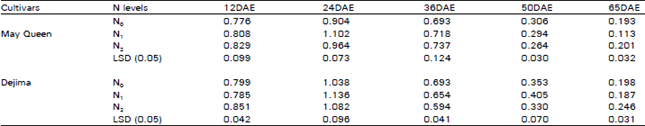
| Table 2: | Effect of different N levels on stomatal conducatance (μ mol m–2 s–1) of two potato cultivars |
 | |
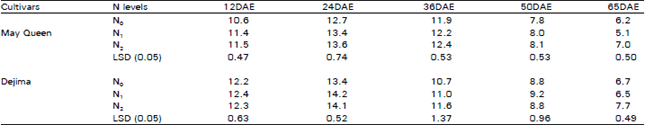
| Table 3: | Effect of different N levels on transpiration rate (μ mol m–1 s–1) of two potato cultivars |
 | |
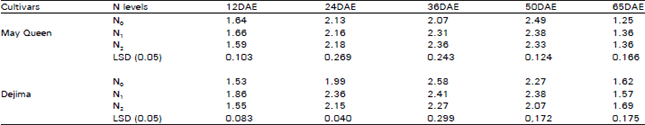
| Table 4: | Effect of different N levels on water use efficiency (μ mol/m mol) of two potato cultivars |
 | |
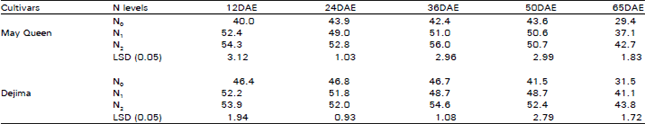
| Table 5: | Effect of different N levels on leaf chlorophyll (SPAD value) of two potato cultivars |
 | |
Stomatal conductance (SC): Treatment effects on SC were similar to those observed for PN. Increased PN also increases the SC of plants (Table 2). SC showed the similar trend of increase by the N treated plant. But the level of increase was sometimes higher in N1 and sometimes in the N2 treatments. SC initially increased and reached the peak at the time of tuber initiation and then gradually decreased. SC, was also higher in Dejima than May Queen.
Transpiration rate (TR): Higher N fertilizer treated plant showed higher TR in both cultivars throughout the growing season (Table 3). The difference between the N1 and N2 treatments is much lower than the difference in N0 and N1, N2 treatments. Like PN, TR value was also reached the peak at the time of tuber initiation and then gradually decreased. TR value was also higher in Dejima than May Queen but the trend was similar in both cultivars.
| Table 6: | Correlation coefficients between photosynthesis and other physiological properties of two potato cultivars under different N levels (n = 15) |
 | |
**: Indicates significant at 0.01 level, Pn: net photosynthesis, Sc: stomatal conducatnce, TR: transpiration rate, WUE: water use elficiencey, LC: leaf chlorophyll | |
Water use efficiency (WUE): The WUE also increased by the applied N1 but did not maintain any consistent increase between N1 and N2 treatment throughout the season (Table 4). WUE initially increased and reached the peak at 36 DAE and 50 DAE in Dejima and May Queen respectively. WUE was also relatively higher in Dejima than May Queen.
Leaf chlorophyll (LC): The increase in LC with increased N supply was observed in both cultivars throughout the growing season (Table 5). The difference between the N1 and N2 is lower than the difference between N0 and N1, N2. The LC reached the maximum at the same time in both cultivars. The LC value reached the peak at 12 DAE, 24 DAE and 36 DAE in N1, N0 and N2 treatments respectively. LC value was slightly higher in Dejima than May Queen.
Relationship between photosynthesis and other properties: The highly significant correlation was observed between PN and other properties and the relationship was similar in both the cultivars. The PN showed significant positive correlation with SC, TR, WUE and LC (Table 6).
DISCUSSION
Our results clearly indicate the increased PN by the application of nitrogen fertilizer which agree with the report of Nevins and Loomis (1970) in sugar beet. But in contrast, rates of photosynthesis in plant leaves grown without applied N reached levels as high as those receiving N and suggesting that arable soil N replenish the photosynthesis rates (Firman and Allen, 1988), This discrepancy of results might be due to the growth pattern and growing conditions of the plants. Here potato growing season is nearly half of the Firman and Allen (1988) experiment.
The gas exchange of plants supplied with N in the N1 and N2 did not differ too much in the growing season for both cultivars. The effects of N1 and N2 treatments is not similar for all cases. In some cases, N1 treated plant showed the higher rates of photosynthesis and sometimes N2 treated plant showed the higher rates of photosynthesis. This difference might be due to the cultivar, growth of the leaf and major sink organs (tuber).
Higher photosynthesis value is associated with the increased SC and LC resulted by the increase of TR and also WUE under higher N treated conditions. Higher SC increased the TR of the potato plants. But in all cases, the higher N treated plant did not show higher photosynthetic rates.
The PN value reached the maximum at the same time in all treatments and both the cultivars. This might be related to the initiation of tuber of the cultivar. The higher photosynthetic rate might be related to higher sink capacity of the growing tuber after their formation (Mihhorpe, 1963).
The increased N treated plant (N2) resulted in the increased vegetative growth than that of tuber growth and produced higher amount of total dry matter (Ghosh et al., 2000). And this higher dry matter production is related to the higher PN and LAI of the increased N treated plant, which initiated vegetative growth instead of tuber growth. So, the increase in dry matter production by N application is therefore achieved through effects of higher PN and leaf area duration.
Among the properties studied, LC increased substantially higher by the application of N fertilizer. This is related to the higher levels of leaf N or the higher protein content of the chloroplast. Increase of LC value is directly related to the increase of leaf N of the treated plant (Ghosh et al., 2000). Increased LC resulted in the increased PN but the increase was not consistent and might be due to the higher leaf or vegetative growth, as whole plant PN increased by N2 treatment. It is well known that N plays a key role in the establishment and maintenance of photosynthetic capacity of many crops. If no other restrictive factors are present, an increase in N supply would increase the growth, LC content and photosynthetic capacity of the leaves (Below, 1994). Thus once plant are established, sufficient N must be available to maintain the higher photosynthetic apparatus and its activity. In the present study Dejima contained higher leaf chlorophyll than May Queen and maintained higher PG and produced higher dry matter.
The decline in photosynthesis with plant age has been reported in many crops including potato (Vos and Qyarzun, 1987). Here PG decreased relatively slower in Dejima than May Queen. This might be due to the difference in plant type and growth habit of the cultivars (Fonseka and Asanuma, 1996). It has been shown that leaf N status is closely related to the LC for many crops (Piekielek and Fox, 1992; Turner and Jund, 1991), as most of the N in the leaf is associated with the protein in the chloroplast. The highly significant correlation between PN and LC can therefore be related with close association with the N status of the leaf. Photosynthesis also showed significant correlation with SC, TR and WUE.
The photosynthesis of potato leaves must be increased by the applied N, but in the field condition available N in arable soil is not low enough to limit leaf photosynthesis of crop plants directly. Therefore, applied N in this experiment did not increase the PN too much. But photosynthesis increased in the early grown potato by the application of N fertilizer up to certain level. SC and WUE were increased and more pronouncedly the leaf LC and TR were increased by N. It is suggested that judiciary application of. N fertilizer can improve the tuber yield through higher photosynthesis if other factors remain sufficient.
REFERENCES
- Dwelle, R.B., G.E. Kleinkopf and J.J. Pavek, 1981. Stomatal conductance and gross photosynthesis of potato (Solanum tuberosum L.) as influenced by irradiance, temperature and growth stage. Potato Res., 24: 49-59.
CrossRefDirect Link - Dwelle, R.B., G.E. Kleinkopf, R.K. Steinhorst, J.J. Pavek and P.J. Hurley, 1981. The influence of physiological processes on tuber yield of potato clones (Solanum tuberosum L.): Stomatal diffusive resistance, stomatal conductance, gross photosynthetic rate, leaf canopy, tissue nutrient levels and tuber enzyme activities. Potato Res., 24: 33-47.
CrossRefDirect Link - Firman, D.M. and E.J. Allen, 1988. Field measurements of the photosynthetic rate of potatoes grown with different amounts of nitrogen fertilizer. J. Agric. Sci., 111: 85-90.
CrossRefDirect Link - Fox, R.H., W.P. Piekielek and K.M. Macneal, 1994. Using a chlorophyll meter to predict nitrogen fertilizer needs of winter wheat. Commun. Soil Sci. Plant Anal., 25: 171-181.
CrossRefDirect Link - Gaastra, P., 1959. Photosynthesis of crop plants as influenced by light, carbon dioxide, temperature and stomatal diffusion resistance. Mededelingen Landbouwhoegsch Wageningen, 59: 1-68.
Direct Link - Ghosh, S.C., K.I. Asanuma, A. Kusutani and M. Toyota, 2000. Nitrogen distribution and uptake efficiency traits of potato under different nitrogen regimes. Pak. J. Biol. Sci., 3: 943-948.
CrossRefDirect Link - Arcia, F.V., S. Peng, H.C. Gines, R.C. Laza, A.L. Sanico, A.M. Visperas and K.G. Cassman, 1996. Chlorophyll Meter Based Nitrogen Management Improves Nitrogen use Efficiency of Irrigated Rice in Farmer's Fields. In: Crop Research in Asia: Achievements and Perspective, Ishii, A. and T. Horie (Eds.). Asian Crop Science Association, Fukui, Japan, pp: 187-190.
- Gregory, P.J., B. Marshall and P.V. Biscoe, 1981. Nutrient relations of winter wheat: 3. Nitrogen uptake, photosynthesis of flag leaves and translocation of nitrogen to grain. J. Agric. Sci., 96: 539-547.
CrossRefDirect Link - Paez, A. and M.E. Gonzalez, 1995. Water stress and clipping management effects on Guineagrass: II. Photosynthesis and water relations. Agron. J., 87: 706-711.
CrossRefDirect Link - Pearman, I., S.M. Thomas and G.N. Thorne, 1979. Effect of nitrogen fertilizer on photosynthesis of several varieties of winter wheat. Ann. Bot., 43: 613-621.
CrossRefDirect Link - Peng, S., F.C. Garcia, R.C. Laza, A.L. Sanico, R.M. Visperas and K.C. Cassman, 1996. Increased N-use efficiency using a chlorophyll meter on high-yielding irrigated rice. Field Crops Res., 47: 243-253.
CrossRefDirect Link - Piekielek, W.P. and R.H. Fox, 1992. Use of a chlorophyll meter to predict sidedress nitrogen requirements for maize. Agron. J., 84: 59-65.
Direct Link - Piekielek, W.P., R.H. Fox, J.D. Toth and K.E. Macneal, 1995. Use of a chlorophyll meter at the early dent stage of corn to evaluate nitrogen sufficiency. Agron. J., 87: 403-408.
Direct Link - Thomas, S.M. and G.N. Thorne, 1975. Effect of nitrogen fertilizer on photosynthesis and ribulose 1, 5-diphosphate carboxylase activity in spring wheat in the field. J. Exp. Bot., 26: 43-51.
CrossRefDirect Link - Turner, F.T. and M.F. Jund, 1991. Chlorophyll meter to predict nitrogen topdress requirement for semidwarf rice. Agron. J., 83: 926-928.
Direct Link - Vos, J. and P.J. Oyarzun, 1987. Photosynthesis and stomatal conductance of potato leaves-effects of leaf age, irradiance and leaf water potential. Photosynth. Res., 11: 253-264.
CrossRefDirect Link - Zelitch, I., 1975. Improving the efficiency of photosynthesis. Science, 188: 626-633.
CrossRefDirect Link