
Pear Mohammad
The United Graduate School of Agricultural Sciences and *Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
Masaya Shiraishi
The United Graduate School of Agricultural Sciences and *Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 2 | Page No.: 565-570







ABSTRACT
Anatomical responses of roots in trifoliate orange budded with satsuma mandarin (Citrus unshiu Marc. cv. Okitsu Wase) to aerated and stagnant culture solutions were investigated. Roots, under aerated condition, had sharply pointed tips and showed intact anatomical features having a few starch grains on the rootcap and cortical cells. The cortical cells were smaller close to the root tip and gradually became elongated towards the base. All these characteristics maintained the same level even up to 10 days of observations. On the contrary, under stagnant condition, secretion of mucilaginous substances and root tip injuries were found on the 3rd day of induction which became more conspicuous on the 5th. These injuries resulted the severe sloughing of cells and starch grain degradation in the rootcap. Eight day induced roots displayed more distinct root tip injuries and discharging of mucilaginous substances. On the 10th day, air spacing between the cortical cells were conspicuous. Epidermal injuries were also evidenced sporadically on the 5th day and became more severe subsequently. Degradation of starch grains gradually completed in the cortical cells along the exposure time. The enlargement of periblemic cells were noticed which gradually resulted larger cells close to the root apex on longer exposures.
PDF Abstract XML References Citation
How to cite this article
Pear Mohammad and Masaya Shiraishi, 1999. Anatomical Responses of Roots in Trifoliate Orange Budded with Satsuma Mandarin to Aerated and Stagnant Culture Solutions. Pakistan Journal of Biological Sciences, 2: 565-570.
DOI: 10.3923/pjbs.1999.565.570
URL: https://scialert.net/abstract/?doi=pjbs.1999.565.570
DOI: 10.3923/pjbs.1999.565.570
URL: https://scialert.net/abstract/?doi=pjbs.1999.565.570
INTRODUCTION
Plants in anoxia have been the subject of experimental studies since the early 1900's as the higher plants have an absolute requirement for oxygen to sustain metabolism and growth. The classic works of Steward (1935) and Hoagland and Broyer (1936) established that oxygen is indispensable for vital activity in plant roots and that the energy for nutrient and water absorption comes from the oxidation of carbohydrates in roots. Other complementary studies also showed that oxygen concentration in the root environment relates to plant activities (Willey, 1970; Buwalda et al., 1988; Nobel and Palta, 1989; Palta and Nobel, 1989) and lack of oxygen or an excess of CO2 both of which can depress root respiration (Jackson and Drew, 1984; Smit and Stachowiak, 1988). Root respiration is required for plant growth, mineral uptake and perhaps, indirectly for water absorption (Lemon, 1962). However, these phenomena under anaerobiosis are extensively described in different crops (Nobel and Palta, 1989; Palta and Nobel, 1989; Zieslin and Snir, 1989; Fox and Kennedy, 1991; Kennedy et al., 1992; Zhang et al., 1994).
Up to date it is known that although citrus is not much sensitive to poor aeration, it is fairly intolerable like other plants to lack of sufficient oxygen for root respiration and the secondary soil conditions resulting therefrom (Chapman, 1968). Considering the importance of aeration to root health and nutrient and water absorption, this subject has not received deserved attention in citrus (Chapman, 1968). How this plant species is reacting against anaerobiosis may be an useful topic to enrich our present knowledge on stress physiology. Recently we have observed that root formation, root growth and root morphology in satsuma mandarin were adversely affected by stagnant condition (Mohammad and Shiraishi, 1999). For increased understanding, sufficient knowledge on the matter at the cellular level are warranted. In this connection it is suggested that growing cells do not necessarily respond to environmental stresses in the same way as the mature tissue (Nonami and Boyer, 1989; Pritchard et al., 1991), indicating that the presence of mature tissue may mask the response of growing cells. Thus determination of different parameters in mature tissue does not accurately reflect controls of growth since the investigations on growing tissue or cells are logical. In addition, apparently the cell length patterns help the plant adapt to environmental stress (Silk, 1992) which can be used as the indicator of plant growth state.
This investigation was designed to elucidate the anatomical responses of young roots of trifoliate orange budded with satsuma mandarin to stagnant culture solution compared to aerated condition. The root tip shapes, their changes, starch grain degradation trends in the cortical and rootcap cells and cell length patterns of cortical files close to the root apex were emphasized.
MATERIALS AND METHODS
The experiment was conducted in the Citriculture Laboratory, Faculty of Agriculture, Ehime University, Japan during June to August of three consecutive years (1996-1998) using three-year-old satsuma mandarin trees grafted on trifoliate orange rootstocks. Twelve trees for uniformity of vigor were placed into six styrofoam boxes retaining nutrient solution each year. The nutrient solution contained N, Ca, K, S, Mg, P, Na, CI, Mn, Fe, B, Mo, Zn and Cu at approximately the following ppm, respectively: 198, 160, 80, 38, 23, 15, 11, 1, 0.5, 0.4, 0.25, 0.1, 0.05 and 0.02 as was used by Smith (1971). The pH of the culture solutions were adjusted to 5.0 daily by using 1M H2SO4 or 3M NaOH as were required. The nutrient solution of all boxes were continuously aerated and stirred by air pumps. To ensure an adequate supply of all essential elements, the nutrient solutions were completely renewed at every 14 days intervals. The average daily air temperatures were recorded as 22-29°C during the experiment.
On the formation of numerous new roots they were marked with wax-coated paper tags. Then three boxes with six trees were continuously aerated and stirred by air pumps whereas the another three boxes having other six trees were not done so to create stagnant condition. Roots of 3, 5, 8 and 10 days induction under aerated and stagnant conditions were fixed in 4 percent glutaraldehyde buffered to pH 7.4 in sodium cacodylate buffer for 48 hr in the refrigerator, dehydrated in a graded ethanol series and infiltrated and embedded in JB-4 resin. Median longitudinal sections of three micrometer thickness were cut with dry glass knives on a sorval MT-1 ultramicrotome and were stained with iodine-potassium-iodide. The sections were viewed under light microscope and the photographs were taken with the camera mounted on the microscope. About 30 sections of each root were viewed and at least five were photographed. Data on cell lengths were obtained from prints with a total magnification of X1000 as was done by Bertaud and Gandar (1985). The same files were used for the measurements. Cell length at each point was taken as an average of 25 to 35 cells. At least 5 roots from each treatment at every sampling date were taken into account. The 3-year-observations were summarized and reported.
RESULTS
Anatomical features with special emphasis on root tip: The roots under aerated condition had normal anatomy at every exposure (Fig. 1A-D). The roots also showed normal growing state with sharply pointed tips. Normal removal of older rootcap cells was distinct. Growing region was quite wide and clearly distinguished. The rootcap cells were furnished with well developed column of cells. A similar trend of anatomical features continued and even up to the end of sampling normal anatomy was found. The cortical cells were elongated, especially beyond the meristematic zone (Fig. 2A-H). The quiescent center positioned apart from the root terminal due to the presence of well developed rootcap cells (Fig. 1A-D). These roots received no injury in any parts.
In contrast, under stagnant condition, root tip injuries and other growth inhibiting symptoms were gradually evidenced (Fig. 1E-H). However, on the 3rd day, root tip became narrower and dome-shaped with terracing immediately back to the quiescent center (Fig. 1E). Secretion of mucilaginous substances from the root tips became distinct (Fig. 1E) and the meristematic cells near the root apex became round followed by the formation of air space (arrow heads) through the lysigenous breakdown of cells (Fig. 2I). The epidermal injuries (El) initiated in few places (Fig. 2J). The rootcap cells severely sloughed off. The 5-day-induced roots showed injuries in the root tips followed by the secretion of mucilaginous substances and the rootcap cells mostly sloughed off (Fig. 1F). The cell rounding and air spacing (arrow heads) became more proximal to the root apex (Fig. 2K). The meristematic region was shorter compared to aerated condition and the epidermal injuries became clear at least in few places (Fig. 1F, 2L). On the 8th day after the sloughing of the rootcap and some portions of apical meristem the root tip became irregular (Fig. 1G) and the epidermal injury (El) induction process continued (Fig. 2N). The cell rounding became totally proximal to the root tip. The secretion of mucilaginous substances and formation of air spaces (arrow heads) in the cortex remained constant (Fig. 1G, 2M). Ten-day-induced roots were similarly under root tip injury and showed irregular arrangement of apical meristem cells (Fig. 1H). The meristematic zone became malformed. The cells in the cortical region were under modification. The formation of intercellular air spaces (arrow heads) on the cortex formed bigger air spaces throughout (Fig. 2O, P). Epidermal injuries were also found in many places.
Starch grain accumulation and degradation pattern: Under aerated condition, starch grains accumulated on the rootcap cells, especially on the outer layers of rootcap region (Fig. 1A-D). The cortical cells were devoid of grains near the root tip (Fig. 2A, C, E, G). More or less 3-5 mm back from the root apex only a few starch grains (SG) were accumulated in the cortical cells near the epidermis (Fig. 2B, D, F, H). The trend of this accumulation presumed to be similar in all exposures. However, a common trend was noticedlhat starch grain deposition was higher towards the root base. In contrast, starch grains were completely absent from the rootcap and the quiescent center adjacent cortical cells of all exposures (Fig. 1E-H, 2I, K, M, O). The 3-day-induced foots contained much starch grains on the cortical cells about 3-5 mm back from the root apex (Fig. 2J). The grains near the epidermal injury were under degradation, The 5-day-induced roots contained only a few grains on the cortical cells apart from the root tip and the degradation of grains were distinct near the injury (Fig. 2L). Starch grains were completely degraded from the cortical cells of 8 and 10-day-induced roots (Fig. 2N, P).
Cell length in the cortical files: The root apex adjacent cells were smaller and gradually became larger at aerated condition (Fig. 3). The trend of enlargement of cells towards the root base was regular in all exposures. Usually up to 1 mm from the root apex, cell length remained within 201 μm and then became larger under this condition. In contrast, under stagnant condition, although up to 3 days the apical meristem adjacent cells were smaller, became larger towards the root base. With the exposure time enlargement of meristematic cells occurred close to the root apex (Fig. 4). Accordingly, the meristematic zone extended up to only the cells around the quiescent center in the roots under 3, 5 and 8 days induction.
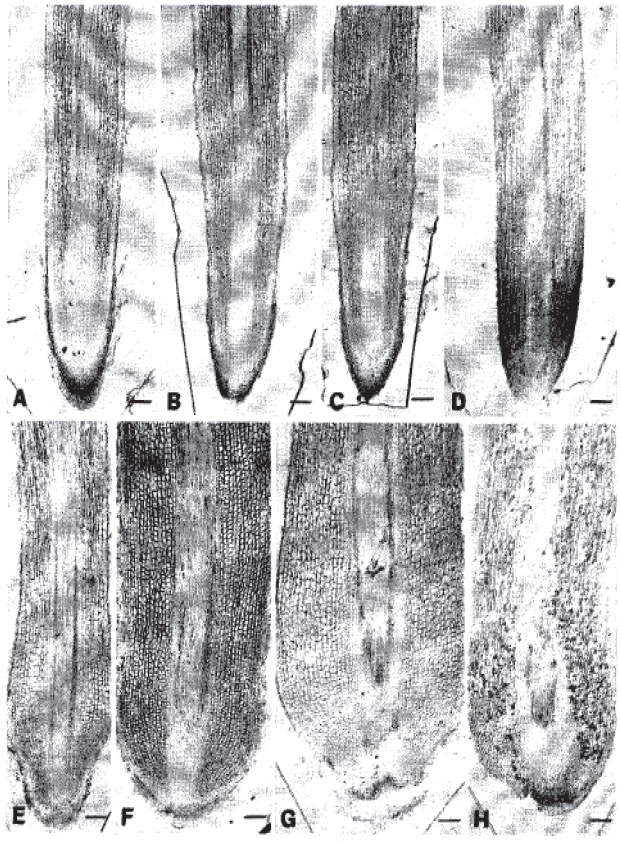 | |
| Fig. 1: | Light micrographs of median longitudinal sections of roots of trifoliate orange budded with satsuma mandarin under aerated and stagnant culture solutions (A, B, C and D indicate aerated while E, F, G and H denote stagnant conditions). A, E: 3-day-induced. B, F: 5-day-induced. C, G: 8-day-induced. D, H: 10-day-induced. Note: Normal anatomy of roots under aerated solution at all exposures but gradual root tip injuries, cell enlargement close to the root apex and finally lysigenous air spacing between the cortical cells under stagnant condition. Bar = 150 μm |
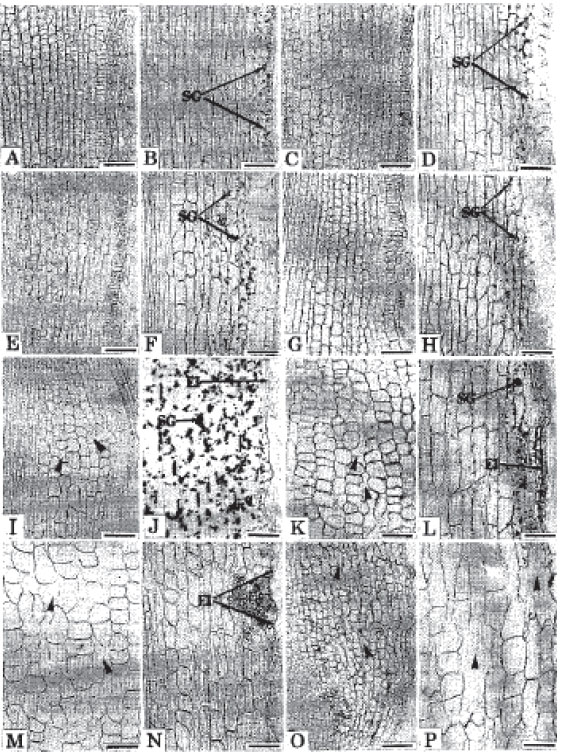 | |
| Fig. 2: | Localized views of longitudinal sections of roots of trifoliate orange budded with satsuma mandarin showing the cellular pattern and starch grain accumulation in the cortical files. (A-H represent aerated condition and I-P denote stagnant solution). A and B: Cells very near and 4 mm apart from the root tip, respectively after 3 days. C and D: Similarly cells of near and apart of root apex after 5 days. E and F: Identically cells from the very near and apart from the root apex after 8 days. G and H: Root tip adjacent and apart cells after 10 days. I and J: Root tip surrounding and 4 mm apart cells after 3 days. K and L: Very near and apart positioned cells after 5 days. M and N: Similarly near and apart positioned cells after 8 days. 0 and P: Identically very near and apart positioned cells after 10 days. Note; No starch grains in the cells close to the root apex but a few grains apart from the apex under aerated solution at all exposures. Initial accumulation of grain in the apart positioned cortical cells followed by complete degradation, cell rounding and air spacing as a gradual manner. SG, Starch grain; El, Epidermal injury, Arrow heads indicate the air spaces. Bar = 100 μm |
This zone became mare shorter and on the 10th day of induction under stagnant condition no meristematic cells were left. The observed data showed that there was a gradual trend of proximity of cell enlargement along the exposure time to the stagnant condition.
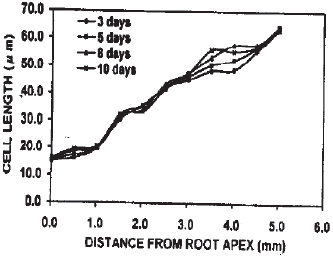 | |
| Fig. 3: | Cell length in the cortical cell files of roots in trifoliate orange budded with satsuma mandarin under aerated condition |
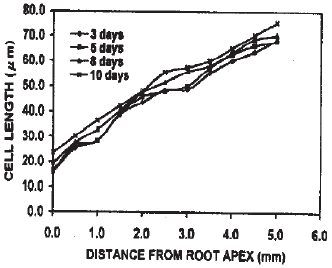 | |
| Fig. 4: | Cell length in the cortical cell files of roots in trifoliate orange budded with satsuma mandarin under stagnant condition |
DISCUSSION
The normal characteristics of roots such as the existence of wider meristematic zone, presence of starch grains in the rootcap cells and well developed column of cells on the rootcap region under aerated solution were distinct as this condition favored the growth of roots even after longer exposure. On the other hand, root tip injuries within a few days of induction to stagnant condition marked that situation as severely adverse. Willey (1970) published root injuries in tobacco under stagnant condition. Therefore, root injury in the present material was consistent with that report. On the contrary, Chapman (1968) reported that citrus is not so sensitive to poor aeration. This variation might be due to the specific material we studied. The injuries under stagnant condition occurred as a gradual manner probably for that reason meristematic zone shortening and cell rounding were also gradually observed. Lunt et al. (1973) published that exposing roots to 0 percent 02 killed Artemisia tridentate and Larrea tridentata in 7 days and Fromseria dumosa in 9 days. In the present study, trifoliate orange roots were under injury and modification of meristematic region but root death was not found. Similarly, Buwalda et al. (1988) stated that membrane integrity of root cells of Triticum aestivum was not adversely affected by 0 percent oxygen for 10 days. The formation of air spaces on the cortex was evidenced gradually under stagnant solution. This formation of air spaces on the cortex resulted through the partial dissolution of cells due to the stress condition and could be marked as the severely adverse effects on the growth of roots. Similar formation of air spaces in the cortical cells of 8 to 10 cm the root tip in corn grown in unaerated nutrient solution have been found (Kramer, 1983). Our study showed that this kind of spaces do not form only in the far apart cortical cells but the periblemic cells might also be under that condition in satsuma mandarin. The secretion of mucilaginous substances by the root tip under stagnant condition also clearly described the adverse effects of this condition.
Starch grains are mainly deposited on the cortical and root cap cells and when the condition is favorable for root growth this deposition became less as was seen by Mohammad and Shiraishi (1998). They found that starch deposition increased in the cortical cells during cold stress and decreased with the resumption of favorable temperature. The slight and constant rate of starch grain deposition under aerated condition was due to normal growth of plants and the much deposition of grains in the preliminary stage under stagnant condition might be due to the resistance of its utilization which was diminished by longer existence of adverse situation.
Cell division under stagnant condition has been reported to be arrested (Berry and Norris Jr., 1949; Williamson, 1968; Summers and Jackson, 1994). Therefore, the shortening of meristematic zone in the present study was probably due to the cessation of cell division in that region. Developing cells are metabolically active, so that in developing tissue a small cell size is necessary for the elevated rates of protein production. Dividing cells may need to be in the 10-20 μm range for protein production. In contrast, intercellular transport is facilitated by longer cells. The meristem must import sucrose and other metabolites from the phloem and the phloem is functional many cell lengths from the meristem (Silk, 1992). In stressed tissue, movement of metabolites from the phloem into the meristem is facilitated by two anatomical adaptations. Slow growth is usually associated with phloem differentiation close to the meristem (Rost and Baum, 1988). The increase in cell length at the base of the meristem also promotes intercellular transport. Apparently the observed cell length patterns help the plant adapt to environmental stress. The enlargement of cells towards the root base might be an stress-induced adaptive mechanism in this plant.
In conclusion, although citrus roots were known to less sensitive to poor aeration, the endurance level is limited by few days. At longer exposures, the roots might be under severe adverse effects like root injury, degradation of stored products, cellular enlargement in the cortex close to the root apex and lysigenous breakdown of cells to form air spaces.
REFERENCES
- Berry, L.J. and W.E. Norris Jr., 1949. Studies of onion root respiration I. Velocity of oxygen consumption in different segments of root at different temperatures as a function of partial pressure of oxygen. Biochimica Biophysica Acta, 3: 593-606.
CrossRefDirect Link - Bertaud, D.S. and P.W. Gandar, 1985. Referential descriptions of cell proliferation in roots illustrated using Phleum pratense L. Bot. Gazette, 146: 275-287.
CrossRefDirect Link - Buwalda, F., C.J. Thomson, W. Steigner, E.G. Barrett-Lennard, J. Gibbs and H. Greenway, 1988. Hypoxia induces membrane depolarization and potassium loss from wheat roots but does not increase their permeability to sorbitol. J. Exp. Bot., 39: 1169-1183.
CrossRefDirect Link - Fox, T.C. and R.A. Kennedy, 1991. Mitochondrial enzymes in aerobically and anaerobically germinated seedlings of Echinochloa and rice. Planta, 184: 510-514.
CrossRefDirect Link - Hoagland, D.R. and T.C. Broyer, 1936. General nature of the process of salt accumulation by roots with description of experimental methods. Plant Physiol., 11: 471-507.
Direct Link - Kennedy, R.A., M.E. Rumpho and T.C. Fox, 1992. Anaerobic metabolism in plants. Plant Physiol., 100: 1-6.
CrossRefDirect Link - Lemon, E.R., 1962. Soil aeration and plant root relations I. Theory. Agron. J., 54: 167-170.
CrossRefDirect Link - Lunt, O.R., J. Letey and S.B. Clark, 1973. Oxygen requirements for root growth in three species of desert shrubs. Ecology, 54: 1356-1362.
CrossRefDirect Link - Mohammad, P. and M. Shiraishi, 1998. Anatomical observations on the accumulation and utilization of storage starch and epidermis-related development in roots of vigorous and non-vigorous satsuma mandarin trees from early winter to early summer. J. Jap. Soc. Hortic. Sci., 67: 660-670.
CrossRefDirect Link - Mohammad, P. and M. Shiraishi, 1999. Root formation, root growth and root morphology in trifoliate orange budded with satsuma mandarin under aerated and stagnant culture solutions. Pak. J. Biol. Sci., 2: 53-59.
CrossRefDirect Link - Nobel, P.S. and J.A. Palta, 1989. Soil O2 and CO2 effects on root respiration of cacti. Plant Soil, 120: 263-271.
CrossRefDirect Link - Nonami, H. and J.S. Boyer, 1989. Turgor and growth at low water potentials. Plant Physiol., 89: 798-804.
Direct Link - Palta, J.A. and P.S. Nobel, 1989. Influence of soil O2 and CO2 on root respiration for Agave deserti. Physiol. Plant., 76: 187-192.
CrossRefDirect Link - Pritchard, J., R.G.W. Jones and A.D. Tomos, 1991. Turgor growth and rheological gradients of wheat roots following osmotic stress. J. Exp. Bot., 42: 1043-1049.
CrossRefDirect Link - Rost, T.L. and S. Baum, 1988. On the correlation of primary root length, meristem size and protoxylem tracheary element position in pea seedlings. Am. J. Bot., 75: 414-424.
Direct Link - Silk, W.K., 1992. Steady form from changing cells. Int. J. Plant Sci., 153: S49-S58.
CrossRefDirect Link - Smit, B. and M. Stachowiak, 1988. Effects of hypoxia and elevated carbon dioxide concentration on water flux through Populus roots. Tree Physiol., 4: 153-165.
CrossRefDirect Link - Steward, F.C., 1935. Mechanism of salt absorption by plant cells. Nature, 135: 553-555.
CrossRefDirect Link - Summers, J.E. and M.B. Jackson, 1994. Anaerobic conditions strongly promote extension by stems of overwintering tubers of Potamogeton pectinatus L. J. Exp. Bot., 45: 1309-1318.
Direct Link - Willey, C.R., 1970. Effects of short periods of anaerobic and near-anaerobic conditions on water uptake by tobacco roots. Agron. J., 62: 224-229.
CrossRefDirect Link - Williamson, R.E., 1968. Influence of gas mixtures on cell division and root elongation of broad bean, Vicia faba L. Agron. J., 60: 317-321.
CrossRefDirect Link - Zhang, F., J.J. Lin, T.C. Fox, C.V. Mujer, M.E. Rumpho and R.A. Kennedy, 1994. Effect of aerobic priming on the response of Echinochloa crus-pavonis to anaerobic stress (Protein synthesis and phosphorylation). Plant Physiol., 105: 1149-1157.
Direct Link - Zieslin, N. and P. Snir, 1989. Responses of rose plants cultivar Sonia and Rosa indica major to changes in pH and aeration of the root environment in hydroponic culture. Scient. Hortic., 37: 339-349.
CrossRefDirect Link