
Yazid A.M
Department of Food Technology, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
A.B. Salina
Department of Food Technology, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
M. Shuhaimi
Department of Food Technology, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
H. Osman
Department of Food Science and Nutrition, Universiti Kebangsaan Malaysia, 47600, Bangi, Selangor, Malaysia
J. Normah
Department of Food Technology, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 3 | Page No.: 660-663







ABSTRACT
Four probiotic microorganisms were tested for antimicrobial activity against selected food-borne pathogens namely Escherichia coli, Salmonella enteritidis, Listetia monocytogenes and Bacillus cereus. Bifidobacterium breve C11 and Streptococcus faecalis showed maximum antibacterial activity against all target microorganisms, while Bifidobacterium infantis C15 had the least antibacterial activity. Furthermore, the results of viable counts and pH profile revealed that the combination of probiotic microorganisms exerts better inhibitory effect against enteropathogenic E. coli than a singIe probiotic dose.
PDF Abstract XML References Citation
How to cite this article
Yazid A.M, A.B. Salina, M. Shuhaimi, H. Osman and J. Normah, 1999. Inhibitory Effects of Probiotic Bacteria Against Selected Food-Borne Pathogens. Pakistan Journal of Biological Sciences, 2: 660-663.
DOI: 10.3923/pjbs.1999.660.663
URL: https://scialert.net/abstract/?doi=pjbs.1999.660.663
DOI: 10.3923/pjbs.1999.660.663
URL: https://scialert.net/abstract/?doi=pjbs.1999.660.663
INTRODUCTION
The concept of probiotic was first introduced in early 1900’s by Tissier (Fuller, 1989). However, the term was only used in 1965 by Lilly and Stillwell and has subsequently evolved. Numerous definitions have been proposed. Initially, Lilly and Stillwell (1965) defined probiotic as a substance secreted by a microorganism which can stimulate the growth of another microorganisms (Fuller, 1989). Today, a probiotic refers to any preparation "of live microorganisms which when applied to man or animal can beneficially affect the host by maintaining the intestinal microbial balance" (Havenaar and Huis, 1992). Lactic acid bacteria (LAB) namely lactobacilli, lactococci, streptococci, enterococci and Bifidobacterium spp. are the common microorganisms which have been used as probiotic preparations (Fuller, 1989).
The effect of probiotic organism on the micro ecology of the gut is to some extent dependent upon its ability to survive and preferably inhibit the proliferation of pathogens. In addition, various compounds produced during growth of the probiotic have been shown to inhibit the growth of pathogen such as Vibrio cholera and Bacillus cereus (Klaenhammer, 1988). These compounds include organic acids such as lactic and acetic acid, and antibiotic-like compounds such as reuterin and bacteriocin (Tagg et al., 1976). The organic acids lower the pH and thereby indirectly affect growth of the pathogen. Numerous bacteriocin have been reported to be produced by probiotic microorganisms namely Acidophilin, Bifidin and Nisin (Klaenhammer, 1988). They can either have a very broad range of activity or specifically inhibit the growth of very limited range of closely related microbes.
The objectives of the present investigation were to study the antagonistic action of probiotic microorganisms against selected food-borne pathogens and to elucidate the mechanism of such action.
MATERIALS AND METHODS
Probiotic strains: The strains Bifidobacterium breve C11 and Bifidobacterium infantis C15 used in this study were obtained from the Probiotic Laboratory Culture Collection, Universiti Putra Malaysia (UPM). They were isolated from fresh infant stool. They were cultured in Trypticase Phytone Yeast-extract medium (TPY) as recommended by Scardovi (1986). Bacillus mesentricus TO-A and Streptococcus faecalis T-110 obtained from Toa Pharmaceutical, Japan and maintained propagated in nutrient medium (Lino et al., 1993) and Glucose-Yeast-Peptone (GYP) medium (Seo et al., 1989) respectively.
Target cultures of pathogenic organisms: The target cultures of Escherichia coli, Salmonella enteritidis, Listeriz monocytogenes and Bacillus cereus were obtained from the Tissue Culture Laboratory, UPM. They were maintained and propagated in Tryptone Soy (TS) medium.
Assay for inhibitory activity: Using a method modified from Vignolo et al. (1993), the 18 h culture of probiotic microorganisms were spotted on the respective agar medium using a sterile toothpick. B. breve C11, B. infantis C15 and S. faecalis T-110 were incubated anaerobically a 37°C for 48 h. B. mesentritus TO-A was incubated aerobically at 37°C for 48 h. The growing cultures were overlaid with soft (TS) agar seeded with 0.1 percent (v/v), freshly prepared target organisms. The overlaid cultures were incubated aerobically at 37°C for 24 h. The diameter of the inhibition zone was measured.
Inhibition of the growth of Escherichia coli in single and mixed cultures of probiotic microorganisms: Method used by Seo et al. (1989) was modified to study the inhibition of Escherichia coli by the probiotic isolates using the following combinations namely, combination (1) S. faecalis T-110 against E. coli, (2) B. breve Cl 1 with B. infantis C15 against E. coli, (3) B. breve C11 with B. infantis C15 and S. faecalis T-110 against E. coil and (4) B. breve C11 with B. infantis C15, S. faecalis T-1 10 and B. mesentricus TO-A against E. coli. All the cultures used were at a concentration of 108 CFU/ml. After inoculation, the viable count of E. coli was determined at times intervals of 0, 2, 4, 6. 8, 10, 12, 16 and 24 h by plating it on McConkey agar and incubated at 37°C for 24 h aerobically. The pH of the mixed cultures was also measured at different time intervals.
RESULTS AND DISCUSSION
B. breve C11 and S. faecalls T-110 showed maximum antibacterial activity followed by B. mesentricus TO-A and B. infantis C15 (Table 1). B. mesentricus TO-A showed maximum zone of inhibition against B. cereus and L. monocytogenes where it inhibited B. cereus greater than L. monocytogenes. The growth of E. coli and S. enteritidis were unaffected by the present of B. infantis C15 and B. mesentricus TO-A. B. infantis C15 showed the least antibacterial activity in most cases.
A combination of probiotic microorganisms was used for the treatment of diarrhea and constipation with enteropathogenic strains including pathogenic E. coli (Seo et al., 1989). They reported that after 12-24 h S. faecalis and C. butyricum, either separately or combined, inhibited E. coli growth at a ratio of 1/100-1/1000. Yet a combination of S. faecalis and C. butyricum resulted a stronger effect and growth of E. coli was not observed on agar plates 36 h after inoculation with these strains. A symbiosis between these bacteria strongly inhibited the proliferation of S. typhirnurium, V. parahaemolyticus, C. difficile and C. botulinum.
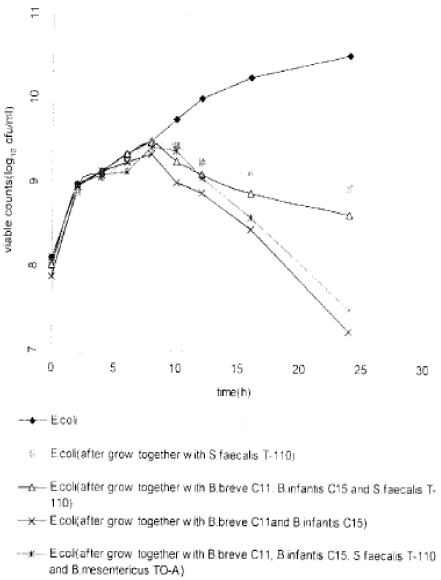 | |
| Fig. 1: | Changes in counts of E. coli grown in pure and mixed cultures of probiotics |
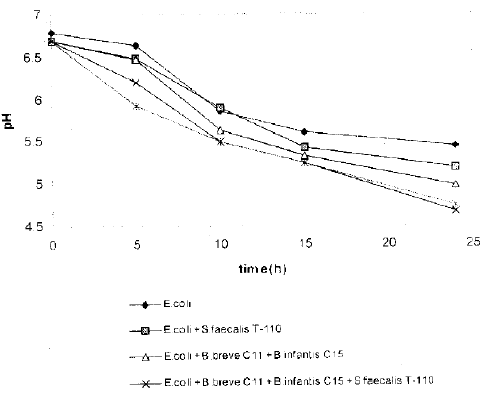 | |
| Fig. 2: | Changes in pH of the culture medium |
| Table 1: | Inhibitory activity of probiotic microorganisms against selected food-borne pathogens |
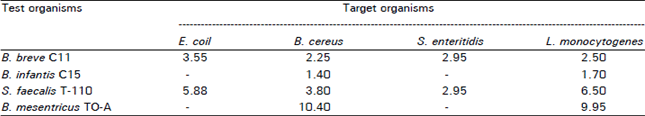 | |
| *Diameter in mm (without the diameter of the spot) of inhibition zone, -: Corresponds to the absence of clear inhibition zone | |
The results of the present study are shown in Fig. 1. The proliferation of combined probiotic microorganisms affected the growth of E.coli more significantly than did proliferation of a single probiotic. We found that after incubating E. coil with S. faecalis T-110 alone, the viable count of E. coil decreased to 101 CFU/ml after 24 h. However, when E. coil was inoculated with mixed cultures of S. faecalis T-1 10, B. breve 011 and B. infantis 015 simultaneously, the viable count of E. coli decreased to 102 CFU/ml. Likewise when E. coli was inoculated with mixed cultures of B. breve C11 and B. infantis C15, the growth of E. coli was inhibited and almost eliminated after 24 h. The decrease in viable count of E. coli was about 104 CFU/ml. The same pattern was observed when E. coli was grown together with B. breve C11, B. infantis C15, S. faecalis T-110 and B. mesentricus TO-A, where the decrease in viable counts of E. coli was about 103 CFU/ml after 24h. These findings show the establishment of the symbiotic relationship among probiotic microorganisms, which enhances the inhibition pathogenic E. coli.
The pH profile of the spent culture medium also suggested that the combination of probiotic microorganism significantly reduced the pH of the medium than a single probiotic did (Fig. 2). The medium pH of combination of B. breve C11, B. infantis 015 and S. faecalis T-110 against E. coli decreased by 2.12 percent (data not shown) companion with a pH of medium of S. faecalis ’T-110 alone which reduced by 0.06 percent (data not shown). This is due the fact that mixed Bifidobacteria and S. faecalis capable of producing much more acetic and lactic ac during their glucose metabolism (Scardovi, 1986). On contrary, the addition of B. mesentricus TO-A in the mi cultures of B. breve C11, B. infantis 015 and S. faecalis 110 slightly increased the medium pH by 0.04 percent (data not shown). B. mesentricus is known to produced amylase arid protease which are neutral and alka respectively, a fact that might explain the increase in the pH of the medium (Leifert et al., 1995).
From the results discussed above, it clearly shows that the combination of the selected probiotic microorganisms exerts better inhibitory effect against food-borne pathogens than a single probiotic does. This antagonistic effect has been attributed much to the symbiotic association among the probiotic microorganisms themselves, which produce higher amount of bactericidal substances. Thus, they provide better prevention of disease to the host.
REFERENCES
- Fuller, R., 1989. Probiotics in man and animals. J. Applied Bacteriol., 66: 365-378.
CrossRefPubMedDirect Link - Seo, G.I., K. Shimizu, M. Sasatsu and M. Kono, 1989. Inhibition of growth of some enteropathogenic strains in mixed cultures of Streptococcus faecalis and Clostridium butyricum. Microbios. Lett., 40: 151-160.
Direct Link - Vignolo, G.M., F. Suriani, A.P.R Holgado and G. Oliver, 1993. Antibacterial activity of Lactobacillus strains isolated from dry fermented sausages. J. Applied Bacteriol., 75: 344-349.
CrossRefDirect Link - Lino, H., K. Fukaya, Y. Hirasawa, K. Shimizu and G. Seo, 1993. Stimulation of bacterial growth of some strains of Bifidobacterium by a crude preparation of metabolites from Bacillus mesentericus TO-A. Biomed. Lett., 48: 73-78.
Direct Link - Klaenhammer, T.R., 1988. Bacteriocins of lactic acid bacteria. Biochimie, 70: 337-349.
CrossRefDirect Link - Leifert, G., H. Li, S. Chidburee, S. Hampson and S. Workman et al., 1995. Antibiotic production and biocontrol activity by Bacillus subtilis CL27 and Bacillus pumilus CL45. J. Applied Bacteriol., 78: 97-108.
CrossRefPubMedDirect Link - Lilly, D.M. and R.H. Stillwell, 1965. Probiotics: Growth-promoting factors produced by microorganisms. Science, 147: 747-748.
CrossRefDirect Link - Tagg, J.R., A.S. Dajani and L.W. Wannamaker, 1976. Bacteriocins of gram positive bacteria. Bacteriol. Rev., 40: 722-756.
Direct Link