
Muhammad Tariq Saeed
Department of Plant Breeding and Genetics, University of Agriculture, Faisalabad, Pakistan
Muhammad Saleem
Department of Plant Breeding and Genetics, University of Agriculture, Faisalabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 7 | Page No.: 1138-1140







ABSTRACT
Nature of gene action was investigated for various agronomic characters such ear length, number of ears per plant, number of kernel rows per ear, number of kernels per row, 100-grain weight and grain yield per plant in a set of complete diallel crosses, involving six elite maize inbred lines (B 70, EX 22$, FR 15, N 28, PA 91, SYP 5). The Vr-Wr graphs indicated over-dominance type of gene action for number of ears per plant while additive type of gene action for all other traits. Inbred line FR 15 and PA 91 were found with possessed most dominant genes for number of ears per plant and ear length respectively. Inbred line B 70 was found to has most dominant genes for 100-grain weight, number of kernel rows per ear, number of kernels per row and grain yield per plant. Inbred line B 70 possessed most recessive genes for number of ears per plant. The five characters like ear length, number of kernel rows per ear, number of kernels per row, 100-grain weight and grain yield per plant were under the additive genetic control and mass selection can be practised for these characters, whereas number of ears per plant was non-additively controlled and selection for these populations must be practised with great care to develop pure breeding line. Thus inbred line B 70 was found to have most promising performance while making selection for higher yield.
PDF Abstract XML References Citation
How to cite this article
Muhammad Tariq Saeed and Muhammad Saleem, 2000. Estimates of Gene Effects for Some Important Quantitative Plant Traits in Maize Diallel Crosses. Pakistan Journal of Biological Sciences, 3: 1138-1140.
DOI: 10.3923/pjbs.2000.1138.1140
URL: https://scialert.net/abstract/?doi=pjbs.2000.1138.1140
DOI: 10.3923/pjbs.2000.1138.1140
URL: https://scialert.net/abstract/?doi=pjbs.2000.1138.1140
INTRODUCTION
Maize (Zea mays L.) is one of the important cereal crop in the world and occupies third position after wheat and rice with respect to area and production. It is extensively grown in temperate, sub-tropical and tropical regions of the world. Its range of cultivation stretches from 50°N to 40°S latitude and at an altitude from sea level to 3300 meters high.
Maize contributes significantly to the overall food production in the country as well as giving economic returns to the growers in a relatively lesser time. In spite of its greater yield potential, we are lagging behind the world average yield on per unit area basis and maximum potential is yet to be exploited.
It is being extensively used for the preparation of corn starch, corn oil, dextrose, corn syrup, corn flakes, gluten, lactic acid, alcohol and acetone which are used for various purposes such as paper, foundry, fermentation and food industries.
The average protein content of maize grain is generally 9-10 %. However, several improved types have been developed in U.S.A. which have 40 % protein in grain.
The primary objective in maize breeding programmes is the evaluation of high yielding and locally adapted varieties and as a result a number of hybrids and synthetics with high yield potential have been released in the past for commercial cultivation. However, breeding for improved varieties is a continuous process and requires a thorough knowledge of the genetic mechanism of yield and yield components.
Grain yield is a complex phenomenon, which results from the interaction of a host of contributing factors, which are highly influenced by the environmental variation. It cannot directly be improved by phenotypic selection of desirable plants, especially when we are dealing with heterozygous crops like maize. To overcome this tedious problem, yield is partitioned into its components and thus the possible way to increase yielding ability is to study inheritance mechanism governing various components of grain yield.
MATERIALS AND METHODS
The present research was conducted in the department of Plant Breeding and Genetics, University of Agriculture, Faisalabad during the year 1997 according to the diallel model of Hayman (1954) and Jinks (1954). The experimental material was comprised of the following six inbred lines of maize maintained at U.A.F.
| 1. | B 70 |
| 2. | EX 228 |
| 3. | FR 15 |
| 4. | N 26 |
| 5. | PA 91 |
| 6. | SYP 5 |
The inbred lines were crossed in a diallel fashion during summer 1997 to obtain all possible single and reciprocal crosses. During the next growing season (spring, 1998), seeds of all possible crosses along with the parents were sown in a triplicated trial according to a Randomized Complete Block Design. Each entry was sown in two rows of 5 m length keeping row to row distance 60cm and plant to plant distance 23 cm. Data for the following characters were recorded on ten guarded plants from each entry.
| 1. | Number of ears per plant |
| 2. | Number of kernel rows per ear |
| 3. | Number of kernels per row |
| 4. | Ear length (cm) |
| 5. | 100-grain weight (g) |
| 6. | Grain yield per plant (g) |
The data were subjected to analysis of variance and further evaluated for gene action according to Hayman (1954) and Jinks (1954).
RESULTS AND DISCUSSION
Number of ears per plant: A reference to analysis of variance for number of ears per plant showed that the differences among the genotypes were highly significant. From the graphical presentation (Fig. 1), it is evident that regression line intercepts the Wr-axis below the point of origin, indicating over-dominance type of gene action for this character. Inbred line FR 15 had most dominant genes, while inbred line B 70 had most recessive genes. Similar results have been reported by Akhtar (1971). The results, however, differ from those of Chaudhry (1974) who reported additive type of gene action for this character.
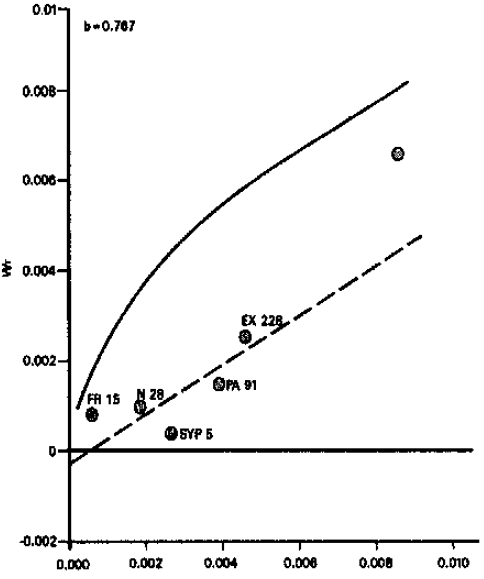 | ||
Fig. 1: | Vr/Wr graph for number of ears per plant | |
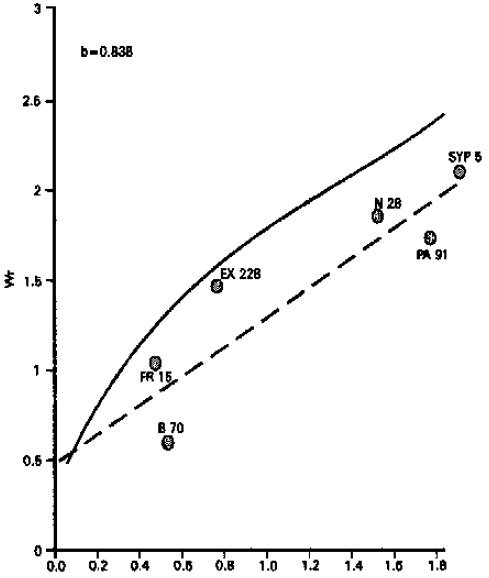 | ||
Fig. 2: | Vr/Wr graph for number of kernels rows per ear | |
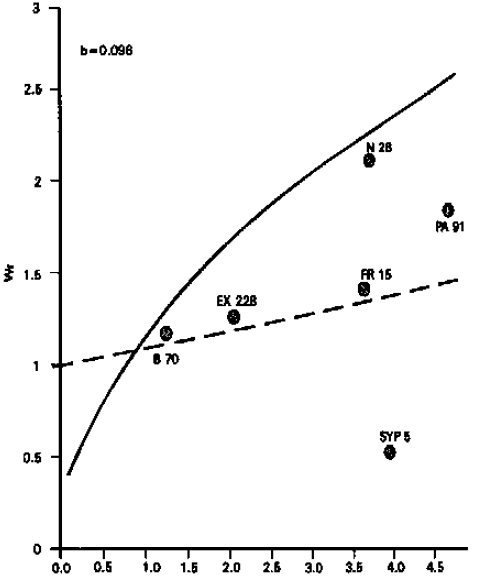 | ||
Fig. 3: | Vr/Wr graph for number of kernels per rows | |
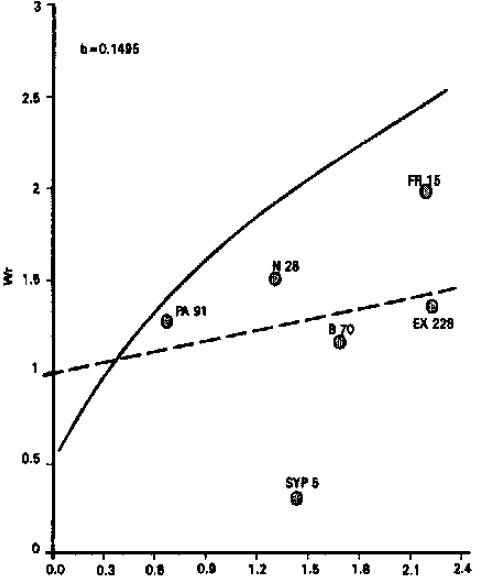 | ||
Fig. 4: | Vr/Wr graph for ear length | |
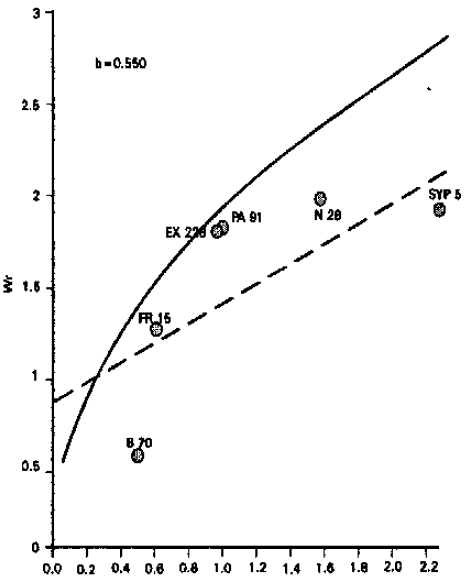 | ||
Fig. 5: | Vr/Wr graph for 100-grain weight (g) | |
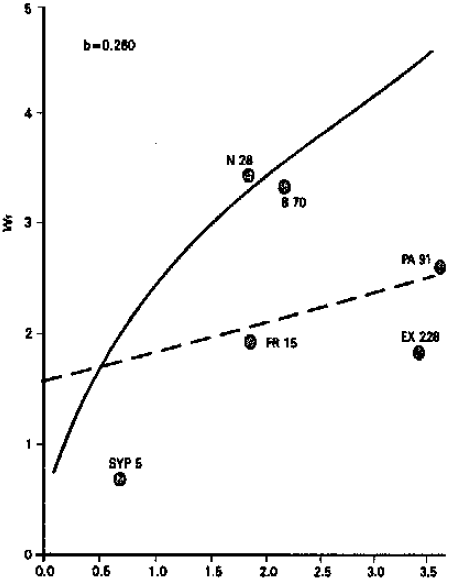 | ||
Fig. 6: | Vr/Wr graph for grain yeild per plant | |
Number of kernel rows per ear: Analysis of variance showed that the differences among the genotypes were highly significant. From the graphical presentation (Fig. 2), it is evident that the regression line passes above the point of origin which suggests the additive type of gene action for this character. Inbred line B 70 possessed maximum dominant genes, while inbred line SYP 5 carried most recessive genes. The results are in line with those of Saghir (1980). However, the results differ from those of Kanaka (1982) who reported over-dominance type of gene action.
Number of kernels per row: Analysis of variance for number of kernels per row indicated that the differences among the genotypes were significant. As the regression line passed above the point of origin (Fig. 3), which demonstrated additive type of gene action for this character. From the relative position of the array points on the regression line, it is evident that inbred line B 70 possessed most dominant genes, while inbred line PA 91 carried most recessive genes. These results are compatible with Saghir (1980) who reported additive type of gene action. The present results, however, differ from those of Akhtar (1971), Kanaka (1982) and Singh et al. (1995) who observed over-dominance type of gene action for this trait.
Ear length: Analysis of variance for ear length indicated that the differences among the genotypes were highly significant. From the graphical presentation (Fig. 4), it is evident that the regression line passes above the point of origin which suggests the additive type of gene action for this character. Inbred line PA 91 possessed most dominant genes, while inbred line FR 15 possessed most recessive genes. The present studies get support from those of Setty (1978) and Singh et al. (1995). On the other hand, Akhtar (1971) reported that ear length was controlled by non-additive type of gene action.
100-grain weight: Analysis of variance for 100-grain weight revealed that he differences among the genotypes were highly significant. From the graphical presentation (Fig. 5), it is evident that the regression line passes above the point of origin, which suggests additive type of gene action controlling this character.
Inbred line B 70 possessed maximum dominant genes, while inbred line SYP 5 carried most recessive genes. Chaudhry (1974) and Ramamurthy (1980) reported that grain weight was under the control of additive type of gene action. However, the results of the present study differ from those of Kanaka (1982) and Saghir (1980) who reported over-dominance type of gene action.
Grain yield per plant: The differences among the genotypes were highly significant. It is apparent from the graphic presentation (Fig. 6) that the regression line passes above the point of origin, which shows additive type of gene action in the inheritance of grain yield per plant. Inbred line B 70 possessed maximum dominant genes, while inbred line PA 91 carried most recessive genes. The results corroborate with the findings of Singh et al. (1995), whereas Kanaka (1982) and Saghir (1980) found that this character was controlled by over-dominance type of gene action. Predictions in the case of additive gene action would be expected to be more reliable as compared to the characters which are controlled by non-additive genes. As suggested by Verhalen et al. (1971) a preponderance of non-additive effects would not favour mass selection in altering any of the traits but pedigree test, sib test, progeny test or various combinations among them, will certainly be required to improve grain yield through various yield components. To conclude, it may be said that the overall information obtained in the present study if practised with care can, in general, go a long way in developing promising synthetics and hybrids of maize.
REFERENCES
- Jinks, J.L., 1954. The analysis of continuous variation in a diallel cross of Nicotiana rustica varieties. Genetics, 39: 767-788.
Direct Link - Verhalen, L.M., W.C. Morrison, B.A. Al-Raw, K.C. Fun and Y.J.C. Murray, 1971. A diallel analaysis of several agronomic traits in upland cotton (Gossypium hirsutum L.). Crop Sci., 11: 92-96.
CrossRefDirect Link