
Muhammad Jurial Baloch
Cotton Research Institute, Sakrand, District Nawabshah Sindh, Pakistan
Abdul Rahim Lakho
Cotton Research Institute, Sakrand, District Nawabshah Sindh, Pakistan
Hidayatullah Bhutto
Cotton Research Institute, Sakrand, District Nawabshah Sindh, Pakistan
Ali Mohammad Memon
Cotton Research Institute, Sakrand, District Nawabshah Sindh, Pakistan
Ghulam Nabi Panhwar
Cotton Research Institute, Sakrand, District Nawabshah Sindh, Pakistan
Abdul Hameed Soomro
Cotton Research Institute, Sakrand, District Nawabshah Sindh, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 7 | Page No.: 1183-1186







ABSTRACT
Two commercially grown cotton varieties and four advanced strains were crossed in 6×6 diallel mating design, involving F1s only. Study was made on the estimates of general and specific combining ability of the parents and also genetic components of variances for seed cotton yield, lint%, staple length and uniformity ratio. The per se hybrid performance generally revealed that parents Karishma and CRIS-134 formed the best hybrids for yield, lint% and uniformity ratio. It is also assumed that parents that perform well in per se hybrids will also perform similar for specific combining ability (sca) but this prediction did not hold true without few exceptions. The discrepancy between per se hybrid and sca value is not a good indicator to predict parental performance in hybrid combinations. However, hybrids per se result generally connoted that parents Karishma and CRIS-134 form good combinations with other parents for hybrid crop production. Specific combining ability estimates further indicated that hybrids NH-26×CRIS-78 for yield, CRIS-134×NH-26 for lint%, NH-26×CRI5-122 for staple length and CRIS-134×NH-26 for uniformity ratio are greatly preferred. Based on general combining ability (gca) estimates, parents NH-26 and CRIS-78 are preferred for yield, Karishma for lint%, CRIS-129 for staple length and CRIS-134 for uniformity ratio. These parents are expected to be improved for above characters through selection in segregating generations. The proportion of variance due to gca (based on the mean squares) is higher than sca for seed cotton yield, lint% and staple length suggested that these traits are predominantly controlled by additive genes whereas variation due to sca for only uniformity ratio is significant suggesting that this trait is entirely controlled by dominant genes. The higher proportion of dominant variance (δ2D) for uniformity ratio also denoted the similar position as variation due to sca for this trait. The additive genetic variance ((δ2A) for yield, lint% and staple length were quite substantial insinuating the importance of additive genes for these traits.
PDF Abstract XML References Citation
How to cite this article
Muhammad Jurial Baloch, Abdul Rahim Lakho, Hidayatullah Bhutto, Ali Mohammad Memon, Ghulam Nabi Panhwar and Abdul Hameed Soomro, 2000. Estimates of Combining Ability and Genetic Parameters for Yield and Fibre Traits in Upland Cotton. Pakistan Journal of Biological Sciences, 3: 1183-1186.
DOI: 10.3923/pjbs.2000.1183.1186
URL: https://scialert.net/abstract/?doi=pjbs.2000.1183.1186
DOI: 10.3923/pjbs.2000.1183.1186
URL: https://scialert.net/abstract/?doi=pjbs.2000.1183.1186
INTRODUCTION
In quantitative genetic terms, it is in fact the breeding value of an individual that advocates the average performance of its progeny (Falconer, 1989). Breeding value also referred as additive effect is the property of additive genes. On the contrary, dominant genetic variance is the result of interaction of genes within the specific locus also referred as genotypic value of an individual (Falconer, 1989) is important for hybrid vigour, consequently useful for hybrid crop development. The essential idea to consider a set of crosses among the parents is to determine as to what extent the variance among the crosses is attributed statistically to additive genes and what portion is due to dominance and their interactions. In cotton breeding, making huge number of crosses to identify potential parents based on progeny performance seems a chance game. This approach of identifying desirable parents naturally is very tedious and expensive in the sense of labour, time and space. To make large number of crosses also require effective and vigorous selection in the self ing generations. Diallel analysis is one of the mating designs where selected parents are crossed in certain order and is used in predicting combining ability of the parents. The gca, therefore designates additive genes and is used to determine the performance of parents in general terms whereas sca effect is determined by dominant genes and is used to designate the hybrid performance in specific terms. The objective of this study was to identify potential parents for hybridization and selection programmes. Knowing the type of gene actions through gca, sca and genetic variance will also help cotton breeders to formulate an effective breeding strategy to improve various cotton characters.
Materials and Methods
Six parents that included two commercially grown cotton varieties from Punjab (Karishma and NH-26) and four advanced strains of Sindh (CRIS-I29, CRIS-78, CRIS-122 and CRIS-134), developed at Cotton Research Institute, Sakrand, Pakistan were crossed in a diallel mating design. The 6×6 dialled crosses which excluded parents and reciprocals, thus formed 15 cross combinations. The crosses were made in large numbers so as to produce enough F1 seed to be grown in replicated progeny test during 1998. The F1 hybrid seeds of all 15 crosses were planted in a Randomized Complete Block Design arranged in Four replications of four rows each and 40.0 long. To avoid loss of valuable F1 seed, instead of drilling, dibbling with two to three seeds per hole was preferred. Where two to three seedlings germinated were later thinned leaving only one vigorously growing plant per hill with a distance of 2.5' row to row and 9.0" plant to plant. Normal inputs such as fertilizer, insecticide and irrigations were applied whenever required. For taking the observations, twenty five F1 plants from each progeny in each replicate were chosen at random and tagged as index plants.
The data on four quantitative traits such as seed cotton yield per plant weighed in grams, lint% calculated as the ratio of lint from seed cotton sample, staple length (fibre length at 50% span length) measured in millimeters and uniformity ratio calculated from 2.5 and 50% span length. The estimates of general combining ability (gca) and specific combining ability (sca) and their variances were determined by adopting of Griffing (1956) statistical method-4 used by Singh and Chaudhary (1979) where parents and reciprocals were excluded from the experimental material. The analysis of variance for gca and sca was also calculated through method-4 so as to ascertain the relative importance of additive and dominant gene actions functioning for characters under study.
RESULTS AND DISCUSSION
Cotton breeders are well known of the phenomenon that certain crosses form better combinations than the others in transmitting favourable alleles to their offspring. Mating designs that provide appropriate information regarding the potential of parents in both hybrid combinations and segregating generations therefore are very useful to the plant breeders. Diallel analysis as proposed by Spraguei and Tatum (1942) is one of such mating designs that distinguishes the ability of the parents to contribute favourable alleles to the progeny.
The mean squares of the crosses (hybrids) for yield, lint%, staple length and uniformity ratio were highly significant that allowed to arbitrate the proportions of genetic variability due general combining ability (gca) and specific combining ability (sca) and also the importance of their effects on each hybrid.
The per se hybrid performance of 15 cross combinations for all the characters is presented in Table 1. For seed cotton yield, hybrid CRIS-134×CRIS-129 produced the maximum yield per plant and the next in the rank was Karishma×CRIS-134. However, without few exceptions, Karishma and CRIS-134 when crossed with other four parents, gave higher yields than the other parents in hybrid combinations. Regarding per se hybrid performance for lint%, Karishma with CRIS-78 produced highest lint% of 38.20 and next in the order was Karishma×CRIS-122. In staple length, NH-28 with CRIS-122 developed the longest fibre of 29.75 mm whereas the next ranking was CRIS-129×CRIS-122. The maximum uniform fibre of 66.10% was recorded by Karishma×NH-28, however, the second high scoring hybrid was CRIS-134 with NH-26. The hybrids' performance per se thus suggested that for yield, parents Karishrna. CRIS-134 and CRIS-129; and for tint%, parents Karishma, CRIS-122 and CRIS-78; and for staple length, CRIS-122, CRIS-129 and CRIS-78 are parents of the choice. These results further suggested that selection of parents reckon on the preference of the character as none of the parents is simultaneously desirable for all the four traits except Karishma which is worthwhile for yield, lint% and uniformity ratio, based on per se hybrid performance.
The mean squares of the crosses (Table 2) for all the four traits were significant that allowed to further partition this factor into the variation due to gca and sea. For seed cotton yield, lint% and staple length, the proportions of variation due gca is higher than sca, suggesting the importance of additive genes functioning for these traits. Nonetheless, genetic variability due sca for seed cotton yield and lint% were also significant, thus further indicating that dominant genes were also responsible for these characters. Ubaidullah and Hussain (1997) also reported the importance of both additive and dominant genes for yield and lint%. For other characters, staple length and uniformity ratio either of the gene action was predominant where for staple length, only gca was significant and for uniformity ratio, sca was important. Considerable portion of additive and dominant genetic variability (Table 2) for seed cotton yield and lint% however suggested that both the
| Table 1: | Performance of F1 hybrids for yield and fibre characters in Upland cotton |
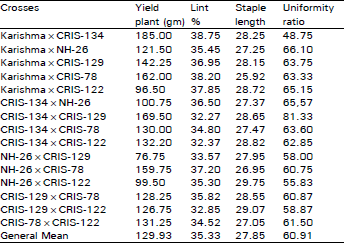 | |
traits could be improved through hybridization and imposing effective selection in segregating generations and have potential for hybrid crop production also. However, staple length can only be improved through selection whereas uniformity ratio by exploring hybrid vigour. Our results are very much in accordance with the results of Baloch et al. (1993, 1995, 1997, 1999). With our courage, we have also attempted to determine the genetic parameters such as additive (δ2A) and dominance (δ2D) variances, through the assumptions of no maternal effect, regular diploid segregation, no multiple alleles, random distribution of genes, no epistasis and no linkage effects that all may not have been met because of fewer selected parents under study. The trend regarding the importance of additive and dominance variances as indicated by the genetic variability due to ace for all the traits are very similar except the proportion of δ2D is greater than δ2A which is vice-versa in case of variability due to ace for yield only. These genetic components of variances further suggested that yield is controlled by both dominant and additive genes, lint and staple length by additive genes whereas uniformity ratio entirely by dominant genes hence completely agreeing with genetic variability due sea in the analysis of variance Table 2. Ubaidullah and Hussain (1997) however did not find significant genetic variances for any of the traits under our study.
Plant breeders generally predict that parents that perform better in hybrids per se will also perform similar for gca and definitely for sca effects. However, this type of prediction did not always remain true as reported by Srinvivasan and Gururajan (1973) and Baloch et al. (1993, 1995, 1997). The results for gca estimates are presented in Table 3. For seed cotton yield, parent NH-26 scored the maximum, 22.25 gca and next in the rank was CRIS-78 (15.40). Regarding lint%, Karishma scored the highest and for staple length, CRIS-129 was among the top scoring parent. In general, parent CRIS-78 was the good general combiner simultaneously for yield, lint% and uniformity ratio thus, suggesting that this parent retains more additive genes than the others, therefore is more favorable for developing segregating population for selection.
| Table 2: | Genetic analysis of yield and fibre characters in upland cotton |
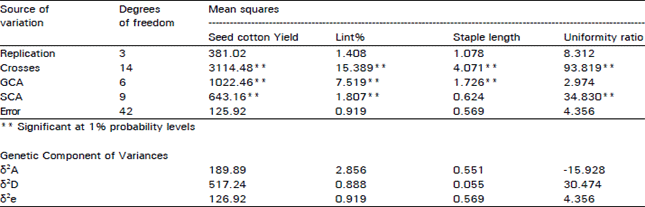 | |
| Table 3: | General combining ability estimates of yield and fibre characters in upland cotton |
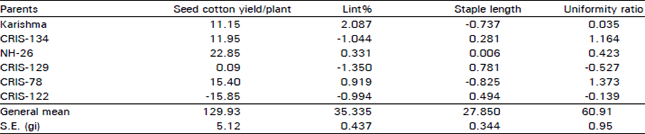 | |
| Table 4: | Specific combining ability estimates of yield and fibre traits in upland Cotton. |
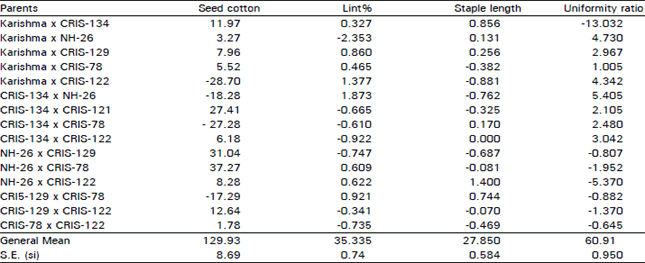 | |
In per se hybrid performance (Table 1), parents karishma and CRIS-134 were better parents for yield in particular and other traits in general. It is nevertheless expected that per se hybrid performance of parents is correlated with at least sca. This prediction did not always hold true in our case as parents Karishma and CRIS-134 which almost always formed good hybrid combinations per se but their sca effects remained generally poor without few exceptions. The sca estimates of NH-26 x CRIS-78 and CRIS- 134 x CRIS-122 were the first and second scoring hybrids respectively for yield, CRIS-134 with NH-26 was the highest scoring for lint%, NH-26 x CRIS-122, the highest scoring for staple length whereas CRIS-134 x NH-26 and Karishma x NH-26 the first and next maximum scoring specific combiner hybrids respectively for uniformity ratio. This discrepancy between per se hybrid performance and sca estimates is therefore not unusual.
Our results are very coinciding with the observations obtained by Baloch et al. (1997) who noted that some of the parents perform equally better either in hybrids per se or for sca. These findings are also in consonance with the results of Soomro (1989) and Baloch et al. (1993, 1995). Specific combining ability estimates thus suggested that dominant gene action is very obvious in the parents for yield and uniformity ratio however, less important for lint% and staple length because without one exception in both later traits, sca estimates did not reach even a unit value. Baloch et al. (1997, 1999) also noted less importance of lint% and staple length based on gca and sca estimates (Table 4).
REFERENCES
- Baloch, M.J., H. Bhutto, R. Rind and G.H. Tunio, 1995. Combining ability estimates in 5x5 diallel intrahirsutum crosses. Pak. J. Bot., 27: 121-126.
Direct Link - Baloch, M.J., H. Bhutto and A.R. Lakho, 1997. Combining ability estimates of highly adapted tester lines crossed with pollinator in breeds of cotton. Pak. J. Sci. Ind. Res., 40: 95-98.
Direct Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link