
C. Akem
Germplasm Program, International Center for Agricultural Research in the Dry Areas (ICARDA), P.O. Box 5466, Aleppo, Syria
S. Kabbabeh
Germplasm Program, International Center for Agricultural Research in the Dry Areas (ICARDA), P.O. Box 5466, Aleppo, Syria
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 2 | Page No.: 277-279







ABSTRACT
In an annual monitoring visit of chickpea (Cicer arietinum L.) fields for diseases occurring on promising chickpea breeding lines in Syria, Sclerotinia stem rot caused by Sclerotinia sclerotiorum was found in more than 30 percent of the fields in which the trials were planted. The disease incidence ranged from 5 to 100 percent and none of the lines in the trials showed high resistance to the disease. A program was initiated to start screening chickpea lines to Sclerotinia stem rot with known resistance to Fusarium wilt caused by Fusarium oxysporum f. sp. ciceris, to identify lines with combined resistance. A detached shoot technique was used to determine the preliminary reaction of chickpea genotypes to infection by S. sclerotiorum. Results using this technique under controlled conditions in the growth chamber, showed that out of 15 chickpea genotypes evaluated, 5 exhibited some resistance to Sclerotinia stem rot. This was shown by delayed initial infection, restricted lesion development and no sclerotial production. These results were confirmed using potted plants in the plastic house. The technique is simple, less labor-intensive and requires a very short period of time to obtain results that can be confirmed under field seasonal or other conditions.
PDF Abstract XML References Citation
How to cite this article
C. Akem and S. Kabbabeh, 1999. Screening for Resistance to Sclerotinia Stem Rot in Chickpea: A Simple Technique. Pakistan Journal of Biological Sciences, 2: 277-279.
DOI: 10.3923/pjbs.1999.277.279
URL: https://scialert.net/abstract/?doi=pjbs.1999.277.279
DOI: 10.3923/pjbs.1999.277.279
URL: https://scialert.net/abstract/?doi=pjbs.1999.277.279
INTRODUCTION
Sclerotinia stem rot caused by Sclerotinia sclerotiorum (Lib.) De Bary is an important disease of legumes and other crops with a worldwide distribution. It is particularly severe on beans (Phaseolus vulgaris L.) and soybean (Glycine max Merr.) (Boland and Hall, 1987; Coyne et al., 1977). Symptoms of the disease on these crops include flagging, wilting, necrosis and relatively dry lesions on stems, stalks or branches with demarcations between healthy and diseased tissue (Purdy, 1979). Under moist humid conditions, white fluffy mycelium appears on the base of diseased plants. Infected branches become chlorotic and eventually die. The pathogen can produce many scierotia on the surface and within infected stems (Melouk et al., 1992). S. sclerotiorum is also the primary cause of stem rot of chickpea (Cicer arietinum L.) (Nene, 1980) and is one of the pathogens associated with the root rot complex of chickpea (ICARDA, 1994). Its occurrence is increasing in both incidence and severity on chickpea grown in the Mediterranean region. This increase has been aggravated by the cultivation of the crop during the winter months when conditions favouring plant growth also favour the development of the disease.
In recent surveys to monitor the prevalence of diseases occurring on winter chickpea trials in Syria, Sclerotinia stem rot was second only to Ascochyta blight (Ascochyta rabiei) (ICARDA, 1996). It was found in more than 20 percent of the 53 fields surveyed, with disease incidence ranging from 5 to 100 percent. There are also reports that this disease is becoming increasingly important on irrigated chickpea in Egypt (Abou-Zeid et al., 1997).
The recent observations suggested that Sclerotinia stem rot could become a major disease affecting winter chickpea in the Mediterranean region as the area under cultivation expands. There was thus a need to start considering options of control for this disease. Host-plant resistance has often been the preferred option to address control for new diseases. This is because it is often effective, usually economic and considered to be environmentally friendly. Some research was therefore initiated to screen for possible sources of resistance to this potentially important disease of chickpea in the available germplasm at ICARDA. The main objective of the research was to use a small number of genotypes with known resistance to other important diseases such as Ascochyta blight and Fusarium wilt (Fusarium oxysporum f. sp. ciceris) in order to develop and test a suitable simple screening technique.
MATERIALS AND METHODS
A detached shoot-screening technique was tested to determine the reaction of 15 chickpea genotypes to an isolate of S. sclerotiorum collected from a diseased chickpea plant in the field. This technique is similar to the one developed and used effectively to screen peanut genotypes for resistance to S. minor Jagger (Melouk et al., 1992) and soybean to Sclerotium rolfsii Saco (Akem and Dashiell, 1991). Chickpea plants were grown in pots in the greenhouse for 6 weeks and 10 shoots, 12-14 cm long were detached from each genotype for inoculation.
Individual shoots were supported by a cotton plug and the cut ends were immersed individually in 1x14 cm test tubes containing Hoagland solution. Each shoot was inoculated by placing a 4 mm mycelial plug of S. sclerotiorum from a 2-day old culture grown on potato dextrose agar (PDA), at the axil between the stem and the petiole at about the mid-portion of the shoot. Inoculated shoots were put on metal racks and placed in a growth chamber at 25-27°C and 80-90 percent relative humidity. Shoots were completely randomized after inoculation before placement on the racks. To confirm the detached shoot reactions, 10 plants of each genotype were inoculated by placing a mycelial plug of S. sclerotiorum on the lowest axil towards the base of the plants in the pots. Each pot containing 5 inoculated plants was covered with a transparent polyethylene bag for 24 hrs. to increase initial relative humidity to above 8 percent. Pots were completely randomized on the plastic house benches after inoculations. The potted plants were evaluated for disease incidence 14 days after inoculation. Only plants with measurable lesions were considered infected.
Lesion development on the shoots was measured from day 3 after inoculation through day 8 when some of the shoots were completely colonized with mycelia of S. sclerotiorum. Mean lesion lengths on each genotype were calculated as the sum of individual lesion lengths divided by the total number of inoculated shoots.
Upon conclusion of lesion measurements, the relative humidity in the chambers was lowered to 60 percent and the Hoagland solution was drained from the test tubes. Tubes with infected shoots were left in the chamber for two weeks to allow for possible sclerotial formation. The number of shoots within each genotype with sclerotial formation were noted. Mean length of lesions was linearly regressed against time after inoculation to determine the rate of lesion expansion, where the slope of the line represented the rate of lesion expansion (cm/day) on each genotype. Analysis of variance was carried out to determine the least significant differences between lesion expansion rates in the detached shoots and disease incidence in the inoculated potted plants among the genotypes.
RESULTS AND DISCUSSION
The following types of lesions were observed on inoculate shoots: 1) small superficial lesions, less than 0.2 cm length, generally restricted to the point of contact of inoculum and stem; 2) rapidly expanding lesions restricte to one side of the stem; and 3) rapidly expanding lesion that completely girdled the stem with the fungus activel colonizing the entire shoot.
In the most susceptible reaction, the first symptoms of infected shoots were water soaked lesions that starte forming at the points of contact of the pathogen and the stem two days after inoculation. These water soake lesions were not readily observed on the other reaction groups with one side and point infections. These lesions expanded rapidly in the susceptible lines and completed girdled the stems within 72 hrs., causing wilting. Stem girdling was observed on 9 genotypes which were classified as highly susceptible. Genotypes that showed moderat susceptibility were girdled slowly, starting with infection of one side and wilting was accordingly delayed. This was observed on ILC 190, F 86-68 and F 92-66. Lesions did not develop beyond points of contact of inoculum an stems on some genotypes. These points were restricted on confined and no further lesion expansion was observed These reactions were noticed on stems of ILC 200 and 84-18. Table 1 shows the average length of the lesions of the various chickpea genotypes at three different times of measurements after inoculation with S. sclerotiorum. The rates of lesion expansion as determined by slopes of regression lines for all the genotypes were also compare (Table 1). Five genotypes (F 86-68, F 84-18, F 85-7, ILC 200 and F 92-66) had significantly lower infection rates than the susceptible genotypes in the trial.
| Table 1: | Average lesion length per shoot, lesion expansion rate, sclerotial formation and disease incidence on potted plant of 15 chickpea genotypes inoculated with S. sclerotiorum |
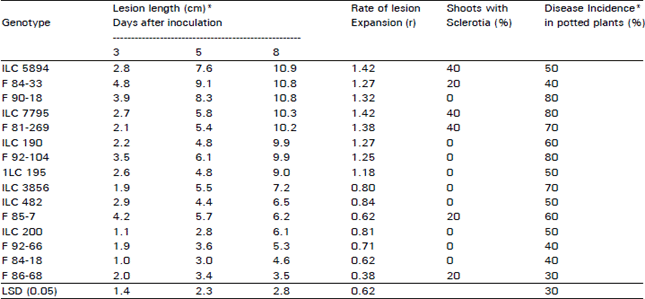 | |
| *Averages from 10 shoots per genotype; **Incidence scores taken 14 days after inoculations from 10 plants | |
Genotypes with the least number of infected stems also had the lowest rates of lesion expansion.
Not all infected stems produced sclerotia (Table 1). The genotype F 90-18, for example, with a high lesion expansion rate did not produce any sclerotia, while F 86-68 with a low lesion expansion rate produced sclerotia. There was a low (r = 0.38) but positive and significant (p<0.05) correlation between lesion expansion rate and sclerotial production. This is in agreement with results obtained from the reaction of peanut genotypes to S. minor (Melouk et al., 1992) evaluated under similar conditions where a high correlation between lesion expansion rate and sclerotial production was observed.
The results obtained showed that most of the 15 chickpea genotypes evaluated were susceptible to S. sclerotiorum. Few genotypes, however, showed moderate levels of resistance to the pathogen which was confirmed by inoculation of potted plants in the greenhouse. Among these were F 84-18, F 92-66 and 1LC-200, which had a low lesion expansion rate, produced no sclerotia and had a relatively low disease incidence in the confirmation test with potted inoculated plants (Table 1).
The detached shoot technique described and used in these evaluations is a rapid procedure for preliminary screening of chickpea genotypes for resistance to S. sclerotiorum under controlled conditions. It may also be used in screening segregating populations for resistance to the disease in a breeding program. It is simple, less labor-intensive and requires a very short period to obtain results, as compared to field seasonal evaluations. Several shoots can be obtained for evaluation from a single plant, thus maximising the utility and economising on plant material. The technique should not, however, be used as a substitute for field evaluations because some genotypes may react differently under controlled and field conditions. This may be due, among other factors, to canopy effects that may influence reactions and to multiple infections occurring throughout the cropping season.
REFERENCES
- Akem, C.N. and K.E. Dashiell, 1991. Detached shoot technique to evaluate the reaction of soybean cultivars to Sclerotium rolfsii. Crop Prot., 10: 325-327.
Direct Link - Boland, G.J. and R. Hall, 1987. Evaluating soybean cultivars for resistance to Sclerotinia sclerotiorum under field conditions. Plant Dis., 71: 934-936.
Direct Link - Purdy, L.H., 1979. Sclerotinia sclerotiorum: History, diseases and symptomatology, host range, geographic distribution and impact. Phytopathology, 69: 875-880.
CrossRefDirect Link