
S. Shahid Shaukat
Department of Botany, University of Karachi, Karachi-75270, Pakistan
G. R. Sarwar
Department of Botany, University of Karachi, Karachi-75270, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 10 | Page No.: 1648-1653







ABSTRACT
The deposits of silicon in the leaves of several grasses and sedges have been investigated. Shape and distribution patterns of opal phytoliths were examined in the adaxial surface of leaf blades. Dumb- bell shape was found to be most common, such phytoliths were generally arranged in two or three rows. Rod, sphere and saddle-shaped phytoliths were specific for certain species and this can be used as a diagnostic character to delimit such species.
PDF Abstract XML References Citation
How to cite this article
S. Shahid Shaukat and G. R. Sarwar, 2000. Opaline Silica in the Leaves of Some Grasses and Sedges of Karachi. Pakistan Journal of Biological Sciences, 3: 1648-1653.
DOI: 10.3923/pjbs.2000.1648.1653
URL: https://scialert.net/abstract/?doi=pjbs.2000.1648.1653
DOI: 10.3923/pjbs.2000.1648.1653
URL: https://scialert.net/abstract/?doi=pjbs.2000.1648.1653
INTRODUCTION
The plant opals are microscopic amorphous bodies which occur in living plants. The term "phytoliths" is used to describe plant opals having repetitive shapes. Phytoliths survive in sediments and soils and have therefore been used as indicators of paleosols, in archaeological reconstructions and as indicators of past vegetation. Classification of the opal was on a morphological basis since in soil materials such opal become fragmented and less reliably identified as being derived from a particular cell. They identified rods, spheres, etc. and in some cases related them to various cells within the plants, although not to a particular species. Assemblages of phytoliths in soils have also been used to assist in identification of the originating flora (Rovner, 1983).
The occurrence of silica deposits in plants has been investigated by many workers. Comprehensive studies have been conducted on Zea mays (Lanning et al., 1980); on cereals (Bennett and Parry, 1980); on Zea mays (Bennett and Sangster, 1981); on higher plants (Kaufman et al., 1981) and on Miscanthus sacchariflorus (Sangster, 1983). Metcalfe and Chalk (1983) investigated opaline silica in angiospermic trees and shrubs. Parry et al. (1985) reported the distribution of silicon deposits in the fronds of Pteridium aqullinium. Hart (1988) studied trees and shrubs of Australia and (Borre and Watson, 1994) investigated opaline silica in leaves of Eragrostis. Watson and Dallwitz (1993) used opaline silica for identification of grasses. Silica deposition in rice (Oryza swivel L.) has been extensively studied (Whang, 1993; Whang and Kim, 1994; Whang et al., 1998).
The present study examines the occurrence, structure and distribution of phytoliths in the leaves of some grasses and sedges commonly found in Karachi, Pakistan.
MATERIALS AND METHODS
Fifteen grass species and two sedge species were collected from different localities in Karachi. Voucher specimens have been deposited in Plant Ecology Laboratory at the Department of Botany, University of Karachi, Pakistan. The species studied are listed below: The technique used for making surface preparations was similar to that employed for the study of opaline silica in the leaves of grasses (Parry and Smithson, 1958, 1964). Leaves were washed with distilled water and small pieces were transferred onto the glass slides. A few drops of dilute chromic acid were added and the material left for about 15 minutes. The tissues were washed with distilled water and mounted in glycerine. The adaxial surfaces of leaves were examined under light microscope (x10, x40) and the shape and distribution of opaline phytoliths were observed. The length and breadth of opaline phytoliths were measured using occular micrometer after calibration. Photographs were taken at x10 and x40 magnifications.
In order to assess the phonetic similarities among the taxa on the basis of phytolith characteristics and their distribution patterns, a cluster analysis was performed. Gower (1971) similarity coefficient was used as it has the ability to deal with mixed data types. Hierarchical agglomerative clustering was accomplished using average linkage clustering strategy (Everitt, 1980).
RESULTS
The opaline silica bodies in the leaves of the 17 species studied (15 grasses and 2 sedges) exhibited considerable variation with respect to shape, distribution pattern, size, density and position on leaves (Table 1). In grass leaves some of the costal cell (long silica cells) and some of the intercostal cells (short silica cells) usually forming part of silico-suberose couples normally acquire silica at an early stage and in a regular manner. The bodies vary in form, straight-sided to lobed, with two to three lobes on each edge. The various shapes of opaline silica were observed, including dumb-bell shaped, rod-shaped square and saddle-shaped. Since the shape of phytolith is the most consistent and a diagnostic feature, further results are described in sequel with respect to shape.
The various shapes of the plant opal found are described in the sequel.
Dumb-bell shaped phytoliths: The adaxial leaves of some grasses showed well aligned bodies usually with two lobes on each edge forming the so called (dumb-bells) and also three lobes edges were seen. Dumb-bell shaped phytoliths were found in Panicum turgidum (Fig. 3a, b), Eragrostis poaeoides (Fig. 1e), Sporobolus marginatus (Fig. 3d), Setaria verticillata (Fig. 3e), Cyperus bulbosus (Fig. 2a, b), Lasiurus hirsutus (Fig. 1f), Cynodon dactylon (Fig. 1b) and Aristide mutabllis (Fig. 1a). Opal phytoliths were arranged in chains in Panicum turgidum (Fig. 3a, b), Eragrostis poaeoides (Fig. 1e), Cyperus bulbosus (Fig. 2a, b), Lasiurus hirsutus (Fig. 1f), Cynodon dactylon (Fig. 1b) and Aristide mutabllis (Fig. 1a) while in Sporobolus marginatus (Fig. 3d) and Setaria verticillata (Fig. 3e) dumb-bell shaped phytoliths were not arranged in chain. Three lobed phytoliths were found in Dichanthium annulatum (Fig. 2g, h), Dactyloctenium aegyptium (Fig. 1h) and Cenchrus pennisetiformis (Fig. 1d, Table 1).
Rod-shaped or Rectangular-shaped phytoliths: Rod-shaped phytoliths was found in only one species i.e. Eragrostis pilosa (Fig. 2d). They were distributed sparsely and had smooth surface (Table 1).
Sphere-shaped or Rounded phytoliths: Cyperus rotundus (Fig. 3c) showed only sphere or rounded phytoliths. They were arranged in one row. All the sphere were approximately of the same size (Table 1).
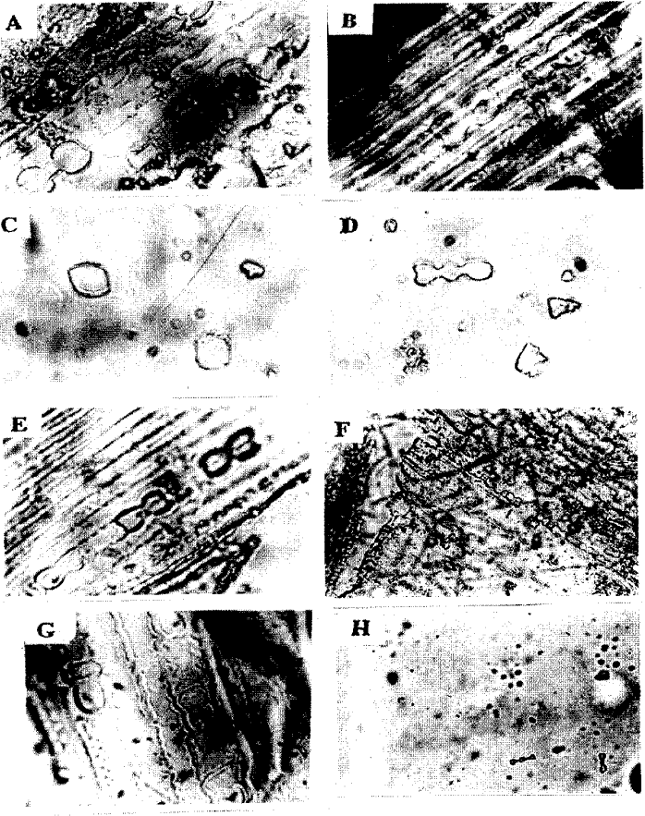 | |
| Fig. 1: | Light micrographs showing opaline silica in the adaxial surface of leaves. A. Aristida mutabilis x600, B. Cynodon dactylon x600, C. Chloris barbata x600, D. Cenchrus pennisetiformis x600, E. Eragrostis poaeoides x600, F. Lasiurus hirsutus x150, G. Saccharum munja x600, H. Dactyloctenium aegyptium x150 |
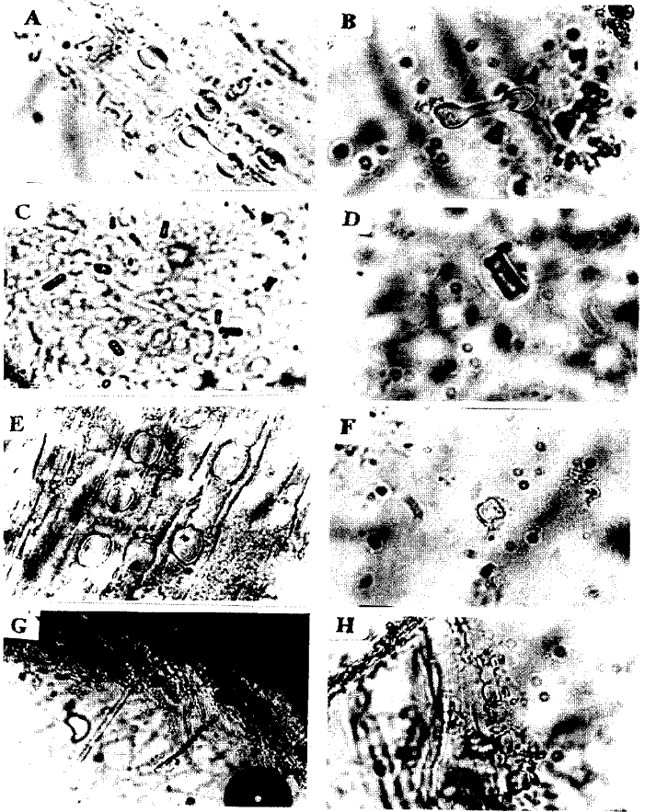 | |
| Fig. 2: | Light micrographs showing opaline silica in adaxial surface of leaves. A. Cyperus bulbosus x600, B. Cyperus bulbosus x600, C. Eragrostis pilosa x150, D. Eragrostis pilosa x600, E. Brachiaria ramosa x600, F.Brachiaria ramosa x600, G. Dichanthium annulataum x150, H. Dichanthium annulatum x600 |
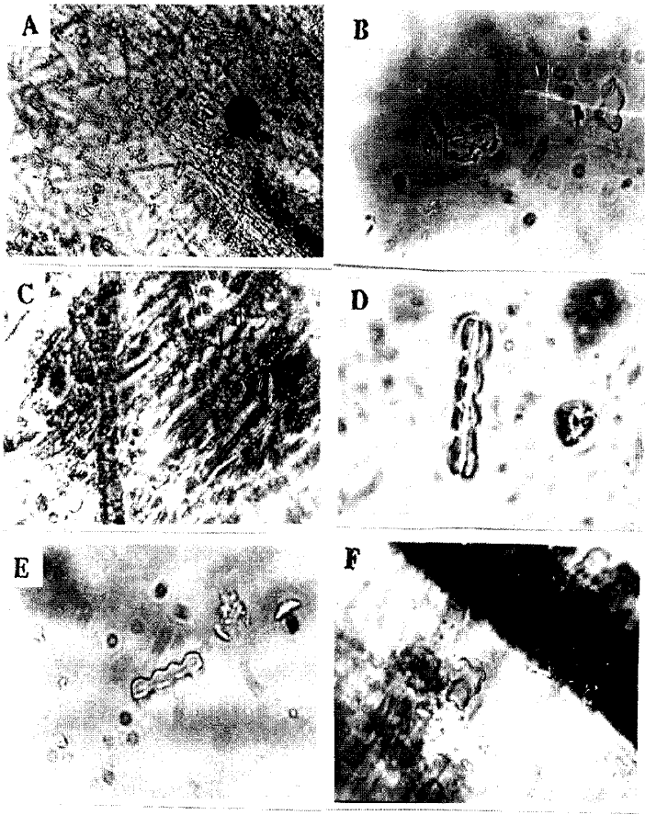 | |
| Fig. 3: | Light micrographs showing opaline silica in the adaxial surface of leaves. A. Panicum turgidum x150, B. Panicum turgidum x600, C. Cyperus rotundus x600, D. Sporobolus marginatus x600, E. Setaria verticillata x600, F. Andropogon aucheri x600 |
| Table 1: | Name of Species collected from different localities |
 | |
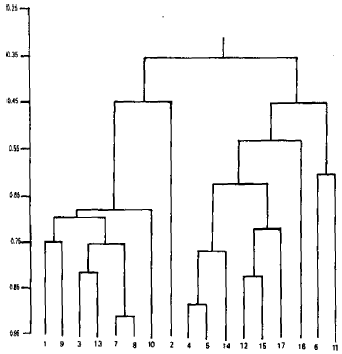 | |
| Fig. 4: | Dendrogram derived from agglomerative hierarchical clustering of 17 species based on opaline silica characteristics |
Square-shaped phytoliths: Square-shaped phytoliths were observed in Chloris barbata (Fig. 1c) and Bracharia ramosa (Fig. 2e, f). Opal phytoliths were arranged in two to three rows in Bracharia ramosa (Fig. 2e, f) whereas in Chloris barbata (Fig. 1c), they were sparsely scattered. Andropogon aucheri (Fig. 3f) was the only species which had square-shaped phytolith with an appendage (Table 1).
Saddle-shaped: Saddle-shaped phytoliths were found only in one species i.e. Saccharum munja (Fig. 1g). They were costal, arranged in three rows on adaxial surface.
The dendrogram resulting from the average linkage clustering (Fig. 4) showed two major groups I and II. Group I usually includes taxa with either randomly distributed phytoliths or arranged in one row, with their length ranging from 20 to 30 μm and density from 1-3 phytoliths per μm and with mostly intercostal arrangement. By contrast group II comprises of taxa with phytpliths usually arranged in two to three rows, with lengths greatr than 30 μm and density mostly between 3-7 per μm with costal distribution.
DISCUSSION
The use of light microscope to study intact and isolated silica deposits in the leaves of different species adds significantly to our knowledge of their shape, structure and distribution pattern. During studies, five types of opaline phytoliths were observed i.e. dumb-bell-shaped, rod-shaped, square-shaped, sphere-shaped and saddle-shaped. These shapes have previously been noticed by earlier workers in different grasses (Baker, 1959; Twiss et al., 1969; Blackman, 1971; Sangster, 1978).
Dumb-bell shape opaline phytoliths was found to be most common. It was found in 8 out of 17 taxa studied. Whereas rod, sphere and saddle- shaped phytoliths were found in one species each. Dumb-bell-shaped phytoliths were further classified with respect to their distribution. In Dichanthium annulatum, Panicum turgidum, Eragrostis poaeoides, Bracharia ramose, Cyperus rotundus, Cyperus bulbosus, Lasiurus hirsutus, Cynodon dactylon, Saccharum, nunja and Aristida mutabilis, they were arranged in one to three rows in the form of chains, while in Chloris barabata, Eragrostis pilosa, Sporobolus marginatus, Setaria vertiala, Andropogon aucheri, Dactyloctenium aegyptium and Cenchrus pennisetiformis, they were distributed sparsely and without any definite pattern. In some species the lobes were bigger and the connecting rod was small e.g. Aristide mutabilis (Fig. 1a), while in Panicum turgidum (Fig. 1a, b), the lobes were smaller with the connecting rod relatively longer e.g. Cyperus bulbosus. The dumb-bell shape is considered diagnostic for the Gramineae by Twiss et al. (1969). The variation in the dumb-bell-shaped observed could have a great potential as a diagnostic character, in addition to other diagnostic characters. Geis (1973) considered that silicified epidermal hairs were lacking in grasses and found in deciduous forest species. Thus this character could be used to distinguish between grass species and deciduous forest species. Sphere, rod and saddle shapes found as less frequent forms could have a potential as diagnostic characters that could be helpful for identification purposes (Rapp and Mulholland, 1992). There is a great need for the study of the phytoliths in other grasses and sedges of this region to disclose this type of information of diagnostic value. In addition, the charateristics of opaline silica observed here can be used for identification of species during nonflowering season.
REFERENCES
- Baker, G., 1959. Opal phytoliths in some Victorian soils and red rain residues. Aust. J. Bot., 7: 64-87.
CrossRefDirect Link - Blackman, E., 1971. Opaline silica bodies in the range grasses of southern Alberta. Can. J. Bot., 49: 769-781.
CrossRefDirect Link - Bennett, D.M. and A.G. Sangster, 1981. The distribution of silicon in the adventitious roots of the bamboo Sasa palmata. Can. J. Bot., 59: 1680-1684.
CrossRefDirect Link - Bennett, D.M. and D.W. Parry, 1980. Electron-probe microanalysis studies of silicon in the elongating basal internodes of Avena sativa (L.), Hordeum sativum (Jess.) and Triticum aestivum (L.). Ann. Bot., 45: 541-547.
CrossRefDirect Link - Geis, J.W., 1973. Biogenic silica in selected species of deciduous angiosperms. Soil Sci., 116: 113-130.
Direct Link - Gower, J.C., 1971. A general coefficient of similarity and some of its properties. Biometrics, 27: 857-871.
CrossRefDirect Link - Hart, D.M., 1988. The plant opal content in the vegetation and sediment of a swamp at Oxford Falls, New South Wales, Australia. Aust. J. Bot., 36: 159-170.
CrossRefDirect Link - Lanning, F.C., T.L. Hopkins and J.C. Loera, 1980. Silica and ash content and depositional patterns in tissues of mature Zea mays L. plants. Ann. Bot., 45: 549-554.
CrossRefDirect Link - Parry, D.W. and F. Smithson, 1958. Techniques for studying opaline silica in grass leaves. Ann. Bot., 22: 543-549.
CrossRefDirect Link - Parry, D.W. and F. Smithson, 1964. Types of opaline silica depositions in the leaves of British grasses. Ann. Bot., 28: 169-185.
CrossRefDirect Link - Parry, D.W., M.J. Hodson and R.H. Newman, 1985. The distribution of silicon deposits in the fronds of Pteridium aquilinum L. Ann. Bot., 55: 77-83.
CrossRefDirect Link - Rovner, I., 1983. Plant opal phytolith analysis: Major advances in archaeobotanical research. Adv. Archaeol. Method Theory, 6: 225-266.
Direct Link - Sangster, A.G., 1983. Silicon distribution in the nodal roots of the grass Miscanthus sacchariflorus. Can. J. Bot., 61: 1199-1205.
Direct Link - Whang, S., K. Kim and W. Hess, 1998. Variation of silica bodies in leaf epidermal long cells within and among seventeen species of Oryza (Poaceae). Am. J. Bot., 85: 461-466.
Direct Link