
Rufus. S. A. Gill
Department of Botany, University of Karachi-75270, Pakistan
S. Shahid Shaukat
Department of Botany, University of Karachi-75270, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 1 | Page No.: 114-117







ABSTRACT
The Fungicide Captan was screened in Allium cepa multiple assay test for mutagenic activity. Actively dividing root tips were treated with four different concectrations 5, 10, 20 and 40 ppm of the fungicide for 1, 3, 6, 12 and 24 hours. Almost all trearments exerted a mitodepressive action indicating that the fungicide has an immediate effect on cell division . It was noted that the fungicide exerts a clastogenic effect on chromosomes. Mitotic aberrations observed included stickiness of chromosomes at prophase, metaphase, bridging at anaphase and bi and tri nucleated cells. Such aberrations are important as they may have evolutionary significance.
PDF Abstract XML References Citation
How to cite this article
Rufus. S. A. Gill and S. Shahid Shaukat, 2000. Genotoxic Effects of Captan Fungicide on Root Meristems of Allium cepa L. In vivo. Pakistan Journal of Biological Sciences, 3: 114-117.
DOI: 10.3923/pjbs.2000.114.117
URL: https://scialert.net/abstract/?doi=pjbs.2000.114.117
DOI: 10.3923/pjbs.2000.114.117
URL: https://scialert.net/abstract/?doi=pjbs.2000.114.117
INTRODUCTION
Agro-chemicals have become an integral part of modern agriculture and annually several million tons of organic and inorganic chemicals with antimicrobial and insecticidal properties are used. Some of these while inhibiting pathogens not only upset the ecosystem but also induce undesirable changes in higher organisms (Jain and Sarbhoy, 1988). Quite a large number of compounds have been identified for clastogenic, mutagenic, carcinogenic and teratogenic effects. Compounds producing chromosomal abnormalities usually also induce genetic damage as well (Sugimura et al., 1982). Studies have showen that indiscriminate use of Agro-chemicals results in many undesirable secondary consequences on higher plants (Ferrin and Rohde, 1992a,b). The present study was carried out to evaluate the effect of fungicide Captan on the dividing cells of the root tips of onion (Allium cepa L.).
MATERIALS AND METHODS
Bulbs of a diploid (2n = 16) onion (Allium cepa L. var. elope) obtained locally were used as experimental material, Clean onion bulbs having their basal portion shaved were placed on 80 ml glass beakers containing distilled water for four days at 25-28°C (day/night) with 14 hours day length in a growth chamber to obtain roots of approximately 4-5 cm length. These roots were then exposed to Captan, fungicide at concentrations of 5,10, 20 and 40 ppm for 1, 3, 6, 12 and 24 hours. Controls were kept in distilled water. The experimental set up was completely randomized design with eight replications (Table 1). Six root tips from each replicate bulb were fixed in freshly prepared acetic-ethanol (1:3) fixative in separate vials. Squash preparations were made using 2,0 percent aceto-orcein (Pandita, 1983). For each replicate about 1000-1500 cells from 6 roots were examined and analyzed with respect to mitosis and chromosomal aberrations. For each treatment mitotic index was calculated as number of dividing cells as a percentage of total number of cells. Chromosomal abnormalities were scored in pro, meta, ana and telophase stages and expressed as percentage of the total dividing cells. The data on mitotic index was subjected to analysis of variance (ANOVA). Before performing ANOVA the percentage data was transformed by an arcsin transformation (Sakai and Rholf, 1995).
RESULTS AND DISCUSSION
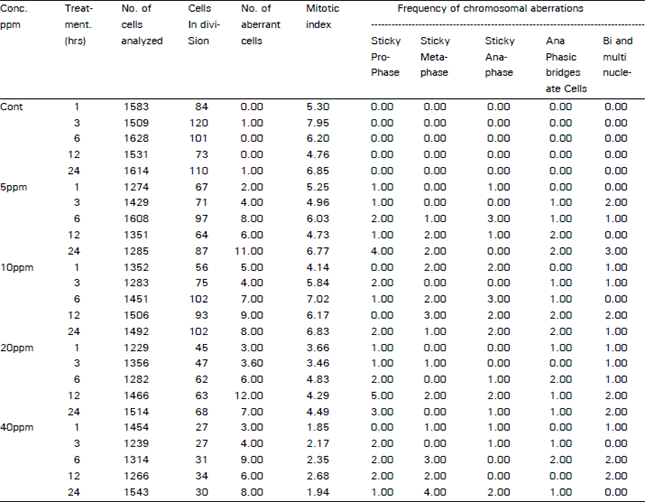
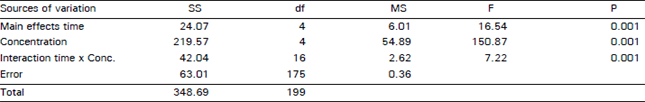
Captan treated cells showed a reduction in the mitotic index at all doses and treatment durations when compared to the control sets. Results of ANOVA revealed that there was a significant effect of fungicide concentration (F = 150.87, p<0.001) while time of exposure was also highly significant (F=16.54, p<0.0) (Table 2). The divisional frequency was found to be reduced even at 1 hour exposure period in all the treatments indicating that the fungicide has an immediate effect on cell division. This provides evidence on the mitotoxicity of the test chemical. With the increase in concentration mitotic index decreased gradually in all the treatments. Results indicating decrease in mitotic index with increasing chemical concentration have also been reported by many workers (Tomkins and Grant 1972; Amer and Farah, 1985; Badr, 1988; Rabindra et al., 1989). Schneiderman et al. (1971) have suggested that the reduction in the mitotic activity could be due to inhibition of DNA synthesis.
The chromosomal aberrations induced by the fungicide Captan in the present study included disturbed prophase, disturbed and sticky metaphase, stickiness and anaphasic bridges, bi and trinucleated cells. Chrosomal stickiness was observed at all concentration of the fungicide though concentration itself seems to have no effect (Fig. 1). However, with increasing exposure time to the fungicide stickiness at prophase and metaphase was found to increase.
 | |
| Fig. 1: | Mitotic abnormalities found in dividing cells of Allium cepa resulting from Captan fungicide treatment (A) distubed metaphase. (B) Stickiness at metaphase.(C,D,E) stickiness at anaphase. (F) stickiness and bridging at anaphase. (G <H) bridging at anaphase. (1) Si tri-nucleated cells. |
| Table 1: | Mitotic index, percentage of normal and abnormal mitotic cell is out of observed number of dividing root tip cells of Allium cepa with the fungicide Captan in various concentrations for different durations. |
 | |
| Table 2: | Analysis of variance for the data of mitotic index of meristematic root tip cells of Allium cepa exposed to Captan at different concentrations and durations. |
 | |
Stickiness results when chromosomes do not unfold correctly and the chromatids become connected due to bridging between DNA fibers (Sharma, 1991). Anaphasic bridges were recorded at various concentrations of the fungicide, but their frequency did not differ with the concentration. Anaphasic bridges are produced due to chromosomal breakage and joining of incorrect ends (Ahmad and Yasmin, 1992). Anaphasic bridges generally appeared after 3 hours exposure to the fungicide.Bi- and trinucleated cells increased with increasing concentration. These cells arise due to the impaired function of the Golgi vesicles. These vesicles are the source of the carbohydrate that form the cell plate and eventually the cell wall (Sheeler and Bianchi, 1983). Further mitotic division produces aneuploidy as demonstrated by Chauhan and Suanderaman (1990). Similar chromosomal aberrations are also known to be induced by herbicides, chemical mutagens and ionizing radiations. Such aberrations are important as they may often have evolutionary significance.
REFERENCES
- Ahmad, S. and R. Yasmin, 1992. Effects of methyl parathion and Tri-miltox on the mitosis of Allium cepa. Cytologia, 57: 155-160.
Direct Link - Jain, A.K. and R.K. Sarbhoy, 1988. Cytogenetical studies on the effects of some chlorinated pesticides III. Concluding remarks. Cytologia, 53: 427-436.
CrossRefDirect Link - Amer, S.M. and O.R. Farah, 1985. Cytological effects of pesticides XV. Effect of the insecticide Methamidophos on root-mitosis of Vicia faba. Cytologia, 50: 521-526.
CrossRefDirect Link - Chauhan, L.K.S. and V. Sundararaman, 1990. Effect of substituted ureas on plant cells I. Cytological effects of isoproturon on the root meristem cells of Allium cepa. Cytologia, 55: 91-98.
CrossRefDirect Link - Ferrin, D.M. and R.G. Rohde, 1992. Population dynamics of Phytophthora parasitica, the cause of root and crown and rot of Catharanthus roseus, in relation to fungicide use. Plant Dis., 76: 60-63.
Direct Link - Ferrin, D.M. and R.G. Rohde, 1992. In vivo expression of resistance to metalaxyl by a nursery isolate of Phytophthora parasitica from Catharanthus roseus. Plant Dis., 76: 82-84.
Direct Link - Pandita, T.K., 1983. Mutagenic studies on the insecticide Metasystox-R with different genetic systems. Mutat. Res./Genet. Toxicol., 124: 97-102.
CrossRefDirect Link - Sinha, R.K., R. Choudhury and R. Mallick, 1989. Cytological effects of phosalone on root meristem of Allium cepa L. Cytologia, 54: 429-435.
CrossRefDirect Link - Schneiderman, M.H., W.C. Dewey and D.P. Highfield, 1971. Inhibition of DNA synthesis in synchronized Chinese hamster cells treated in G1 with cycloheximide. Exp. Cell Res., 67: 147-155.
CrossRefDirect Link - Tomkins, D.J. and W.F. Grant, 1972. Comparative cytological effects of the pesticides menazon, metrobromuron and tetrachloroisophthalonitrile in Hordeum and Tradescantia. Can. J. Genet. Cytol., 14: 245-256.
CrossRefDirect Link