
M. Kashif Riaz Khan
Depatment of plant breading & genetics, university of agriculture, Faisalabad-38040, Pakistan
Abdus salam Khan
Depatment of plant breading & genetics, university of agriculture, Faisalabad-38040, Pakistan
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 2 | Page No.: 340-343







ABSTRACT
A 5 x 5 diallel cross experiment in wheat was conducted to investigate the mechanism of gene action controlling, inheritance of spike length, number of spikelets per spike, number of grains per spike, spike density and grain yield per plant. Highly significant differences were observed among genotypes for all the traits except grain yield per plant for which differences among genotypes were only significant. The graphical representation indicated the additive type of gene with partial dominance for all the characters studied. No epistatic effects were found for all the traits under considerable. From this discussion it could be concluded that selection for yield and its components would be effective in early genera from this material. The genotypes KLR-16 and 6544-6 would be the best promising lines to developed wheat varieties enhanced genetic potential.
PDF Abstract XML References Citation
How to cite this article
M. Kashif Riaz Khan and Abdus salam Khan, 1999. Graphical Analysis of Spike Characters Related to Grain Yield In Bread Wheat (Triticum aestivum L.). Pakistan Journal of Biological Sciences, 2: 340-343.
DOI: 10.3923/pjbs.1999.340.343
URL: https://scialert.net/abstract/?doi=pjbs.1999.340.343
DOI: 10.3923/pjbs.1999.340.343
URL: https://scialert.net/abstract/?doi=pjbs.1999.340.343
INTRODUCTION
Wheat (Triticurn aestivum L.) is the world's most important cereal being the chief source of staple food for about one-third of global population. To cope with the problem of ever increasing population in the country, it appears to be a dire need to enhance the productivity of this cereal grain. So in bread wheat, breeding based on economic traits had been receiving maximum attention for a long time and this factor, has been fully exploited. Hence an estimate of gene action is important in launching a successful breeding programme. Chowdhry et al. (1982), Sharma et al. (1986), Mao and Feng (1987) and Malik et al. (1988) reported that additive gene effect were important for spike length. Khan et al. (1992) from a study of 6 x 6 complete diallel cross reported additive gene action with some degree of partial dominance for spike length.
Khan et al. (1984), Sharma et al. (1986), Prodanovic (1993) and Dimitrijevic et al. (1995) revealed that number of spikelets per spike was conditioned by additive with partial dominance gene action. Malik et al. (1988), Chowdhry et al. (1989) and Khan et al. (1992) studied the gene action for number of grains per spike and found that it was of additive type.
Singh et al. (1985), Chandrakar et al. (1994) and El-Hennawy (1996) reported the grain yield per plant was conditioned by additive gene action with partial dominance. The present studies were carried out to have the information on inheritance pattern of grain yield and spike characters related to it. The information so derived could be very beneficial in the wheat breeding programe carried out for evolution of high yielding wheat varieties.
MATERIALS AND METHODS
The present investigations were carried out in the experimental area of the Department of Plant Breeding and Genetics, University of Agriculture, Faisalabad. The experimental material consisted of five wheat varieties/strains viz., K-65, LU26S, Tob-66, 6544-6, 16.
The varieties/strains were crossed in a complete fashion during Feb-March 1997. Seeds of all crosses with their parents were planted in the field during 1997 for evaluation in a randomized complete block design three replications. Single row of 5 meter length serve an experimental unit. Inter plant and inter-row dista were as 15 and 30 cm, respectively. All other treatment were kept constant for whole the experiment. At guarded plants from each row of each replication randomly selected. The data were recorded on the length (cm), number of spikelets per spike, number of g per spike, spike density and grain yield per plant (g individual plant basis.
Means of ten plants for each character were use (statistical analysis. The data recorded were subject (analysis of variance technique (Steel and Torrie, 1980) where the differences were found significant, means subjected to diallel analysis technique developed by Hayman (1954) and Jinks (1955 and 1956).
RESULTS AND DISCUSSION
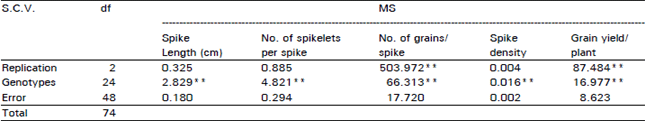
Statistical analysis: Perusal of the Table 1 reveals the genotypes have highly significant differences for all traits under consideration except grain yield per plant which the differences among genotypes were significant.
Spike length: The Fig. 1 showed that regression line of the Wr-axis above the origin indicating the additive gene action with partial dominance. As the regression is of unit slope, it confirms the absence of epistasis. The results seems to be compatible with those Chowdhry et al. (1982), Sharma et al. (1986), Mao and Feng (1987), Malik et al. (1988) and Khan et al. (1992) It is also apparent from the graphic illustration that vi KLR-16 being closest to the origin possessed most of dominant genes while variety K-65 carried most of the recessive genes being farthest from the origin. The variety KLR-16 having maximum array mean (12.3601 cm) appeared to be the best general combiner while variety K-65 having minimum array mean (10.6466 cm) indicated poor combining ability (Table 2).
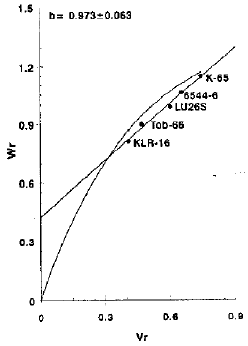 | |
| Fig. 1: | Vr/Wr graph for spike length (cm) |
As the character exhibits additive gene action with partial dominance in the absence of epistasis, the selection in early generations will be beneficial.
Number of spikelets per spike: A reference to Fig. 2 indicated that inheritance pattern for number of spikelets per spike appeared to be of additive with partial dominance, as the regression line intercepts the Wr-axis above the point of origin.
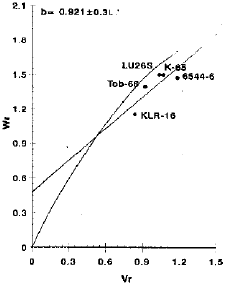 | |
| Fig. 2: | Vr/Wr graph for number of spikelets per spike |
The estimated regression line did not deviate significantly from unit slope, suggesting the absence of non-allelic interaction. The information obtained from the present study also confirms the earlier conclusions of Khan et al. (1984), Sharma et al. (1986), Prodanovic (1993) and Dimitrijevic et al. (1995).
The distribution of varietal array points on the regression line suggested that the variety KLIR-16 possessed maximum dominant genes being closest to the origin while the line 6544-6 being farthest from the origin contained maximum recessive genes (Fig. 2). The variety KLR-16 having maximum array mean (20.48) proved to be the best general combiner while the variety K-65 indicated poor combining ability due to minimum array mean (18.3901) (Table 2). Additive gene action with partial dominance in the absence of epistasis suggests the possibility of obtaining transgressive segregates for this character and selection will be beneficial in early segregating generations.
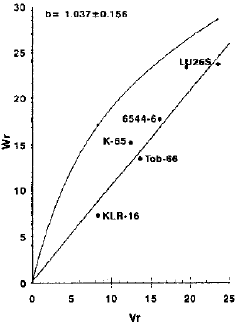 | |
| Fig. 3: | Vr/Wr graph for number of grains per spike. |
Number of grains per spike: The graphical representation of Vr-Wr indicated the additive gene action with partial dominance for this character, as the regression line cuts Wr-axis just above the point of origin. The regression line did not deviate significant from unit slope. The results are in comparison with the findings of Malik et al. (1988), Chowdhry et al. (1989) and Khan et al. (1992).
It is apparent from the graphic illustration that variety KLR-16 being closest to the origin possessed maximum dominant genes where as W265 contained maximum recessive genes being farthest from the origin. The variety Tob-66 has both dominant and recessive genes (Fig. 3).
The variety KLR-16 having maximum array mean (57.84) showed to be the best general combiner while Tab-66 indicated poor combining ability due to minimum array mean 50.4334 (Table 2).
| Table 1: | Analysis of Variance |
 | |
| Table 2: | Array means for different characters |
 | |
Additive gene action with partial dominance in the absence cef epistasis suggests the possibility of obtaining transgressi,,e segregates for this character and selection will be effective in early segregating generations.
Spike density: From Vr.Wr graph for spike density (Fig. 4) it is evident that the regression line cuts the Wr-axis above the origin, indicating partial dominance with additive type of gene action. The estimated regression line did not deviate significantly from unit slope, suggesting the absence of non-allelic interaction. The array points distribution on regression line depicts that variety LU26S being closest to the origin, had maximum dominant genes whereas line 6544-6 possessed maximum recessive genes being farthest from the origin.
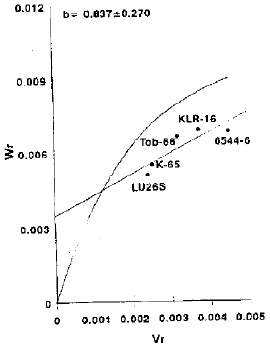 | |
| Fig. 4: | Vr/Wr graph for spike density |
The line 6544-6 having maximum array mean (1.7593) showed to be the best general combiner while the variety Ll.)265 having minimum array mean value (1.6323) exhibited poor combining ability (Table 2). Partial dominance with additive gene action revealed that selection will be beneficial in the early segregating generations.
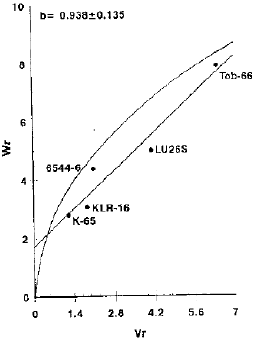 | |
| Fig. 5: | Vr/Wr graph for grain yield per plant (g) |
Grain yield per plant: The graphical representation for grain yield per plant showed that the partial dominance with additive gene action was present for this character as the regression line cuts the Wr-axis above the origin (Fig. 5). The results are in conformity with those of Singh et al, (1985), Chandrakar et al. (1994) and El-Hennawy (1996) As the regression line did not deviate from the unit slope, the absence of epistasis was concluded. From the relative position of array points on the regression line, it is apparent that K-65 showed maximum dominant genes being closest to the origin while variety Tob-66 contained minimum dominant genes being farthest from the origin. The line 6544-6 with maximum array mean (19.6064 g) appeared to be the best general combiner while Tob-66 due to minimum array mean (16.0758 g) showed to have poor combining ability for this trait (Table 2). The genotypes used in the diallel cross provided a broad range of expression for this character. The partial dominance, with additive type of gene action suggested that selection will be beneficial in early segregating generations.
From this discussion it could be concluded that selection for yield and its components would be effective in early generation from this material. The genotypes KLR-16 and 6544-6 would be the best promising lines to developed wheat varieties with enhanced genetic potential.
REFERENCES
- Chowdhry, M.A., A.R. Chowdhry and K. Alam, 1989. Gene action controlling yield and some of its components in spring wheat. Pak. J. Agric. Sci., 26: 451-459.
Direct Link - Jinks, J.L., 1955. A survey of the genetical basis of heterosis in a variety of diallel crosses. Heredity, 9: 223-238.
CrossRefDirect Link - Jinks, J.L., 1956. The F2 and Backcross generations from a set of diallel crosses. Heredity, 10: 1-30.
CrossRefDirect Link - Khan, M.Q., K. Alam and M.A. Chowdhry, 1992. Diallel cross analysis of some morphological traits in spring wheat. Pak. J. Agric. Res., 13: 211-215.
Direct Link - Malik, A.J., A.R. Chaudhry, M.M. Rajpur and K.A. Siddiqui, 1988. General and specific combining ability estimates in spring wheat diallel crosses. Pak. J. Agric. Res., 9: 10-15.
Direct Link - Singh, R.P., R.B. Singh and S. Singh, 1985. Combining ability analysis for yield and its contributing characters in breadwheat. Indian J. Genet. Plant Breed., 45: 539-544.
Direct Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link