
Zakirullah
Michigan State University, USA
Zahoor A. Swati
NWFP Agricultural University, Peshawar
Anwaar Ahmad
CEMB University of Punjab, Lahore, Pakistan
Raziuddin
NWFP Agricultural University, Peshawar
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 1 | Page No.: 130-132







ABSTRACT
Drought tolerance potential of five F10 brassica lines derived from interspecific crosses involving three genotypes of B. napus, two of B. juncea and one of B. carinata was assessed under green house conditions during crop season 1992-93. Relative leaf water contents (RLWC) were significantly altered by the three levels of moisture stress at flowering stage but genotypic differences were significant at the pod formation stage only. Leaf water potential (LWP) decreased with increasing stress and increased with age from flowering to pod stage. Under stress conditions plants generally took longer to complete 100% flowering, however, maturity was not significantly delayed. Brassica juncea was the earliest in reaching maturity followed by B. carinata and B. napus, respectively, while lines showed little differences. Leaf area decreased significantly with moisture stress, Whereas genotypic differences were significant for pod length, number of pods/main stock and number of secondary branches and non-significant for number of seed/pod and grain yield/plant. The Lines AY and WY derived from crosses between Altex X Yellow Raya and Westar x Yellow Raya, respectively were proved to be stress tolerant.
PDF Abstract XML References Citation
How to cite this article
Zakirullah, Zahoor A. Swati, Anwaar Ahmad and Raziuddin, 2000. Morpho-physiological Response of Selected Brassica Lines to Moisture Stress. Pakistan Journal of Biological Sciences, 3: 130-132.
DOI: 10.3923/pjbs.2000.130.132
URL: https://scialert.net/abstract/?doi=pjbs.2000.130.132
DOI: 10.3923/pjbs.2000.130.132
URL: https://scialert.net/abstract/?doi=pjbs.2000.130.132
INTRODUCTION
With the development of double zero cultivars, Brassica is rapidly becoming a major oilseed crop of the world. However a review of the literature shows that its drought resistance potential has yet to be exploited fully. The various species of Brassica currently under cultivation possess drought resistance/tolerance to a variable degree. Scientists working on the drought tolerance potential of Brassica have drawn different conclusions from their experiments. Rapaport and Fereres (1983) while observing little differences in drought tolerance ability of various Brassiba species grown in green house has reported Brassica carinata as the most drought tolerant and Brassica juncea as the least but Ashraf and Mehmood (1990) under similar growth conditions observed that Brassica napus was the most drought tolerant, Brassica juncea and Brassica campestris intermediate and Brassica carinata as the most sensitive to drought stress. Within the same species genotypes genotypes show considerable differences in the degree of drought tolerance (Ali et al. 1988, Thukral et al. 1985). Siag et al. (1993) reported almost fifty percent reduction in the yield of B. carnpestris varieties under unirrigated condition as compared to irrigated. In this study five selected Brassica lines derived from interspecific crosses were evaluated for moisture stress using a number of morphological and physiological indices under screen house conditions.
MATERIALS AND METHODS
Five F8 through F9 selected F10 lines of Brassica derived from interspecific crosses viz Altex X P53-48-2 (AP), Wester X LL-84 (WL), LL-84 X Kivi Salem (LS), Altex X Yellow Raya (AY), and Westar X Yellow Raya (WY) belonging to six genotypes of three species namely Altex, Westar, Kivi Salam (B. napus), LL-84, P53-48-2 (B. juncea) and Yellow Raya (B. carinata) were used in this study. The experiment was conducted in the green house during crop season 1992-93. Pots of medium size (external diameter 24 cm, depth 22cm) were filled with 3 Kg clay humus mixture in 1:1 ratio and then arranged in RCB design with three replications and split plot arrangement. Initially 6 to 10 seeds were sown in each pot which after germination were thinned down to 1 to 2 seedlings/pot. Each pot received 0.34 g DAP (19% Nitrogen) and 0.35 g Urea (46% N) at the time of sowing. Based on the water holding capacity of the medium, three levels of irrigation representing application of water at field capacity (1200 ml), half of the field capacity (600 ml) and one fourth of the field capacity (300 ml) were applied. Initially all the plants received equal quantity of water for a period of 45 days, till the seedlings were established, however, the two genotypes of B. juncea started flowering at this stage. Afterwards water supply was controlled as per above specifications and the plants were watered after every fifteen days interval. Relative Leaf Water Content (RLWC) was collected at flowering and pod formation (40% pods) stages by using the following formula (Stocker, 1929):
Six discs (6mm) were taken from three fully expanded leaves (two discs) of each plant for this purpose. Leaf Water Potential was taken at three stages of plant development: a. vegetative stage (42 days after sowing), b. flowering stage (when the plants started flowering) and c. pod formation stage (when 30-40% pods formed) using dew point microvoltmeter (WESCOR model HR-33T with C-52 sample chamber), Number of days to 100 percent flowering, Leaf area at 100 percent flowering stage by the formula (Rao and Dao, 1978):
A = aBbCc
Where a, b and c are constants with values 0.9817, 1.1270, and 0.7503, respectively and A = Area, B = Breadth, L = Length. Four and fifth leaves from the top of the main stalk were selected for calculating leaf area. Pod length was taken by randomly selecting six pods/plant at maturity and then averaging the values. Number of pods/main stalk, Number of secondary branching, Number of days to maturity, Number of seeds/plant and Grain yield/plant.
RESULTS AND DISCUSSION
Relative Leaf Water Content (RLWC) increased with increasing water stress but decreased with the age from flowering to pod formation stage whereas leaf water potential (LWP) decreased with reducing water supply but increased with age during the same period, however, there were certain exceptions (Table 1). At the flowering stage, increase in RLWC was higher for Kivi Salam and LL-84, however, both maintained fairly constant values at the pod stage under the three water regimes. Selected line LS which also had a high RLWC at flowering showed a considerable decline during pod formation at which stage Yellow Raya (B. carinata) showed a reasonable increase in RLWC with increasing moisture stress. Among the lines, it was observed that LS and WL inherited the trait well from their parents while AP and AY which earlier (flowering stage) exhibited lower values compared to their parents were able to catch up with them at pod stage. Altex, their common parent also showed fluctuating response to different levels of moisture stress, it also exhibited a sharp decline in LWP values both at flowering and pod formation stages. Lines AP and AY also showed a marked decrease in LWP at the pod stage. Unlike the parents, LS maintained lower LWP under stress. The WL, which had a lower value compared to both of its parents at flowering, maintained the highest LWP under conditions of maximum stress at the pod stage.
The number of days required to reach 100 percent flowering were significantly influenced by drought and the forces of interaction also came into prominence. With certain exception the genotypes generally took longer to reach this mark but the delay was not more than 5 days for any genotype (Table 1). Maturity was not significantly delayed by water stress, however, the genotypic differences were significant and B. jucea was the earliest followed by B. carinata and B. napus, respectively. Among the lines, differences were less than a week. As the genotypes took longer to reach 100 percent flowering under conditions of water stress but maturity was not altered significantly, it can be concluded that the interval between 100 percent flowering and maturity was reduced by drought. For pod length, only genotypic differences were significant and water stress had no effect of statistical significance on it. The mean pod length was high for WL despite the fact that the character was not the best among its parents, which could be attributed to a new combination of some positive genes. Between WY and AY, both having Yellow Raya as a common parent, the latter had a higher mean pod length under all the three water regimes. Altex also showed a higher mean value compared to Westar, indicating that the character is controlled by more dominant genes in Altex and has a good transmissibility.
Leaf area was found to be very sensitive to water availability. Reduction in leaf area was observed in all the genotypes (Table 1) but was more than 60 percent for Westar, 73 percent for altex, 62 percent for Yellow Raya and 66 percent for AY under severe stress (T3). Brassica juncea responded with relatively less reduction, mainly because of its early maturity and little exposure to drought. Hunter (1980) and Younis et al. (1988) have reported similar results in corn, Shalaby et al. (1988) in wheat Khan and Agrawal (1988) in B. juncea. Yellow Raya and WY showed the highest and almost identical mean values, but AY was no way near to them. Line LS also had a high mean at T1, but under severe stress its value was close to Kivi Salam. Both AP and AY, like Altex showed significant reductions in leaf area at T3. Reductions in the number of secondary branches and pods per main stalk under the influence of drought were no statistical significance. Only WY and AY showed marked reductions in the number of secondary branches. Five of the genotypes showed an increase in the number of pods per main stalk at T3 and these included WY and AY as well. Thukral et al. (1985) has also reported reduction in the number of secondary branches and the number of siliqua/plant in only a few of the several genotypes of B. juncea, B. napus, B. carinata and B. tournifortii. Genotypic differences were, however, significant for both the characters and it was generally observed that genotypes having more number of pods on the main stalk had less number of secondary branches. Brassica napus had the highest number of pods on the main stalk followed by B. juncea and B. carinata, respectively, while for number of secondary branches it was in the reverse order. The LS, however, showed a high number of secondary branches and evenly good number of pods/main stalk.
Neither treatment effects nor genotypic differences were significant for number of seeds/pod and grain yield/plant. Interaction of genotypes with the three moisture regimes for both the characters was of no statistical significance as well. A reduction in the number of seeds/pod and grain yieldiplant under the influence of drought was, however, observed. Compared to T1, only five of the genotypes showed reduction of more than 30 percent in the number of seeds/pod at T3 while for grain yield the reduction ranged from 29 to 80 percent, except in the case of WL which showed an unusual increase of 100 percent, Among the five lines, LS had the smallest number of seeds/pod and the poorest yield at T3 while AP and AY exhibited high values over their parents for both the characters.
| Table 1: | Percent increase or decrease at T3 over T1 (T3-T1/T1) |
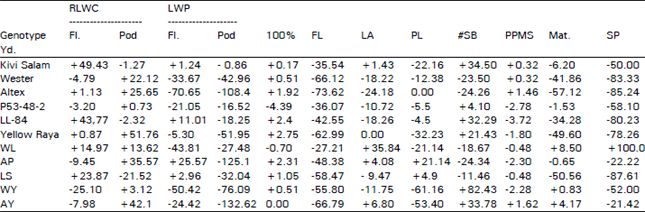 | |
| RLWC = Relative Leaf Water Content Fl = Flowering. LWP = Leaf Water Pot Length #SB = Number of Secondary Branches. PPMS = Pods Per Main Stalk Mat Yd. = Yield | |
Reduction in the yield and yield components under drought conditions is a universal phenomenon and has been observed by several workers in different crops. For instance Mailer and Cornish (1987) reported a decrease in yield and 1000 grain weight of B, napus and B. rapa, Thukral et al. (1985) in B. juncea, B. napus, B. Cannata and B. tourifortii, Keim and Kronstad (1981) in wheat, Wright et al. (1983) in sorghum and Downey (1971) in corn.
REFERENCES
- Ashraf, M. and S. Mehmood, 1990. Response of four Brassica species to drought stress. Environ. Exp. Bot., 30: 93-100.
CrossRefDirect Link - Hunter, R.B., 1980. Increased leaf area (Source) and yield of maize in short-season areas. Crop Sci., 20: 571-574.
CrossRefDirect Link - Keim, D.L. and W.E. Kronstad, 1981. Drought response of winter wheat cultivars grown under field stress conditions. Crop Sci., 21: 11-15.
CrossRef - Downey, L.A., 1971. Effect of gypsum and drought stress on maize (Zea mays L.). I. growth, light absorption and yield. Agron. J., 63: 569-572.
CrossRefDirect Link - Mailer, R.J. and P.S. Cornish, 1987. Effects of water stress on glucosinolate and oil concentrations in the seed of rape (Brassica napus L.) and turnip rape (B. rapa L. var. silvestris). Aust. J. Exp. Agric., 27: 707-711.
PubMed - Rao, S.C. and T.H. Dao, 1987. Soil water effects on low-temperature seedling emergence of five Brassica cultivars. Agron. J., 79: 517-519.
CrossRef - Wright, G.C., R.C.G. Smith and J.R. McWilliam, 1983. Differences between two grain sorghum genotypes in adaptation to drought stress. I. Crop growth and yield responses. Crop Pasture Sci., 34: 615-626.
CrossRefDirect Link