
I.A. Hassan
Department of Botany, Alexandria University, 21526 El-shatby, Alexandria, Egypt
S. Anttonen
Department of Ecology and Environmental Science, Univeristy of Kuopio, P.O. Box 1627, Fin-70211, Kuopio, Finland.
M.R. Ashmore
Imperial College Centre for Environmental Technology, Th Huxley School of Environment, Earth Science and Engineering, 48 Prince`s Garddens, London Sw7 2 Pe. Uk.
J.N.B. Bell
Imperial College Centre for Environmental Technology, Th Huxley School of Environment, Earth Science and Engineering, 48 Prince`s Garddens, London Sw7 2 Pe. Uk.
J. Bender
Department of Ecology and Environmental Science, Univeristy of Kuopio, P.O. Box 1627, Fin-70211, Kuopio, Finland.
H.J. Weigel
Institute of Ecotoxicology, Fal, Bundesailee 50, D-38116, Braunschweig, Germany
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 2 | Page No.: 332-335







ABSTRACT
An Egyptian cultivar of wheat (Triticum aestivum L.) was exposed to 60 nl I–1 O3 (8 h d–1) in open-top chambers for 61 d. Ethylene, 1-aminocyclopropane-1-carboxylic acid (ACC), N-malonyl 1-aminocyclopropane-1-carboxylic acid (MACC) chlorophyll contents, visible injury symptoms and yield were measured in this study. O3 was found to increase ethylene, ACC and MACC contents by 22, 29 and 38 percent, respectively, while chlorophyll content and yield were decreased by 29 and 61 per cent, respectively. The potential relationships between these parameters are discussed in the context of O3 induction of accelerated senescence. These results have significance in terms of the maintenance of agricultural yields as O3 rises in Egypt.
PDF Abstract XML References Citation
How to cite this article
I.A. Hassan, S. Anttonen, M.R. Ashmore, J.N.B. Bell, J. Bender and H.J. Weigel, 1999. Effect of O3 on Ethylene Biosynthesis and Yield of Egyptian Cultivar of Wheat (Triticum aestivum L.). Pakistan Journal of Biological Sciences, 2: 332-335.
DOI: 10.3923/pjbs.1999.332.335
URL: https://scialert.net/abstract/?doi=pjbs.1999.332.335
DOI: 10.3923/pjbs.1999.332.335
URL: https://scialert.net/abstract/?doi=pjbs.1999.332.335
INTRODUCTION
Ozone (O3) plays an important role in the earth's atmosphere and changes in its concentrations are of concern for several reasons: increased penetration of UV radiation, a contribution to global warming and direct toxic effects on terrestrial vegetation (Ashmore and Bell, 1991). Ambient concentrations of O3 in Egypt are high enough to cause negative effects on sensitive crops (Hassan et al., 1995). O3 is a strong oxidant, injuring plant tissues and disturbing the anatomical, physiological and biochemical functions of plants (Heath et al., 1984; Nast et al., 1993; Reddy et al., 1993; Mortensen et al., 1995). Although phytotoxic effects of O3 are well known, very little is yet known regarding its possible effects on crop species in Egypt (Elkiey and Ormrod, 1987; Hassan et al., 1994; 1995; Hassan, 1998).
When plants are exposed to O3, ethylene (C2H4) is produced (Taylor et al., 1988; Langebartels et al., 1991; Mehlhorn et al., 1991). It is known that biosynthesis of phytohorrnone ethene (ethylene) can reduce gene expression during fruit ripening, plant senescence and plant stress (Mehlhorn et al., 1991; Reddy et al., 1993; Hippeli and Elstner, 1996). Whether the emission of C2H4 regulates senescence-related responses to O3 awaits further investigations.
Fuhrer and Grandjean (1988) observed that chlorophyll loss (yellowing) of wheat leaves was associated with increases in the synthesis of ethylene and its precursors 1-aminocyclopropan-1-carboxylic acid (ACC) and N-malonyl 1-aminocyclopropane-1-carboxylic acid (MACC) when plants were stressed by O3.
The aim of the present study was to quantify the impact of O3 on Egyptian cultivar of wheat (Triticum aestivum L.) in terms of growth, yield, ethylene and ethylene precursors.
MATERIALS AND METHODS
Plant material and experimental design: Grains of an Egyptian cultivar of wheat (Triticum aestivum L.) were purchased from a commercial source in Alexandria. They were washed with tap water to remove excess fungicides and then imbibed with water over night. Wheat was sown in pots filled with loamy clay soil with pH 7.1 , at about 26/15̊C day/night temperature in a glasshouse in Alexandria in 8/5/96. After emergence, seedlings were transferred in plastic bags to Germany on 29/6/1996 where they were thinned to one plant/pot. Pots were transferred to four open-top chambers (OTCs) of the design of Weight and Jager (1988) on 1/7/1996. There were 15 pots/chamber. Plants were irrigated twice a week with tap water.
Four OTCs were used in this experiment in a split plot design (for more details of 03 fumigation and experimental design (Hassan, 1998).
Visible injury assessment: Leaf injury symptoms we assessed on day 115 of fumigation by counting the number of injured leaves and estimating the percentage of each leaf's area showing chlorotic or necrotic spots (0 ' injury' to 5' 100%' injury).
Destructive harvest and chlorophyll determination: A single destructive harvest was performed at maturity on day 110 after sowing, with measurements of number of ears/plant number of grains/plant, grain weight and 1000-grain weight. Chlorophyll was extracted in 80 percent ethanol and determined on double beam spectrophotometer according to Knudson et al. (1977).
Analysis of ethylene and its biosynthesis: Ethylene and its precursors were determined after 20 minutes of incubation of small pieces of flag leaves (from the final harvest) in closed vials in the dark at room temperature. Use of this period removes any problem caused by the production of wound ethylene which starts to be produced about 25 minutes after excision. Ethylene emission was determined by gas chromatography according to Reddy et al. (1993). For 1-aminocyclopropane-1-1 carboxylic acid (ACC) and N-malonyl ACC (MACC) determinations, wheat leaves were ground to a fine powder in liquid nitrogen and homogenized in 10 cm3 g 80 percent ethanol. The extract was centrifuged at 2800 rpm for 40 min at room temperature and the supernatant dried. The residue was dissolved in 1 cm3 of water, partitioned against 1 cm3 of chloroform and the aqueous phase assayed for ACC according to Langebartels et al. (1991). MACC was determined after neutralization of the extract with NaOH according to Reddy et al. (1993).
Statistical analysis: Data were subjected to one-way analysis of variance based on pot means using STATGIRAPHICS statistical package. There were no covariates used in ANOVA.
RESULTS
O3 was found to increase the number of injured leaves and degree of injury by 3-fold each (Table 1).
| Table 1: | Effect of O3 on visible injury symptoms |
 | |
| FA = Characoal-filtered air | |
| Table 2: | Ethylene, ACC, MACC and chlorophyll content of foliage of wheat plants treated with either 60 nl I–1 O3 or FA |
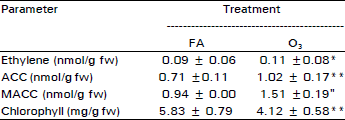 | |
| fw = fresh weight, FA = Characoal-filtered air | |
Ethylene emission increased by 22 percent in O3 in the flag leaves (Table 2). Similar results were observed for ACC and MACC concentrations; their concentrations rose in response to fumigation with 60 nI I–1 O3 by 29 and 38 percent, respectively (Table 2). Table 2 also showed that chlorophyll content decreased by 29 percent after exposure to O3.
Table 3 summarises the effects of O3 on yield and the major yield components. A close examination of this table indicates that the greatest overall contribution to the yield reduction was the reduction in the number of ears/plant (24%). There were larger effects on number of grains/ear (48%) and 1000-grain weight (20%) and this accounted for the large effect on yield (61% reduction) (Table 3).
| Table 3: | Effect of 03 on final yield of wheat |
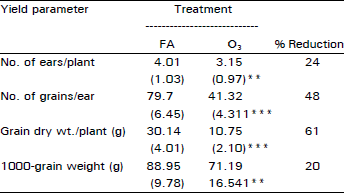 | |
| FA = Characoal-filtered air, Values in parenthesis are±SE | |
DISCUSSION
The present study showed the susceptibility of the cultivar chosen for visible injury symptoms from O3 (both foliar injury parameters were increased by 3-fold each).
The chlorotic and necrotic spots reflect chlorophyll bleaching as a result of reactions involving O2 activation, and this could be a response of physiological and biochemical impairments due to exposure to 03. The percentage reduction in chlorophyll content was highly correlated to the degree of injury (R2 = 0.89) and this is in agreement with the results of Knudson et al. (1977) and Fuhrer and Grandjean (1988).
Exposure to O3 led to enhanced production of ethylene (C2H4), with higher accumulation of ACC and MACC. Excess C2H4 production occurs in response to a number of environmental stresses, including air pollutants (Stan and Schicker, 1982; Fuhrer et al., 1988; Mehlhorn et al., 1991; Reddy et al., 1993; Yin et al., 1994; Schraudner et al., 1994; Hippeli and Elstner, 1996; Finlayson and Reid, 1996). Ethylene is a phytohormone associated with senescence. The increase in C2H4 observed in this study is consistent with many earlier reports in the literature (e.g. Taylor et al., 1988; Langebartels et al., 1991; Yin et al., 1994). O3 could be inducing C2H4 formation through a regulated stimulation of a precursor (e.g. stimulation of ACC synthase leading to formation of ACC). The results of the present study supports this possibility as there was an increase in ACC in wheat plants stressed by O3. Alternatively free radicals a needed to convert ACC to C2H4 and O3 could b mediating this step (Yang and Hoffman, 1984). MACC is formed when there is excess ACC which is not converted to C2H4. In O3-stressed leaves the increase in C2H4, ACC and MACC correlates significantly with the reduction in chlorophyll content (R2 = 0.46, 0.72, 0.67, respectively),(n= 30) and this is consistent with the results of Fuhrer and Grandjean (1988) with wheat leaves and Reddy et al. (1993) with potato leaves.
All yield component parameters measured in this study showed significant reductions following to fumigation with 60 nI I–1 O3 (20-38%) and the reduction in total grain weight were attributed to the combined effects of reductions in the number of ears/plant, number of grains/ear and the 1000-grain weight. These are of great interest in that the results showed the sensitivity to O3 of Egyptian cultivar of wheat and this is consistent with the results of many other workers e.g. Wahid et al. (1995), who reported 20 percent reductions in number of ears/plant, 19 percent reduction in number of grains/ear and 16 percent reductions in 1000-grain weight. Moreover, the reduction in 1000-grain weight is consistent with observations of Heagle et al. (1979), Mulchi et al. (1986), Fuhrer et al. (1989, 1992) and De Temmerman et al. (1992) after exposure to O3.
Reduction in grain dry weight by 38 percent in plants fumigated with 60 nI I–1 O3 compared with plants grown in charcoal-filtered air is considerably greater than nearly all those reported from OTC studies with wheat at similar concentrations in Europe, North America and Pakistan (e.g. Kohut et al., 1987; Fuhrer et al., 1989; De Temmerman et al., 1992; Nussbaum et al., 1995; Wahid et al., 1995). This may be due to difference in cultivar sensitivity, climate or experimental conditions.
The results indicate the potential for substantial yield losses and ethylene emission caused by O3, in a region where yields needed to be increased to feed the rapidly increasing population and where pollution emissions are also increasing rapidly and this must be a matter of great concern. If such impacts are widespread, the implications for agricultural production are considerable not only for Egypt but also for many other developing countries. Moreover, the concentration used in this experiment (60 nl I–1) was slightly lower than that recently recorded in rural area in Egypt (99 nl I–1, as annual mean) (Hassan, 1999). Since the O3 level is likely to continue to rise over the coming decades in many developing countries, it is clear that its potential impacts on agriculture need urgent consideration. Moreover, wheat plant has been the major crop plant to establish critical levels for O3 (level I) in recent years, and in the present study 20000 nI I–1 h 03 in total was applied which is far above AOT 40 level of 3000 nl I–1 h. It would have been interesting in future investigation to determine biochemical parameters, symptoms and growth at the time point when the dose of 3000 nI I–1 h was recorded.
ACKNOWLEDGMENTS
This work was jointly supported with fellowship visit. The Royal Society of London and C1M0 of Finland I.A.Hassan. Part of this project was supported with D.A scholarship to IAH. We would like to express our thanks Professor S. Barakat (Faculty of Science, Alexandria University) for his critical suggestions and revising manuscript. Our special thanks to Prof. H. Helal at FA his help in biochemical analysis.
REFERENCES
- Ashmore, M.R. and J.N.B. Bell, 1991. The role of ozone in global change. Ann. Bot., 67: 39-48.
CrossRefDirect Link - De Temmerman, L., K. Vandermeiren and M. Guns, 1992. Effects of air filtration on spring wheat grown in open-top field chambers at a rural site. I. Effects on growth, yield and dry matter partitioning. Environ. Pollut., 77: 1-5.
CrossRefDirect Link - Elkiey, T. and D.P. Ormrod, 1987. Casuarina and Eucalyptus response to single and multiple gaseous air pollutants. Water Air Soil Pollut., 36: 365-370.
CrossRefDirect Link - Finlayson, S.A. and D.M. Reid, 1996. The effect of CO2 on ethylene evolution and elongation rate in roots of sunflower (Helianthus annuus) seedlings. Physiologia Plantarum, 98: 875-881.
CrossRefDirect Link - Fuhrer, J., A. Egger, B. Lehnherr, A. Grandjean and W. Tschannen, 1989. Effects of ozone on the yield of spring wheat (Triticum aestivum L., cv. Albis) grown in open-top field chambers. Environ. Pollut., 60: 273-289.
CrossRefDirect Link - Fuhrer, J., A.G. Grimm, W. Tschannen and H. Shariat-Madari, 1992. The response of spring wheat (Triticum aestivum L.) to ozone at higher elevations. II. Changes in yield, yield components and grain quality in response to ozone flux. New Phytol., 121: 211-219.
CrossRefDirect Link - Hassan, I.A., 1999. Air pollution in the Alexandria region, Egypt-I: An investigation of air quality. Environ. Educ. Inform., 18: 67-78.
Direct Link - Hassan, I.A., M.R. Ashmore and J.N.B. Bell, 1994. Effects of O3 on the stomatal behaviour of Egyptian varieties of radish (Raphanus sativus L. cv. Baladey) and turnip (Brassica rapa L. cv. Sultani). New Phytol., 128: 243-249.
CrossRefDirect Link - Hassan, I.A., M.R. Ashmore and J.N.B. Bell, 1995. Effect of ozone on radish and turnip under Egyptian field conditions. Environ. Pollut., 89: 107-114.
CrossRefDirect Link - Heagle, A.S., S. Spencer and M.B. Letchworth, 1979. Yield response of winter wheat to chronic doses of ozone. Can. J. Bot., 57: 1999-2005.
CrossRefDirect Link - Hippeli, S. and E.F. Elstner, 1996. Mechanisms of oxygen activation during plant stress: Biochemical effects of air pollutants. J. Plant Physiol., 148: 249-257.
CrossRefDirect Link - Kohut, R.J., R.G. Amundson, J.A. Laurence, L. Colavito, P. van Leuken and P. King, 1987. Effects of Ozone and sulfur dioxide on yield of winter wheat. Phytopathology, 77: 71-74.
CrossRefDirect Link - Knudson, L.L., T.W. Tibbitts and G.E. Edwards, 1977. Measurement of ozone injury by determination of leaf chlorophyll concentration. Plant Physiol., 60: 606-608.
CrossRefDirect Link - Langebartels, C., K. Kerner, S. Leonardi, M. Schraudner, M. Trost, W. Heller and H. Sandermann Jr., 1991. Biochemical plant responses to ozone. I. Differential induction of polyamine and ethylene biosynthesis in tobacco. Plant Physiol., 95: 882-889.
CrossRefDirect Link - Mehlhorn, H., J.M. O'Shea and A.R. Wellburn, 1991. Atmospheric ozone interacts with stress ethylene formation by plants to cause visible plant injury. J. Exp. Bot., 42: 17-24.
CrossRefDirect Link - Mortensen, L., A. Bastrup-Birk and H. Ro-Poulsen, 1995. Critical levels of O3 for wood production of European beech (Fagus sylvatica L.). Water Air Soil Pollut., 85: 1349-1354.
CrossRefDirect Link - Mulchi, C.L., D.J. Sammons and P.S. Baenziger, 1986. Yield and grain quality responses of soft red winter wheat exposed to ozone during anthesis. Agron. J., 78: 593-600.
CrossRefDirect Link - Nast, W., L. Mortensen, K. Fischer and I. Fitting, 1993. Effects of air pollutants on the growth and antioxidative system of Norway spruce exposed in open-top chambers. Environ. Pollut., 80: 85-90.
CrossRefDirect Link - Nussbaum, S., M. Geissmann and J. Fuhrer, 1995. Effects of nitric oxide and ozone on spring wheat (Triticum aestivum). Water Air Soil Pollut., 85: 1449-1454.
CrossRefDirect Link - Reddy, G., R.N. Arteca, Y.R. Dai, H.E. Flores, F.B. Negm and E.J. Pell, 1993. Changes in ethylene and polyamines in relation to mRNA levels of the large and small subunits of ribulose bisphosphate carboxylase/oxygenase in ozone‐stressed potato foliage. Plant Cell Environ., 16: 819-826.
CrossRefDirect Link - Schraudner, M., U. Graf, C. Langebartels and H. Sandermann, 1994. Ambient ozone can induce plant defence reactions in tobacco. Proc. R. Soc. Edinburgh Sect. B: Biol. Sci., 102: 55-61.
CrossRefDirect Link - Stan, H.J. and S. Schicker, 1982. Effect of repetitive ozone treatment on bean plants-stress ethylene production and leaf necrosis. Atmos. Environ., 16: 2267-2270.
CrossRefDirect Link - Taylor, Jr. G.E., B.M. Ross-Todd and C.A. Gunderson, 1988. Action of ozone on foliar gas exchange in Glycine max L. Merr: A potential role for endogenous stress ethylene. New Phytol., 110: 301-307.
CrossRefDirect Link - Wahid, A., R. Maggs, S.R.A. Shamsi, J.N.B. Bell and M.R. Ashmore, 1995. Air pollution and its impacts on wheat yield in the Pakistan Punjab. Environ. Pollut., 88: 147-154.
CrossRefDirect Link - Yang, S.F. and N.E. Hoffman, 1984. Ethylene biosynthesis and its regulation in higher plants. Ann. Rev. Plant Physiol., 35: 155-189.
CrossRefDirect Link - Yin, Z.H., C. Langebartels and H. Sandermann, 1994. Specific induction of ethylene biosynthesis in tobacco plants by the air pollutant, ozone. Proc. R. Soc. Edinburgh Sect. B: Biol. Sci., 102: 127-130.
CrossRefDirect Link