
Mohammad Arif
Department of Plant Pathology, Faculty of Crop Protection Sciences,
N. W. F. P. Agricultural University, Peshawar, Pakistan
Sher Hassan
Department of Plant Pathology, Faculty of Crop Protection Sciences,
N. W. F. P. Agricultural University, Peshawar, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 1 | Page No.: 1-9







ABSTRACT
Pathogen-derived resistance strategies using coat proteins, movement proteins, non-structural proteins. replicases, antisense and satellite RNAs for the production of virus-resistant transgenic plants are briefly discussed. The concept of recombination of viral RNA with transgene products is critically analyzed and potential risks associated with commercialization of transgenic plants with viral inserts and its effects on bio-safety, are highlighted.
PDF Abstract XML References Citation
How to cite this article
Mohammad Arif and Sher Hassan, 2000. Pathogen-Derived Resistance Against Plant Viruses: Postscript and Prospects. Pakistan Journal of Biological Sciences, 3: 1-9.
DOI: 10.3923/pjbs.2000.1.9
URL: https://scialert.net/abstract/?doi=pjbs.2000.1.9
DOI: 10.3923/pjbs.2000.1.9
URL: https://scialert.net/abstract/?doi=pjbs.2000.1.9
INTRODUCTION
Virus diseases cause serious losses in horticultural and agricultural crops. They are especially trouble-some in tropical and subtropical areas of developing countries where population density is high, plant disease diagnostic techniques and preventive measures are few and vectors of viruses are in abundance. Once a particular crop plant is infected by a virus, no proper and effective cure is available. Prophylactic measures are the only tools to control plant virus diseases to minimize yield and quality losses. Crop losses caused by viruses are variable and mostly depend upon host genotype, virus strains, environmental conditions and prevalent virus vectors. Crop losses in a few crops due to viruses are listed in Table 1.
For the farmers, vector control through chemical sprays or inherit resistant crop varieties are the options for controlling plant virus diseases after planting certified virus-free propagative materials (seed, cuttings, bud-wood, bulbs, etc.). But continuous spread of virus diseases through various agencies, build up of inoculum and development of severe virus strains, increase in population of viruliferous vectors (aphids, white flies, hoppers, beetles, nematodes and fungi) and gradual deterioration and degeneration of crop varieties especially those grown vegetatively, lead to unprofitable agricultural production. Conventional breeding programme is one of the option to develop resistant crop varieties but this practice is time consuming and nondurable because the resistance is quite often over come by the development of new virulent virus strains.
Transformation of plants with segments of viral genomes frequently results in the development of plants that are resistant or immune to the virus from which the sequences were derived. This new approach is generally called "pathogen-derived resistance (PDR)". Important PDR strategies used in crop protection against viruses are listed below (Table 2).
Although this technology has been successfully exploited to control virus infections in plants but an environmental risk factor is evident in this work based on propensity of viruses to interact during simultaneous replication in mixed infection, The hazards are associated with the possibility that a virus infecting a PDR-transgenic plants: (a) might be genetically altered through genomic recombination between viral RNA and mRNA of transgenic viral coat protein; (b) might be altered in transmission properties by transcapsidation with coat protein generated in the transgenic plants, thereby gaining for the virus potential for access to new plant communities to which it was previously excluded, or © might acquire a new capacity to infect the transgenic plant by virtue of function provided by expression of trangenic gene. This paper reviews briefly the information available on PDR and highlights the risks associated with commercialization of PDR-transgenic plants with virus-inserts.
Pathogen-derived resistance strategies in crop protection: During early 1980s an alternative approach emerged from various laboratories that it might be possible to engineer and incorporate stable resistance in a susceptible crop by introducing a genome segment of the pathogen into the plant genome (Hamilton, 1980; Sanford and Jhonston, 1985). The theory of PDR became reality in 1987 when Abel et al. (1986) demonstrated that tobacco (Nicotiana tabacum cv. Xanthi) plants which had been stablely transformed (by Agrobacterium tomefacins) to express the coat protein (CP) gene of tobacco mosaic virus (TMV) were resistant to the virus. Several independently transformed lines of tobacco plants were challenged by mechanical inoculation either with virion or with naked TMV RNA (Abel et al., 1986). A direct positive correlation existed between the amount of TMV CP gene expressed and the level of resistance and that the resistance was the highest against subsequent TMV infection. In most cases, the resistance was not absolute (i.e., the plants were not immune to virus), nor had the plants become tolerant (i.e., supportive of symptomless infections) but there was a delay of several days in the appearance of disease symptoms and due to slow systemic movement of the virus in the plants. This coat protein-mediated resistance (CPMR) was eventually overcame when the transgenic plants were inoculated with very high concentration 1 μg/ml) of the virus inoculum.
| Table 1: | Crop losses due to viruses (1992 values) |
 | |
| [Source: modified from Wilson and Davies (1994)] | |
| Table 2: | Pathogen-derived resistance strategies used in crop protection |
 | |
| [Source: modified from Wilson (1993) and Beachy (1997)] | |
When plants of the same tobacco line 3404 were inoculated with naked TMV RNA at 0.5-1.0 μg/ml which could easily initiated infection on susceptible controls, failed to infect CP-transgenic plants (Reimann-Philipp and Beachy, 1993) indicating a degree of resistance to TMV RNA challenge inoculation. Similar results have also been reported in 1987 using potato viruses X or S (PVX or PVS) CP transgenic plants and PVX or PVS RNA inocula (Hemenway et al., 1988; MacKenzie and Tremaine, 1990; MacKenzie et al., 1991) (Table 3). Occasionally, an antisense CP gene construct would confer some protection against the cognate virus in transgenic test plants (Hemenway et al., 1988: Cuozzo et al., 1988). In general, however, this strategy, which has been successful in down-regulating or interfering with plant gene expression (e.g., as in the Flavr SavrTM tomato of Calgene Inc.), has proved almost universally disappointing and ineffective against plant RNA viruses which replicate to very high copy numbers in the cytoplasm of infected cells. Only in the case of a single-stranded (ss)DNA gemini virus an antisense gene sequence provided significant protection against challenge virus replication, which takes place in the cell nucleus (Day et al., 1991; Bejarano and Lichtenstein, 1994).
Between 1987 and 1995, large number of reports appeared on CPMR targeted against members of all the major groups of plant RNA viruses (Table 3). These have been reviewed extensively (Beachy et al., 1990; Wilson, 1993; Lomonossoff, 1995; Beachy, 1997; Lecoq, 1997). CPMR against a ssDNA gemini virus has also been reported (Kunik et al., 1994) and work is still in progress to demonstrate CPMR against double-stranded (ds)DNA caulirnoviruses or badnaviruses. Recently, coat protein-mediated immunity (CPMI) has been reported against potato mop-top, a fungally-transmitted rod-shaped virus (PMTV) and it has been shown that CPMI completely protected CPtransformed Nicotiana benthamona plants against natural transmission of the virus by its fungal vector, Spongospora subterranea f. sp subterranea (Reavy et al., 1995; Arif et al., 1999).
A second revolutionary approach appeared in 1990 when it was shown (Golemboski et al., 1990) that transgenic plants expressing the TMV sequence encoding for non-structural, enzymatic protein, the RNA-dependent RNA polymerase or replicase, had also conferred protection against high concentration of challenge inocula of TMV (100 μg/ml) or TMV RNA (300 μg/ml). But as with TMV CP, these objectives were quickly overlooked with the serendipitous discovery of another pathogen-derived, dominant and effective resistance "the replicase gene mediated resistance". Replicase gene-mediated resistance (RGMR) operates only against very closely-related viruses, even more so than CPMR, but seemed to confer "immunity" by inhibiting all stages of the viral RNA replication cycle (Carr and Zaitlin, 1991; Carr et al., 1994).
Several parallel RGMR systems were soon developed and studied, notably trom pea early browning tobravirus (PEBV; Farlane and Davies 1992), potato virus X (PVX; Braun and Hemenway, 1992; Longstaff et al., 1993), CMV (Anderson et al., 1992; Zaitlin et al., 1994; Carr et al., 1994), the most widely prevalent potato virus Y (Audy et al., 1994) and cyrnbidium ringspot tombusvirus (Rubino et al., 1993). When a large number of independent transgenic lines expressing complete, truncated or mutated replicase protein genes were compared from different laboratories, it became clear that the resistance did not correlate with the amount of detectable protein, or in any predictable way with the specific gene construct (reviewed in Wilson (1993)). To answer or settle some of these discrepancies, subtle (and now largely redundant) arguments were made about the relative infectivity of the full-length virus clone from which the replicase gene(s) had been sub-cloned due to high RNA mutation rates and inter-strain variation.
| Table 3: | Examples of coat protein-mediated resistance against virus infection |
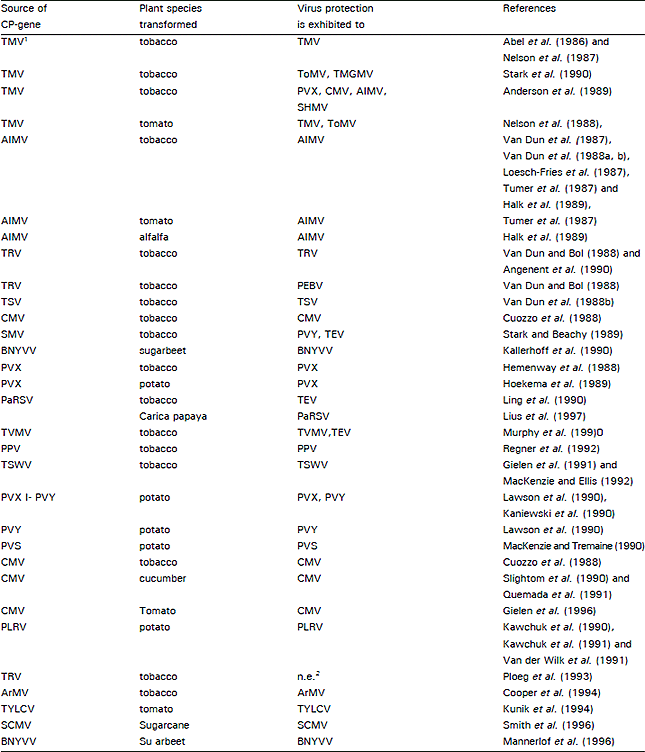 | |
1TMV-tobacco mosaic virus; ToMV-tomato mosaic virus; TMGMV-tobacco mild green mosaic virus; PVX-potato virus X; CMV-cucumber mosaic virus; AIMV-Alfalfa mosaic virus; SHMV-sun-hemp mosaic virus; TRV-tobacco rattle virus; PEBV-pea early browning virus; TSV-tobacco streak virus; SMV-soybean mosaic virus; PVY-potato virus Y; TEVtobacco etch virus; BNYVV-beet necrotic yellow vein virus; TBRV-tomato bushy stunt virus; PaRSV-papaya ringspot virus; PPV-plum pox virus; TVMV-tobacco vein mottle virus; PLRV-potato leafroll virus; TYLCV-tornato yellow leaf curl virus; ArMV-arabis mosaic virus. SCMV•sugarcane mosaic virus; 2 not effective. | |
From the studies, it became apparent that sequences other than the CP gene could function as pathogen-derived resistance genes, thus diverging researchers from strategic connection to virus assembly-disassembly events or classical "cross-protection" ideas. The rush was on functional or mutated cell-to-cell movement of protein genes, virus-coded protease genes, helicase genes, nucleotide binding protein genes and even viral genes for proteins of no known function sub-cloned and inserted into a model or in a real crop plant DNA to demonstrate protection against the virus of the source gene.
Increasingly, the hope was to find a sequence that would confer broad spectrum resistance against many plant viruses, both related and unrelated to the viral source of the gene. By and large this has not happened. These strategies have involved antibody (Tavladoraki et al., 1993) cytotoxin or inhibitor protein genes, often designed to be activated only when a uniquely viral event occurs e.g., dsRNA mediated activation of the 2,5-A (interferon-like) pathway (Truve et al., 1993). Strategies involved in the expression of wound-induced pathogenesis-related proteins, or ribsome-inactiviating proteins (Taylor et al., 1994) such as ricin, pokeweed antiviral protein (Lodge et al., 1993), or diphtheria toxin. The prospect of widespread environmental releases of crop plants expressing some (or any) of these (latent) cytotoxic genes will doubtless call for careful review and stringent risk assessments to be made. A third revolution occurred in 1992 when Dougherty and colleagues (Lindbo and Dougherty, 1992a, b) first showed that a CP gene, mutated to lose its ability of making any protein at all in plants, could, in many transformed tobacco lines, confer very high levels of resistance (immunity) to the parent virus. Taken together with data on non-expressing or low expressing lines of transformed plants with replicase genes, Baulcombe (1994), Dougherty et al. (1993), Silva-Rosales et al. (1994), Smith et al. (1994) and Prins and Goldbach (1996) independently developed theories on RNA-mediated resistance strategies and their operation through an RNA sequence-specific, host cell cytoplasmic,RNA degradation pathway; the same pathway which degrades over-expressed, unnecessary, faulty or already-translated cellular mRNAs that causes the genetic phenomena known as co-suppression.
Working with PVX and tobacco etch potyvirus (TEV), these research groups have fine-tuned the requirements for RNA-mediated transgenic resistance and have begun to dissect the generic, cellular mechanisms responsible for this process. As a current rule-of-thumb, "co-suppression" by a pathogen-derived resistance gene sequence requires at least 1 kb of 100 percent identical nucleotide sequence and up to 4kb of 80-90 percent identical sequence of RNA to be able to prime the cellular RNA degradation pathway in order to destroy incoming RNA from the challenge pathogen. A clue to identify and select those transgenic lines most likely to have effective RNA mediated resistance (in contrast to those for functional or defective viral protein-mediated resistance) is to select the lowest RNA expressors. Low expression is known to be independent of transgene copy number or T-DNA insert position effects-two popular old dogmas. It actually relates to activation of the foreign sequence-specific RNA degradation pathway.
The most elegant demonstration of this hypothesis to date comes from Baulcombe and colleagues (English et al., 1995; Mueller et al., 1995) who acquired both high and low expressing lines of tobacco plants transformed with an Escherichia coli B-glucuronidase (GUS) gene (Hobbs et al., 1993). A full-length clone of PVX with an additional inserted GUS gene sequence was used to create infectious RNA. When these PVX: GUS RNAs were mechanically inoculated onto high or low GUS expressing transgenic tobacco a wild-type PVX infection occurred only on the high GUS expressing lines; the low expressors were resistant to the virus. This PVX resistance operated at the cytoplasmic level, where PVX replicates and arose by plant cell-mediated degradation of PVX:GUS inoculum RNA at the GUS sequence, thus confirming the "co-suppression' hypothesis. Alongside these major developments in plant virus genome-derived transgenes there has been sustained interest in other virus-dependent (parasitic) RNA or DNA sequences with the capacity to ameliorate symptoms and/or to cause reduced virus replication and to increase plant tolerance, resistance or even immunity. Specifically these include satellite RNAs (Harrison et al., 1987; Gerlach et al., 1987; Ponz et al., 1987; Jacquemond et al., 1988; Roossinck et al., 1992; McGarvey et al., 1994; Kurath and Dodds, 1994), defective interfering (DI) RNAs (Hillman et al., 1987; Li et al., 1989; Burgyan et al., 1989; Jones et al., 1990; Marsh et al., 1991a, b; De Oliveira Resende et al., 1991, 1992; Yie et al., 1992; Yie and Tien, 1993; White and Morris, 1994) or DNAs (Stanley et al., 1990; Frischmuth and Stanley, 1991; Stenger,1994) and ribozyme (Uhlenbeck, 1987; Haseloff and Gerlach, 1988; Symons, 1991; Mazzolini et al., 1992; Edington et al., 1992; Steinecke et al., 1992; L'Huillier et al., 1992) sequences connected to viral antisense arm sequences (10-20 nucleotides) for binding to a target site.
Recombination between Viral RNA and Transgene: Risks involved: The expression of viral sequences in transgenic plants is designed to confer protection against the donor virus and its related viruses and undoubtedly these unconventional crop protection strategies have provided extreme resistance or even immunity against virus infection. There are very bright prospects of commercialization of this techonology in the near future. Before the introduction of the transgenic crop plants in fields, the possibility of an interaction between the products of the transgene and a ''superinfecting" virus should properly be investigated, addressed and wisely considered. This could change the epidemiology of the infecting virus! viruses, resulting in the development of new diseases in areas or in crops where the disease has not occurred previously. Specifically there is the possibility that introduction of viral sequences into transgenic plants could, or would, affect RNA recombination events with and change the genomes of subsequent challenged viruses. Thus it could create new diseases or viruses with increased virulence or altered host range. The possibility of this potential danger needs to be resolved before large scale exploitation of transgenic plants. In the follwing paragraph, recombination of viral RNA with transgene products have been critically analyzed to resolve potential risks and bio-safty concerns involved with the commercialization of PDR.
It has been no experimental evidence of recombination between viral genome and the transgene. However, recently an evidence has been produced on RNA recombination and heteroencapsidation between related and unrelated viruses occurring in transgenic plants. The CP of plum pox potyvirus has confered aphid-transmissibility to a nontransmissible isolate of zucchini yellow mosaic virus (Lecoq et al., 1993). This particular case has raised the possibility of encapsidation of the genome of a "superinfecting" virus by the transgerically produced CP and thus has changed the vector specificity (Lecoq et al., 1993). More recently, a report on "Recombination between viral RNA and transgenic plant transcripts" (Greene and Allison, 1993), has posed threats to field release of transgenic plants. Transgenic plants expressing the 3' two-thirds of the cowpea chlorotic mottle virus (CCMV) CP gene were inoculated with a CCMV deletion mutant lacking the 3' one-third of the CP. Four of 125 inoculated transgenic plants became systemically infected and it was shown that systemic infection occurs only if recombination corrects and restores a functional CP gene. Analysis of viral RNA confirmed the recombination of transgenic mRNA and the challenging virus through aberrant homologous recombination.
There are a few reports available on homologous and heteralogous RNA recombinations in tobra- (Robinson et al., 1987; Angenent et al., 1989), bromo- (Bujarski and Kaesberg, 1986; Bujarski et al., 1994), alfamo- (Van der Kuyl et al., 1991), caulimo- (Gal et al., 1992) and nepoviruses (Scott et el., 1992). Recombination during virus replication could contribute to the rapid evolution of RNA viruses and could change host range or vector specificity, traits that have been attributed to the CP of several plant viruses (Dawson and Hilf, 1992) and the association between virus coded structural proteins and heterologous nucleic acid is significant if it creates new opportunities for virus/gene transfer (Cooper et al., 1994). RNA recombination events are clearly perceived as occurring frequently in nature and transgenically expressed viral mRNA is available to recombine with replicating RNA viruses, Therefore, RNA recombination should be considered as a vivid possibility when analysing the risks posed by virus-resistant transgenic crop plants.
The mechanism of PDR is due to a delay/slowdowning of the events of virus infection (entry into plant cell, bio-chemical replication, translocation, etc.) leading to resistance or immunity in some cases in agricultural crops. The mechanisms of different PDR strategies have extensively been reviewed (Lomonossoff, 1995; Baulcombe, 1996). PDR technologies could only be beneficial and commercially adaptable when hazards and risks involved are thoroughly investigated and carefully managed. A number of field trials have been conducted to assess the effectiveness of the PDR (Gonsalves and Slightom, 1993; Jongedijk et al., 1993; Kaniewski and Thomas, 1993). These trials have been mainly carried out with transgenic plants expressing viral coat protein with encouraging results. The main objective of this review is to highlight the prospects and importance of the PDR strategies and its application in the crop protection against viruses keeping in view its judicious application in order to minimize the potential risks involved in its commercialization.
Future Prospects: The significance of PDR for developing virus resistant plant is well established. Hopefully, in near future, more cases of PDR against different viruses, using different and broad spectrum chimeric gene cassettes from virus genomes, in various crops will be developed. Work on field release of plant with viral inserts is in progress and public concern about genetically modified crop is emerging. As scientists, it is our responsibility to accept the challenge and criticisms about the technology that we help to develop and to address the public concerns as honestly and openly as we possibly can. The modern approaches for the control of plant viruses using PDR technology which has extensively been reviewed in this paper would be a source of excitement and stimulation for the young scientists involved in Genetic Engineering and Biotechnology. Wide spread use of PDR techonology in the development of resistant varieties of crops shall revolutionize the agricultural productivity due to minimization crop losses against viruses. This review paper would serve as an easy and handy source of information for molecular biologists, agricultural scientists\researchers associated with the development of resistant varieties against prevalent viruses and shall also enhance the knowledge, understanding, awareness and use of this modern crop protection approach against numerous virus diseases of crop plants. With all things taken together, the future for controlling virus diseases looks bright.
REFERENCES
- Anderson, E.J., D.M. Stark, R.S. Nelson, P.A. Powell, N.E. Tumer and R.N. Beachy, 1989. Transgenic plants that express the coat protein genes of tobacco mosaic virus or alfalfa mosaic virus interfere with disease development of some nonrelated viruses. Phytopathology, 79: 1284-1290.
Direct Link - Anderson, J.M., P. Palukaitis and M. Zaitlin, 1992. A defective replicase gene induces resistance to cucumber mosaic virus in transgenic tobacco plants. Proc. Nat. Acad. Sci., 89: 8759-8763.
CrossRefDirect Link - Angenent, G.C., E. Posthumus, F.T. Brederode and J.F. Bol, 1989. Genome structure of tobacco rattle virus strain PLB: further evidence on the occurrence of RNA recombination among tobraviruses. Virology, 171: 271-274.
CrossRefDirect Link - Angenent, G.C., J.M. Van Den Ouweland and J.F. Bol, 1990. Susceptibility to virus infection of transgenic tobacco plants expressing structural and nonstructural genes of tobacco rattle virus. Virology, 175: 191-198.
CrossRefDirect Link - Arif, M., B. Reavy and H. Barker, 1999. Engineering high level of coat protein-mediated resistance to fungal transmission of potato mop-top furovirus in Nicotiana benthamiana. Pak. J. Biol. Sci., 2: 478-483.
CrossRefDirect Link - Audy, P., P. Palukaitis, S.A. Slack and M. Zaitlin, 1994. Replicase-mediated resistance to potato virus Y in transgenic tobacco plants. Mol. Plant Microbe Interactions, 7: 15-22.
Direct Link - Baulcombe, D.C., 1996. Mechanisms of pathogen-derived resistance to viruses in transgenic plants. Plant Cell, 8: 1833-1844.
CrossRefDirect Link - Baulcombe, D., 1994. Replicase-mediated resistance: A novel type of virus resistance in transgenic plants? Trends Microbiol., 2: 60-63.
CrossRefDirect Link - Bejarano, E.R. and C.P. Lichtenstein, 1994. Expression of TGMV antisense RNA in transgenic tobacco inhibits replication of BCTV but not ACMV geminiviruses. Plant Mol. Biol., 24: 241-248.
CrossRefDirect Link - Beachy, R.N., S. Loesch-Fries and N.E. Tumer, 1990. Coat protein-mediated resistance against virus infection. Annu. Rev. Phytopathol., 28: 451-472.
Direct Link - Beachy, R.N., 1997. Mechanisms and applications of pathogen-derived resistance in transgenic plants. Curr. Opin. Biotechnol., 8: 215-220.
PubMed - Braun, C.J. and C.L. Hemenway, 1992. Expression of amino-terminal portions or full-length viral replicase genes in transgenic plants confers resistance to potato virus X infection. Plant Cell, 4: 735-744.
CrossRefDirect Link - Bujarski, J.J. and P. Kaesberg, 1986. Genetic recombination between RNA components of a multipartite plant virus. Nature, 321: 528-531.
CrossRefDirect Link - Bujarski, J.J., P.D. Nagy and S. Flasinski, 1994. Molecular studies of genetic RNA-RNA recombination in brome mosaic virus. Adv. Virus Res., 43: 275-302.
CrossRefDirect Link - Burgyan, J., F. Grieco and M. Russo, 1989. A defective interfering RNA molecule in cymbidium ringspot virus infections. J. Gen. Virol., 70: 235-239.
CrossRefDirect Link - Carr, J.P., A. Gal-On, P. Palukaitis and M. Zaitlin, 1994. Replicase-mediated resistance to cucumber mosaic virus in transgenic plants involves suppression of both virus replication in the inoculated leaves and long-distance movement. Virology, 199: 439-447.
CrossRefDirect Link - Cooper, J.I., M.L. Edwards, O. Rosenwasser and N.W. Scott, 1994. Transgenic resistance genes from nepoviruses: efficacy and other properties. New Zealand J. Crop Hortic. Sci., 22: 129-137.
CrossRefDirect Link - Dawson, W.O. and M.E. Hilf, 1992. Host-range determinants of plant viruses. Annu. Rev. Plant Biol., 43: 527-555.
Direct Link - Day, A.G., E.R. Bejarano, K.W. Buck, M. Burrell and C.P. Lichtenstein, 1991. Expression of an antisense viral gene in transgenic tobacco confers resistance to the DNA virus tomato golden mosaic virus. Proc. Nat. Acad. Sci., 88: 6721-6725.
CrossRefDirect Link - Dougherty, W.G., J.A. Lindbo, H.A. Smith, T.D. Parks, S. Swaney and W.M. Proebsting, 1993. RNA-mediated virus resistance in transgenic plants: Exploitation of a cellular pathway possibly involved in RNA degradation. Mol. Plant-Microbe Interactions, 7: 544-552.
Direct Link - Frischmuth, T. and J. Stanley, 1991. African cassava mosaic virus DI DNA interferes with the replication of both genomic components. Virology, 183: 539-544.
CrossRefDirect Link - Gal, S., B. Pisan, T. Hohn, N. Grimsley and B. Hohn, 1992. Agroinfection of transgenic plants leads to viable cauliflower mosaic virus by intermolecular recombination. Virology, 187: 525-533.
CrossRefDirect Link - Gerlach, W.L., D. Llewellyn and J. Haseloff, 1987. Construction of a plant disease resistance gene from the satellite RNA of tobacco ringspot virus. Nature, 328: 802-805.
CrossRefDirect Link - Gielen, J.J.L., P. de Haan, A.J. Kool, D. Peters, M.Q.J.M. van Grinsven and R.W. Goidbach, 1991. Engineered resistance to tomato spotted wilt virus, a negative-strand RNA virus. Nat. Biotechnol., 9: 1363-1367.
CrossRef - Gielen, J., T. Ultzen, S. Bontems, W. Loots and A. Schepen et al., 1996. Coat protein-mediated protection to cucumber mosaic virus infections in cultivated tomato. Euphytica, 88: 139-149.
CrossRefDirect Link - Golemboski, D.B., G.P. Lomonossoff and M. Zaitlin, 1990. Plants transformed with a tobacco mosaic virus nonstructural gene sequence are resistant to the virus. Proc. Nat. Acad. Sci., 87: 6311-6315.
CrossRefDirect Link - Harrison, B.D., M.A. Mayo and D.C. Baulcombe, 1987. Virus resistance in transgenic plants that express cucumber mosaic virus satellite RNA. Nature, 328: 799-802.
CrossRefDirect Link - Haseloff, J. and W.L. Gerlach, 1988. Simple RNA enzymes with new and highly specific endoribonuclease activities. Nature, 334: 585-591.
Direct Link - Hillman, B.I., J.C. Carrington and T.J. Morris, 1987. A defective interfering RNA that contains a mosaic of a plant virus genome. Cell, 51: 427-433.
CrossRefDirect Link - Hobbs, S.L.A., T.D. Warkentin and C.M.O. DeLong, 1993. Transgene copy number can be positively or negatively associated with transgene expression. Plant Mol. Biol., 21: 17-26.
CrossRefDirect Link - Hoekema, A., M.J. Huisman, L. Molendijk, P.J. van den Elzen and B.J. Cornelissen, 1989. The genetic engineering of two commercial potato cultivars for resistance to potato virus X. Nat. Biotechnol., 7: 273-278.
CrossRefDirect Link - Jones, R.W., A.O. Jackson and T.J. Morris, 1990. Defective-interfering RNAs and elevated temperatures inhibit replication of tomato bushy stunt virus in inoculated protoplasts. Virology, 176: 539-545.
CrossRefDirect Link - Jongedijk, E., M.J. Huisman and B.J. Cornelissen, 1993. Argonic performance and field resistance of genetically modified, virus-resistant potato plants. Seminars Virol., 4: 407-416.
CrossRefDirect Link - Kallerhoff, J., P. Perez, S. Bouzoubaa, S.B. Tahar and J. Perret, 1990. Beet necrotic yellow vein virus coat protein-mediated protection in sugarbeet (Beta vulgaris L.) protoplasts. Plant Cell Rep., 9: 224-228.
CrossRefDirect Link - Kaniewski, W.K. and P.E. Thomas, 1993. Field testing of virus resistant transgenic plants. Seminars Virol., 4: 389-396.
CrossRefDirect Link - Kawchuk, L.M., R.R. Martin and J. McPherson, 1990. Resistance in transgenic potato expressing the potato leafroll virus coat protein gene. Mol. Plant-Microbe Interact, 3: 301-307.
Direct Link - Kunik, T., R. Salomon, D. Zamir, N. Navot and M. Zeidan et al., 1994. Transgenic tomato plants expressing the tomato yellow leaf curl virus capsid protein are resistant to the virus. Nat. Biotechnol., 12: 500-504.
CrossRefDirect Link - Kurath, G. and J.A. Dodds, 1994. Satellite tobacco mosaic virus sequence variants with only five nucleotide differences can interfere with each other in a cross protection-like phenomenon in plants. Virology, 202: 1065-1069.
CrossRefDirect Link - Lawson, C., W. Kaniewski, L. Haley, R. Rozman, C. Newell, P. Sanders and N.E. Tumer, 1990. Engineering resistance to mixed virus infection in a commercial potato cultivar: Resistance to potato virus X and potato virus Y in transgenic Russet Burbank. Bio/Technology, 8: 127-134.
CrossRefDirect Link - Lecoq, H., M. Ravelonandro, C. Wipf-Scheibel, M. Monsion, B. Raccah and J. Dunez, 1993. Aphid transmission of a non-aphid-transmissible strain of zucchini yellow mosaic potyvirus from transgenic plants expressing the capsid protein of plum pox potyvirus. Mol. Plant-Microbe Interactions, 6: 403-406.
Direct Link - L'Huillier, P.J., S.R. Davis and A.R. Bellamy, 1992. Cytoplasmic delivery of ribozymes leads to efficient reduction in alpha-lactalbumin mRNA levels in C127I mouse cells. EMBO J., 11: 4411-4418.
Direct Link - Li, X.H., L.A. Heaton, T.J. Morris and A.E. Simon, 1989. Turnip crinkle virus defective interfering RNAs intensify viral symptoms and are generated de novo. Proc. Nat. Acad. Sci., 86: 9173-9177.
Direct Link - Lindbo, J.A. and W.G. Dougherty, 1992. Pathogen-derived resistance to a potyvirus: Immune and resistant phenotypes in transgenic tobacco expressing altered forms of a potyvirus coat protein nucleotide sequence. Mol. Plant Microbe Interact, 5: 144-153.
Direct Link - Lindbo, J.A. and W.G. Dougherty, 1992. Untranslatable transcripts of tobacco etch virus coat protein gene sequence can interfere with tobacco etch virus replication in transgenic plants and protoplasts. Virology, 189: 725-733.
CrossRefDirect Link - Lius, S., R.M. Manshardt, M.M. Fitch, J.L. Slightom, J.C. Sanford and D. Gonsalves, 1997. Pathogen-derived resistance provides papaya with effective protection against papaya ringspot virus. Mol. Breed., 3: 161-168.
CrossRefDirect Link - Lodge, J.K., W.K. Kaniewski and N.E. Tumer, 1993. Broad-spectrum virus resistance in transgenic plants expressing pokeweed antiviral protein. Proc. Nat. Acad. Sci., 90: 7089-7093.
Direct Link - Lomonossoff, G.P., 1995. Pathogen-derived resistance to plant viruses. Annu. Rev. Phytopathol., 33: 323-343.
CrossRefDirect Link - Longstaff, M., G. Brigneti, F. Boccard, S. Chapman and D. Baulcombe, 1993. Extreme resistance to potato virus X infection in plants expressing a modified component of the putative viral replicase. EMBO J., 12: 379-386.
Direct Link - Marsh, L.E., G.P. Pogue, J.P. Connell and T.C. Hall, 1991. Artificial defective interfering RNAs derived from brome mosaic virus. J. Gen. Virol., 72: 1787-1792.
CrossRefDirect Link - Marsh, L.E., G.P. Pogue, U. Szybiak, J.P. Connell and T.C. Hall, 1991. Non-replicating deletion mutants of brome mosaic virus RNA-2 interfere with viral replication. J. Gen. Virol., 72: 2367-2374.
CrossRefDirect Link - MacKenzie, D.J. and J.H. Tremaine, 1990. Transgenic Nicotiana debneyii expressing viral coat protein are resistant to potato virus S infection. J. Gen. Virol., 71: 2167-2170.
CrossRefDirect Link - MacKenzie, D.J. and P.J. Ellis, 1992. Resistance to tomato spotted wilt virus infection in transgenic tobacco expressing the viral nucleocapsid gene. Mol. Plant-Microbe Interact, 5: 34-40.
Direct Link - MacKenzie, D.J., J.H. Tremaine and J. McPherson, 1991. Genetically engineered resistance to potato virus S in potato cultivar Russet Burbank. Mol. Plant-Microbe Interact., 4: 95-102.
CrossRefDirect Link - Mannerlof, M., B.L. Lennerfors and P. Tenning, 1996. Reduced titre of BNYVV in transgenic sugar beets expressing the BNYVV coat protein. Euphytica, 90: 293-299.
CrossRefDirect Link - Mazzolini, L., M. Axelos, N. Lescure and P. Yot, 1992. Assaying synthetic ribozymes in plants: high-level expression of a functional hammerhead structure fails to inhibit target gene activity in transiently transformed protoplasts. Plant Mol. Biol., 20: 715-731.
CrossRefDirect Link - McGarvey, P.B., M.S. Montasser and J.M. Kaper, 1994. Transgenic tomato plants expressing satellite RNA are tolerant to some strains of cucumber mosaic virus. J. Am. Soc. Hortic. Sci., 119: 642-647.
Direct Link - Mueller, E., J. Gilbert, G. Davenport, G. Brigneti and D.C. Baulcombe, 1995. Homology‐dependent resistance: Transgenic virus resistance in plants related to homology‐dependent gene silencing. Plant J., 7: 1001-1013.
CrossRefDirect Link - Nelson, R.S., P.P. Abel and R.N. Beachy, 1987. Lesions and virus accumulation in inoculated transgenic tobacco plants expressing the coat protein gene of tobacco mosaic virus. Virology, 158: 126-132.
CrossRefDirect Link - Nelson, R.S., S.M. McCormick, X. Delannay, P. Dube and J. Layton et al., 1988. Virus tolerance, plant performance of transgenic tomato plants expressing coat protein from tobacco mosaic virus. Nat. Biotechnol., 6: 403-409.
CrossRefDirect Link - Ploeg, A.T., A. Mathis, J.F. Bol, D.J. Brown and D.J. Robinson, 1993. Susceptibility of transgenic tobacco plants expressing tobacco rattle virus coat protein to nematode-transmitted and mechanically inoculated tobacco rattle virus. J. Gen. Virol., 74: 2709-2715.
CrossRefDirect Link - Ponz, F., A. Rowhani, S.M. Mircetich and G. Bruening, 1987. . Cherry leafroll virus infections are affected by a satellite RNA that the virus does not support. Virology, 160: 183-190.
CrossRefDirect Link - Prins, M. and R. Goldbach, 1996. RNA-mediated virus resistance in transgenic plants. Arch. Virol., 141: 2259-2276.
CrossRefDirect Link - Reavy, B., M. Arif, S. Kashiwazaki, K.D. Webster and H. Barker, 1995. Immunity to potato mop-top virus in Nicotiana benthamiana plants expressing the coat protein gene ls effective against fungal lnoculation of the virus. Mol. Plant-Microbe Interactions, 8: 286-291.
Direct Link - Regner, F., A. da Camara Machado, M.L. da Camara Machado, H. Steinkellner and D. Mattanovich et al., 1992. Coat protein mediated resistance to plum pox virus in Nicotiana clevelandii and N. benthamiana. Plant Cell Rep., 11: 30-33.
CrossRefDirect Link - De Oliveira Resende, R., P. de Haan, A.C. de Avila, E.W. Kitajima, R. Kormelink, R. Goldbach and D. Peters, 1991. Generation of envelope and defective interfering RNA mutants of tomato spotted wilt virus by mechanical passage. J. Gen. Virol., 72: 2375-2383.
CrossRefDirect Link - De Oliveira Resende, R., P. de Haan, E. van de Vossen, R. Goldbach and D. Peters, 1992. Defective interfering L RNA segments of tomato spotted wilt virus retain both virus genome termini and have extensive internal deletions. J. Gen. Virol., 73: 2509-2516.
CrossRefDirect Link - Robinson, D.J., W.D.O. Hamilton, B.D. Harrison and D.C. Baulcombe, 1987. Two anomalous tobravirus isolates: Evidence for RNA recombination in nature. J. Gen. Virol., 68: 2551-2561.
CrossRefDirect Link - Roossinck, M.J., D. Sleat and P. Palukaitis, 1992. Satellite RNAs of plant viruses: Structures and biological effects. Microbiol. Rev., 56: 265-279.
Direct Link - Rubino, L., R. Lupo and M. Russo, 1993. Resistance to cymbidium ringspot tombusvirus infection in transgenic Nicotiana benthamiana plants expressing a full-length viral replicase gene. Mol. Plant Microbe Interactions, 6: 729-734.
Direct Link - Silva-Rosales, L., J.A. Lindbo and W.G. Dougherty, 1994. Analysis of transgenic tobacco plants expressing a truncated form of a potyvirus coat protein nucleotide sequence. Plant Mol. Biol., 24: 929-939.
CrossRefDirect Link - Slightom, J.L., P.P. Chee and D. Gonsalves, 1990. Field Testing of Cucumber Plants which Express the CMV Coat Protein Gene: Field Plot Design to Test Natural Infection Pressures. In: Progress in Plant Cell and Molecular Biology, Nijkamp, H.J.J., L.P.H.W. van der Plass and J. van Aartriik (Eds.). Kluwer Academic Publisher, Dordrecht, pp: 201-206.
- Smith, G.R., P.A. Joyce, J.A. Handley, P. Sithisarn and M.M. Maugeri et al., 1996. Genetically Engineering to Sugarcane Mosaic and Fiji Disease Viruses in Sugarcane. In: Sugarcane: Research Towards Efficient and Sustainable Production, Wilson, J.R., D.M. Hogarth, J.A. Campbell and A.L. Garside (Eds.). CSIRO Division of Tropical Crops and Pastures, Brisbane, Australia, pp: 138-140.
- Stanley, J., T. Frischmuth and S. Ellwood, 1990. Defective viral DNA ameliorates symptoms of geminivirus infection in transgenic plants. Proc. Nat. Acad. Sci., 87: 6291-6295.
CrossRefDirect Link - Stark, D.M. and R.N. Beachy, 1989. Protection against potyvirus infection in transgenic plants: Evidence for broad spectrum resistance. Nat. Biotechnol., 7: 1257-1262.
CrossRefDirect Link - Stenger, D.C., 1994. Strain-specific mobilization and amplification of a transgenic defective-interfering DNA of the geminivirus beet curly top virus. Virology, 203: 397-402.
CrossRefDirect Link - Steinecke, P., T. Herget and P.H. Schreier, 1992. Expression of a chimeric ribozyme gene results in endonucleolytic cleavage of target mRNA and a concomitant reduction of gene expression in vivo. EMBO J., 11: 1525-1530.
Direct Link - Tavladoraki, P., E. Benvenuto, S. Trinca and D. De Martinis, 1993. Transgenic plants expressing a functional single-chain Fv antibody are specifically. Nature, 366: 469-472.
Direct Link - Taylor, S., A. Massiah, G. Lomonossoff, L.M. Roberts, J.M. Lord and M. Hartley, 1994. Correlation between the activities of five ribosome‐inactivating proteins in depurination of tobacco ribosomes and inhibition of tobacco mosaic virus infection. Plant J., 5: 827-835.
CrossRefDirect Link - Truve, E., A. Aaspollu, J. Honkanen, R. Puska and M. Mehto et al., 1993. Transgenic potato plants expressing mammalian 2′–5′ oligoadenylate synthetase are protected from potato virus X infection under field conditions. Nat. Biotechnol., 11: 1048-1052.
CrossRefDirect Link - Tumer, N.E., K.M. O'Connell, R.S. Nelson, P.R. Sanders, R.N. Beachy, R.T. Fraley and D.M. Shah, 1987. Expression of alfalfa mosaic virus coat protein gene confers cross-protection in transgenic tobacco and tomato plants. EMBO J., 6: 1181-1188.
Direct Link - Van der Kuyl, A.C., L. Neeleman and J.F. Bol, 1991. Complementation and recombination between alfalfa mosaic virus RNA3 mutants in tobacco plants. Virology, 183: 731-738.
CrossRefDirect Link - Van der Wilk, F., D.P.L. Willink, M.J. Huisman, H. Huttinga and R. Goldbach, 1991. Expression of the potato leafroll luteovirus coat protein gene in transgenic potato plants inhibits viral infection. Plant Mol. Biol., 17: 431-439.
CrossRefDirect Link - Van Dun, C.M. and J.F. Bol, 1988. Transgenic tobacco plants accumulating tobacco rattle virus coat protein resist infection with tobacco rattle virus and pea early browning virus. Virology, 167: 649-652.
CrossRefDirect Link - Van Dun, C.M., J.F. Bol and L. Van Vloten-Doting, 1987. Expression of alfalfa mosaic virus and tobacco rattle virus coat protein genes in transgenic tobacco plants. Virology, 159: 299-305.
CrossRefDirect Link - Van Dun, C.M., L. Van Vloten-Doting and J.F. Bol, 1988. Expression of alfalfa mosaic virus cDNA1 and 2 in transgenic tobacco plants. Virology, 163: 572-578.
CrossRefDirect Link - Van Dun, C.M., B. Overduin, L. van Vloten-Doting and J.F. Bol, 1988. Transgenic tobacco expressing tobacco streak virus or mutated alfalfa mosaic virus coat protein does not cross-protect against alfalfa mosaic virus infection. Virology, 164: 383-389.
CrossRefDirect Link - White, K.A. and T.J. Morris, 1994. Enhanced competitiveness of tomato bushy stunt virus defective interfering RNAs by segment duplication or nucleotide insertion. J. Virol., 68: 6092-6096.
Direct Link - Wilson, T.M., 1993. Strategies to protect crop plants against viruses: Pathogen-derived resistance blossoms. Proc. Nat. Acad. Sci., 90: 3134-3141.
CrossRefDirect Link - Wilson, T.M.A. and J.W. Davies, 1994. New roads to crop protection against viruses. Outlook Agric., 23: 33-39.
Direct Link - Yie, Y., F. Zhao, S.Z. Zhao, Y.Z. Liu, Y.L. Liu and P. Tien, 1992. High resistance to cucumber mosaic virus conferred by satellite RNA and coat protein in transgenic commercial tobacco cultivar G-140. Mol. Plant-Microbe Interact, 5: 460-465.
Direct Link - Yie, Y. and P. Tien, 1993. Plant virus satellite RNAs and their role in engineering resistance to virus diseases. Seminars Virol., 4: 363-368.
CrossRefDirect Link - Zaitlin, M., J.M. Anderson, K.L. Perry, L. Zhang and P. Palukaitis, 1994. Specificity of replicase-mediated resistance to cucumber mosaic virus. Virology, 201: 200-205.
CrossRefDirect Link