
Nazeer Hussain Shah
Agricultural Research Institute, Dere Ismail Khan, Pakistan
Gary M. Paulsen
Department of Agronomy, Kansas State University, Manhattan, Kanas, 66506, USA
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 10 | Page No.: 1639-1643







ABSTRACT
High temperature and drought affect plant processes individually and may interact to alter water relations. Our objectives were to ascertain the effect of high temperature and osmotic stress and to identity interactions between them on water relation of wheat (Triticum aestivum L. cv. Len). Plants that were grown uniformly until anthesis were subjected to 15/10, 25/20 or 35/30°C and -0.01 or 0.41 Mpa water stress and were sampled weekly until maturity. Relative water content (RWC) of flag leaves remained constant at low temperature and declined quickly at high temperature. Osmotic stress decreased RWC slowly at low temperature and rapidly at high temperature as plants matured. Water potential and osmotic potential reacted similarly, staying high at low temperature and falling faster as the temperature increased or osmotic stress impinged. Turgor potential was sometimes higher under 15/10°C with osmotic stress than under other regimes, but varied lesser than other pressure components. Cumulative water use ranked 25/20 >15/10 >35/30°C without osmotic stress >15/10°C = 25/20°C = 35/30°C with osmotic stress. Water use efficiency for grain mass was reduced by high temperature and increased by osmotic stress. Results demonstrated that high temperature and osmotic stress interact by complementary effect on water relations and beneficial effects of low temperature on osmotic adjustment to stress.
PDF Abstract XML References Citation
How to cite this article
Nazeer Hussain Shah and Gary M. Paulsen, 2000. Response of Wheat to Combined High Temperature an Osmotic Stress During Maturation. II. Plant Water Relations. Pakistan Journal of Biological Sciences, 3: 1639-1643.
DOI: 10.3923/pjbs.2000.1639.1643
URL: https://scialert.net/abstract/?doi=pjbs.2000.1639.1643
DOI: 10.3923/pjbs.2000.1639.1643
URL: https://scialert.net/abstract/?doi=pjbs.2000.1639.1643
INTRODUCTION
High temperature affects plants directly by injuring photosynthesis and other plant processes (AL-Khatib and Paulsen, 1989). Many indirect effects of high temperature are also evident, The most important among those are on plant water relations. Evaporative loss of Water from soil by high temperature diminishes available moisture for plants and exacerbates drought (Kumar et al., 1985; Hamblin et al., 1987). Increased transpiration loss at high temperature may induce drought even under well-watered conditions (Kramer, 1980) and accelerates depletion of soil moisture (Angus et al., 1983). Either effect eventually alters plant water relations and affects processes that depend on water (Kramer, 1980). Thus, it is not surprising that high temperature and osmotic stress interact to accentuate injury to photosynthesis, leaf viability, and plant productivity (Shah and Paulsen, 1999). Water status of plant is determined by cell water potential, the sum of the turgor pressure potential and the osmotic potential (Gusta and Chen, 1987). Water potential, the difference in free energy per unit.volume between osmotically constrained water and free water is always negative (Baker, 1984). Turgor potential arises by diffusion of water from external sources to the lower potential of cell solution and is always positive, whereas osmotic potential of cell solutes is always negative (Salisbury and Ross, 1985). In wheat, water potential, turgor potential and osmotic potential range from -0.5 to -1.6 Mpa, 0.2 to 1.2 Mpa, and -1.1 to -1.2 Mpa, respectively, in well watered plants (Gusta and Chen, 1987), Relative water content (RWC), the. ratio between the actual and fully turgid water contents of tissue, reflects the interplay between turgar potential and osmotic potential (Baker, 1984). Tissues accumulate solutes, primarily sugars, to lower their osmotic potential (osmotic adjustment) during stress (Gupta and Berkowitz, 1987). Osmotic adjustment enables photosynthesis tissues to maintain high relative water content and photosynthetic rate when moisture becomes deficient (Gupta and Berkowitz, 1987) and may be associated with wheat grain yield (Blum et al., 1983). Views differ on the utility of the different pressure components for assessing plant water status. All of them have been used successfully, however. Kramer (1980) and Sinclair and Ludlow (1985), for instance, concluded that water potential, turgor potential and osmotic potential accurately portrayed the behaviour of water. Relative water content, in comparison, is less controversial and detects significant differences that might not be received by pressure components (Schonfeld et al., 1988). High temperature and drought frequently occur simultaneously instead of singly. Knowledge of interaction between stresses is needed to distinguish their direct and indirect effects and to understand synergistic effects that might exacerbate injury. Objectives of these investigations were to ascertain water relations of wheat under different temperature and osmotic stress regimes during the grain filling period and to identify interactions that might mediate plant processes.
MATERIALS AND METHODS
Plant culture and treatments: Wheat plants were grown uniformly in a green house up to anthesis. Temperature treatments of 15/10, 25/20 or 35/30°C day/night and osmotic stress treatments of -0.01 or -0.41 Mpa from polyethylene glycol (PEG 8000) were imposed in controlled environment chambers after anthesis until maturity. Although, the experiments were conducted' separately, however, all conditions were similar to those for determining treatment effects on photosynthesis and productivity (Shah and Paulsen, 1999). Plants were sampled for water relation measurements when temperature and osmotic stress treatments were imposed and at weekly intervals afterwards until they matured.
Relative water content measurements: Leaf discs of 15 mm diameter were cut with a sharp corks borer from flag leaves of main stems of at least three plants in each treatment and weighed immediately. The discs were immersed in water under compensating illumination of 40 μmol m–2 s–1 PAR for 4 h, blotted with bibulous paper and weighed to obtain turgid weight. The discs were then dried at 70°C for 24 h and weighed. The RWC was calculated as fresh weight-dry weight % (turgid weight-dry weight) (Rhodes and Mastuda, 1976).
Water Components Measurements: Water potential and osmotic potential measurements were obtained by two Psychrometric methods, a screen cage thermocouple psychrometer chamber (Mode 174-13, J.R.D. Morill specialty equipment, Logan, Utah, U.S.A) and a C-52 thermocouple psychrometer chamber (Model 2740-1. We 21640' 1640, Logan, Utah, U.S.A). The two methods were used to verify results because their protocols differ. The screen cage has a large chamber and requires considerable time for equilibration, whereas, the C-52 instrument has a small chamber and equilibrate rapidly. Leaf discs of 15 mm diameter were cut with a sharp corks borer from flag leaves of main stems of three plants, placed in the screen cage thermocouple psyschrometer chamber and equilibrated for 3 h. Water potential was measured with a Wescor HR-33 microvolt-meter (Wescor, Inc.) operating in the psychrometric mode (Kirkham, 1989). Tissue used for water potential measurements was frozen in liquid N to obtain osmotic potential for determining osmotic adjustment. Sap was expressed by crushing the leaf discs between a glass rod and glass plate. Sap wis collected on Whatman 42 Filter paper, which was cut into discs with the same cork borers used for collecting leaf discs (Nus and Hodges, 1985). The paper with sap was immediately placed in the psychrometer chamber and osmotic potential readings were taken with the microvoltmeter again after equilibration for 3 h. Turgor potential was calculated as the difference between water potential and osmotic potential. Small leaf discs 6 mm in dia. were cut with a paper punch from flag leaves on main culms of plants and placed in the model C-52 psychrometer chamber. Samples were equilibrated for 5 min., after which the balance of the procedures for water potential and osmotic potential were similar to measurements with the screen cage chamber (Nus and Hodges, 1985). The same microvoitmeter was used for all measurements, and six readings of water potential and osmotic potential per treatment were taken by both methods.
Total water use and water use efficiency calculations: Addition of water to maintain plant containers at capacity were recorded daily and summed to determine total plant water use during the experimental period. All water consumption from the covered containers was attributed to transpiration. Water use efficiency was calculated as the ratio of grain mass at the final harvest to the cumulative water consumption.
Experimental design and data analyses: The experiment was arranged in completely randomized design of three day/night temperature regimes and two osmotic stress levels. All treatments were replicated three times. Treatment means were compared by least significant difference (LSD) test, when samples sizes were equal and by least squares when sample sizes were not equal.
RESULTS AND DISCUSSION
Flag leaves maintained 90% RWC at 15/10 and 25/20°C during the first week of treatment without osmotic stress (Fig. 1). Plants at the high temperature, however, lost considerable capacity to retain water. The RWC stayed near initial levels in plants at 15/10°C until the experiment was terminated after 70 d. It was also high in plants at 25/20°C for 42 d, after that it declined quickly. No water retention capacity was detected in plants at 35/30°C after the first week.
Osmotic stress did not affect RWC of plants at 15/10°C, but it decreased RWC slightly in plants at 25/20°C and quickly in plants at 35/30°C during the first week (Fig. 1). The RWC stayed high for 42 d in plants at 15/10°C and then decreased slowly and was still measurable after 63 d. The RWC in plants at 25/20°C continued to decline after the first week to a level after 21 d and in plants at 35/30°C, was not measurable after 7 d.
Water components measurements by the screen cage psychrometer: Water potential was high in plants at the low and intermediate temperature, but was low in plants at the high temperature after 7 d of treatment (Fig. 2a). It stayed high for 70 d in plants at 15/10°C and for 42 d in plants at 25/20°C and was not detectable after 7 d in plants at 35/30°C. Osmotic stress decreased water potential at the low and intermediate temperature, but did not diminish the already low values at the high temperature after 7 d. Water potential varied erratically after the first week in plants at 15/10°C and in plants at the same temperature was usually lower with osmotic stress than without osmotic stress. Values in plants under osmotic stress were still measurable after 63 d at 15/10°C; however they decreased linearly for 21 d at 25/20°C and were not detectable after 7 d at 35/30°C. Osmotic potential followed the same trends as water potential under all temperature and osmotic stress treatments (Fig. 2b). Osmotic potential was always lower than water potential for the same treatments on the same dates. For turgor potential, the difference between water potential and osmotic potential, was similar among treatments on most dates (Fig. 2c). Values for plants grown at 15/10°C under osmotic stress exceeded those for plants under other treatments on several dates during mid to late grain-filling stages, however.
Water components by the C-52 psyschrometer: Water potential values in plants after 7 d of treatments with out osmotic stress were highest at 15/10 and 25/20°C and decreased slightly at 35/30°C (Fig. 3a). Values stayed high for 70 d at 15/10°C, decreased abruptly after 42 d at 25/20°C and were not measurable after 7 d at 35/30°C. Osmotic stress did not change water potential significantly at any temperature after the first week. During the following weeks, values stayed high for 42 d and then decreased slowly at 15/10°C, decreased linearly from 7 to 21 d at 25/20°C and were not measurable after 7 d at 35/30°C.
Osmotic potential measured by the C-52 psychrometer, as by the screen cage psyschrometer, paralleled water potential closely (Fig. 3b). Values were again lower for osmotic potential than for water potential. Turgor potential differed little among treatments (Fig. 3c).
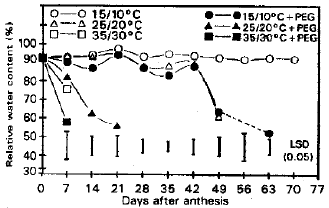 | |
| Fig. 1: | Relative water content (%) of flag leaves as affected by diffrent temperatures and osmatic pressures |
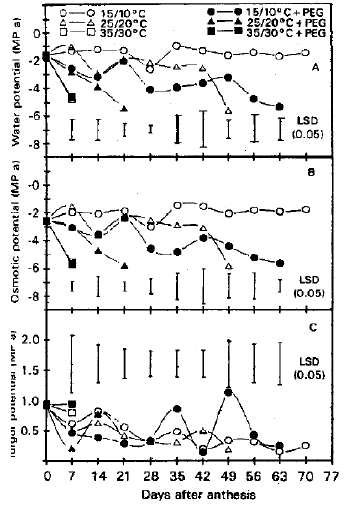 | |
| Fig. 2: | (a) Water potential (MPa) of flag leaves measured by screen Cage Psychrometer |
| (b) Osmatic Potential (MPa) of flag leaves measured by Screen Cage Psychrometer | |
| (c) Turgor Potential (MPa) of flag leaves measured by Screen Cage Psychrometer | |
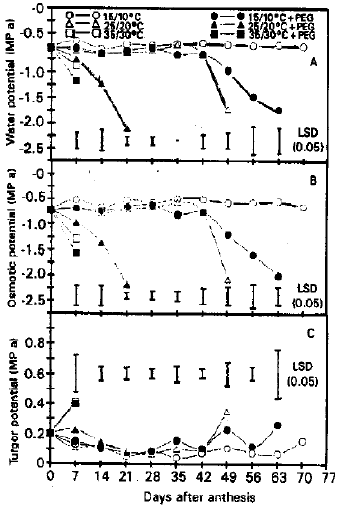 | |
| Fig. 3a: | (a) Water potential (MPa) of flag leaves measured by C-052 Phychrometer |
| (b) Osmatic Potential (MPa) of flag leaves measured by C-52 Psychrometer | |
| (c) Turgor Potential (MPa) of flag leaves measured by C-52 Psychrometer | |
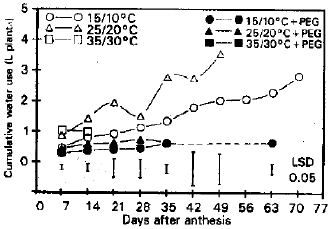 | |
| Fig. 4: | Relative water content 1%) of flag leaves as affected by diffrent temperatures and osmatic pressures |
Values were increased significantly only by high temperature with or with out osmotic stress after 7 d and were increased marginally by the high temperature with out osmotic stress and the low temperature with osmotic stress during later stages.
Plant cumulative water use and water use efficiency: Plants grown at 25/20°C and 35/30°C without osmotic stress consumed more water than plants under other treatments during the first week (Fig. 4). Cumulative water use rose steadily for 70 d at 15/10°C and for 49 d at 25/20°C when plants were not osmotically stressed. Little change was evident under other treatments as plant matured. Total water consumption during grain filling from highest to lowest was 25/20°C>15/10°C>35/30°C with out osmotic stress >15/10°C = 25/20°C = 35/30°C with osmotic stress. Water use efficiency was reduced by the high temperature treatment and was increased by osmotic stress at the low and intermediate temperature treatments. Values were o.72, 0.91 and 0.33 g grain L–1 without osmotic stress and 4.56, 2.53 and 0.65 g grain L–1 with osmotic stress at 15/10, 25/20 and 35/30°C, respectively. The LSD (0.05) for comparing treatment means was 1.27 g grain L–1.
Investigations of high temperature effect on plants usually emphasize direct response of photosynthesis (Al-Khatib and Paulsen, 1989; Harding et al., 1990a, b), reproductive growth (Nicolas et al., 1984; Wardlaw et al., 1989) and other metabolic processes (Gusta and Chen, 1987). Our results demonstrate that temperature also affects plant water relations and alters responses to osmotic stress. The interactions between temperature and osmotic stress have important consequences on plant photosynthesis, Productivity, and adaptation to unfavorable environments (Shah and Paulsen, 1999). Effects of high temperature, unlike those of osmotic stress, on plant water relations during the critical grain-filling period of wheat are not well documented. Many of the effects of high temperature were to those of osmotic stress in the present studies and to drought stress in other studies. Reduction in RWC, water potential, and water potential components with increasing temperature, for instance, resembled response of the same measurements to drought stress (Baker, 1984; Kramer, 1980; Schonfeld et al., 1988; Shimshi et al., 1982). The response that differed most between the two stresses, cumulative water use, undoubtedly reflected temperature effects on enhanced water loss and osmotic effects on diminished water uptake and loss. Because high temperature by itself did not induce stomatal closure until leaves senesced (Shah and Paulsen, 1999), consumption of water for transpirational cooling would be expected to increase markedly (Gates, 1968). Low water potential from the PEG osmoticum, on the other hand , reduced both water uptake from the medium and water loss from the leaves (Baker, 1984; Kramer, 1980). The other contrast between the two treatments, water use efficiency, arose from differences in their quantitative effects on grain yield and their qualitative effects on water consumption. Strong synergistic interactions between high temperature and osmotic stress were evident. Combining the two stresses exacerbated most measures of plant water relations as severely as most parameters of photosynthesis (Shah and Paulsen, 1999) and reproductive development (Nicolas et al., 1984). The interaction originated from the complementary effects of high temperature and osmotic stress on plant water relations and from beneficial effects of low temperature on plant adjustment to osmotic stress. Complimentary effects of high temperature and osmotic stress suggested that the treatments affected RWC, water potential and osmotic potential in the same basic manner. Combining treatments thus compounded the response to either one alone. The RWC, a measure of cell hydration, for instance, was reduced by enhanced loss of water for transpirational high temperature, by diminished gain of water from osmotic stress and by an additive amount when the two treatments were combined. Osmotic potential may have reacted similarly to the treatments' effects on cell water volume and solute concentration to alter turgor and water potential. Osmotic adjustment, defined as an increase in osmotic pressure from more solute molecules per cell (Munns, 1988), enables plants to maintain turgor pressure at low water potential (Johnson et al., 1984; Morgan, 1980). Plants that are able to adjust osmotically usually have high RWC (Bunce, 1986), high photosynthetic rates (Gupta and Berkowitz, 1987), and high yield under stress conditions (Verma and Mathur, 1988). Low temperature of 15/10°C appeared to benefit plants by allowing greater adjustment to osmotic stress from PEG, where high temperature probably acted in opposition. An indirect effect of high temperature hat minimize adverse effect of osmotic stress on photosynthesis in wheat is very evident. Plants maintained high photosynthetic rate, stomatal conductivity and grain yield under low temperature and osmotic stress b ut had low values of all factors under high temperature and osmotic stress (Shah and Paulsen, 1999). The water potential and RWC measurements under the same condi:ions suggested that low temperature enabled plants to adjust osmotically. Continued production of photosynthate at low temperature apparently provided solute for osmotic adjustment as well as assimilates for growth (Morgan, 1980; Munns, 1988). High temperature on the other hand would decrease photosynthate production by damaging photosystem-II (Al-Khatib and Paulsen, 1989; Harding et al., 1990a, b) and assimilates for either osmotic adjustment or growth would be diminished further by high respiratory loss (Gusta and Chen, 1987).
Temperature probably influences plant responses to osmotic stress in large part by determining the ability for osmotic adjustment. Two major implications for producing crops in unfavorable environment are noted. First, favorable temperature for photosynthesis can increase crop survival and yield under drought conditions by promoting osmotic adjustment. Second increasing genetic resistance of crops to high temperature by breeding also should augment osmotic adjustment and, thus, adaptation to arid conditions. The latter benefit is particularly important because high temperature and drought frequently occur together and interact to exacerbate injury to plant growth and most water relations parameters.
REFERENCES
- Al-Khatib, K. and G.M. Paulsen, 1989. Enhancement of thermal injury to photosynthesis in wheat plants and thylakoids by high light intensity. Plant Physiol., 90: 1041-1048.
CrossRefDirect Link - Angus, J.F., S. Hasegawa, T.C. Hsiao, S.P. Liboon and H.G. Zandstra, 1983. The water balance of post-monsoonal dryland crops. J. Agric. Sci., 101: 699-710.
Direct Link - Bunce, J.A., 1986. Volume and osmotic potential changes in relation to inhibition of photosynthesis by water stress in intact leaves. Can. J. Bot., 64: 557-560.
CrossRefDirect Link - Gates, D.M., 1968. Transpiration and leaf temperature. Ann. Rev. Plant Physiol., 19: 211-238.
CrossRefDirect Link - Gupta, A.S. and G.A. Berkowitz, 1987. Osmotic adjustment, symplast volume and nonstomatally mediated water stress inhibition of photosynthesis in wheat. Plant Physiol., 85: 1040-1047.
CrossRefDirect Link - Hamblin, A., D. Tennant and M.W. Perry, 1987. Management of soil water for wheat production in Western Australia. Soil Use Manage., 2: 63-69.
CrossRefDirect Link - Harding, S.A., J.A. Guikema and G.M. Paulsen, 1990. Photosynthetic decline from high temperature stress during maturation of wheat: I. Interaction with senescence processes. Plant Physiol., 92: 648-653.
PubMedDirect Link - Harding, S.A., J.A. Guikema and G.M. Paulsen, 1990. Photosynthetic decline from high temperature stress during maturation of wheat II. Interaction with source and sink processes. Plant Physiol., 92: 654-658.
CrossRefDirect Link - Johnson, R.C., H.T. Nguyen and L.I. Croy, 1984. Osmotic adjustment and solute accumulation in two wheat genotypes differing in drought resistance. Crop Sci., 24: 957-962.
CrossRefDirect Link - Kirkham, M.B., 1989. Growth and water relations of two wheat cultivars grown separately and together. Biol. Agric. Hortic. Int. J. Sustain. Prod. Syst., 6: 35-46.
CrossRefDirect Link - Munns, R., 1988. Why measure osmotic adjustment? Aust. J. Plant Physiol., 15: 717-726.
CrossRefDirect Link - Nicolas, M.E., R.M. Gleadow and M.J. Dalling, 1984. Effects of drought and high temperature on grain growth in wheat. Aust. J. Plant Physiol., 11: 553-566.
Direct Link - Nus, J.L. and C.F. Hodges, 1985. Effect of water stress and infection by Ustilago striiformis or Urocystis agropyri on leaf turgor and water potentials of Kentucky bluegrass. Crop Sci., 25: 322-326.
CrossRefDirect Link - Rhodes, P.R. and K. Matsuda, 1976. Water stress, rapid polyribosome reductions and growth. Plant Physiol., 58: 631-635.
CrossRefDirect Link - Schonfeld, M.A., R.C. Johnson, B.F. Carver and D.W. Mornhinweg, 1988. Water relations in winter wheat as drought resistance indicators. Crop Sci., 28: 526-531.
CrossRefDirect Link - Shah, N.H. and G.M. Paulsen, 1999. Response of wheat to combined high temperature and osmotic stresses during maturation. Plant photosynthesis and productivity. Pak. J. Biol. Sci., 2: 1286-1291.
CrossRefDirect Link - Shimshi, D., M.L. Mayoral and D. Atsmon, 1982. Responses to water stress in wheat and related wild species. Crop Sci., 22: 123-128.
CrossRefDirect Link - Sinclair, T.R. and M.M. Ludlow, 1985. Who taught plants thermodynamics? The unfulfilled potential of plant water potential. Aust. J. Plant Physiol., 12: 213-217.
Direct Link - Verma, H.R. and R.K. Mathur, 1988. Physiological performance of wheat (Triticum aestivum dwarf.) under irrigated situation. J. Agron. Crop Sci., 160: 145-150.
CrossRefDirect Link - Wardlaw, I.F., I.A. Dawson, P. Munibi and R. Fewster, 1989. The tolerance of wheat to high temperatures during reproductive growth. I. Survey procedures and general response patterns. Aust. J. Agric. Res., 40: 1-13.
CrossRefDirect Link