
Nazeer Hussain Shah
Agricultural Research Institute, D.I.Khan, Pakistan
G.M. Paulsen
Department of Agronomy, Kansas State University, Manhattan, Kansas, 66506, USA
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1286-1291







ABSTRACT
The decline in photosynthesis and rise in stomas resistance during maturation were slowed by low temperature and speeded by high temperature and damage from osmotic stress increased substantially with rise in temperature. Viable leaf area persisted longer and was diminished less by osmotic stress under low temperature than under high temperature. Vegetative and grain masses at maturity were increased by low temperature and were reduced most by osmotic stress with high temperature. The results demonstrate that high temperature and osmotic stress reduce photosynthesis and productivity and interact synergistically to accentuate injury. Wheat can tolerate considerable osmotic stress if the temperature is favorable because of a direct effect that maintains photosynthesis and an indirect effect that minimizes injury from osmotic stress.
PDF Abstract XML References Citation
How to cite this article
Nazeer Hussain Shah and G.M. Paulsen, 1999. Response of Wheat to Combined High Temperature and Osmotic Stresses During Maturation. 1: Plant Photosynthesis and Productivity. Pakistan Journal of Biological Sciences, 2: 1286-1291.
DOI: 10.3923/pjbs.1999.1286.1291
URL: https://scialert.net/abstract/?doi=pjbs.1999.1286.1291
DOI: 10.3923/pjbs.1999.1286.1291
URL: https://scialert.net/abstract/?doi=pjbs.1999.1286.1291
INTRODUCTION
Plant productivity is severely limited by environmental stresses, particularly high temperature and drought through out the world (Schonfeld et al., 1988). Extensive investigations has documented their injurious effects on photosynthesis (Frank et al., 1973; Harding et al., 1990a; Johnson and Moss, 1976; Schonfeld et al., 1988), but the stresses have been rarely studied concurrently. This is unfortunate, because high temperature and drought commonly coincide (Gusta and Chen, 1987) and their combined effects are largely unknown. High temperature injury to wheat damages photo-system-II, the most labile process, before other are affected and decreases photosynthesis irreversibly (Harding et al., 1990a; Sayed et al., 1989a,b). Senescence of leaves also is induced rapidly, lowering photosynthetic rate and reducing active leaf area Al-Khatib and Paulsen, 1989). Injury from high temperature is particularly detrimental during the grain filling period because photosynthesis provides most of the kernel mass (Sofield et al., 1977). Initial growth of the wheat kernel is promoted because high temperature promotes mobilization of assimilates from vegetative parts to the grain (Tashiro and Wardlaw, 1990). Duration of grain growth is curtailed severely, an estimated 33 percent to 50 percent, as the temperature increases from 15 to 35°C (Wardlaw et al., 1980). Early cessation of grain filling has little effect on kernel number but seriously reduces kernel mass and density and, thus, the total grain yield (Tashiro and Wardlaw, 1990).
Osmotic stress decreases stomata( conductance and photosynthesis of wheat during onset of drought (Shimshi et al., 1982). Prolonged stress further damages photosynthetic processes, particularly photochemical reactions (Ehdaie et al., 1988), as tissue desiccate. Senescence is affected little by mild stress, but may be accelerated by severe stress (Lu et al., 1989). Translocation of assimilates from vegetative to reproductive parts, on the other hand, continues at water deficit levels that inhibit photosynthesis (Johnson and Moss, 1976). Kernel growth is reduced more than other yield components of wheat by drought during grain filling stage (Fischer and Maurer, 1978). Reasons for the reduction are not clear; however, both an inadequate supply of assimilates from vegetative sources (Wardlaw, 1967) and direct effect of osmotic stress on storage processes in reproductive sinks (Nicolas et al., 1984) have been implicated. It might be relevant that drought stress, at least during early stage of grain development, greatly decreases leaf water status but has little or no effect on spike water potential (Gusta and Chen, 1987).
Combined effects of high temperature and drought must be investigated to understand how two stresses interact when they coincide in nature. Previous studies of the kernel sink have shown a strong synergy between effects of high temperature and drought on cell formation and growth (Nicolas et al., 1984). Similar studies of the assimilate sources are lacking, however objectives of our investigations were to compare effects of high temperature and osmotic stress during grain filling and to ascertain interactions between these stresses on wheat photosynthesis and productivity.
MATERIALS AND METHODS
Plant culture and treatment: The study was conducted in 1990 at Kansas State University. Seeds of spring wheat were germinated in moist vermiculite. Seven-day-old seedlings were transplanted to 2L opaque containers, each holding four plants and grown on hoagland solution in a greenhouse until anthesis. The solutions were continuously aerated, adjusted to pH 5.0 semi-weekly and changed weekly. Greenhouse temperature was maintained near 25/15°C day/night and irradiance at noon averaged 1400 μmol m2 S–1 PAR. Humidity was not controlled.
Tillers were individually labelled by date of anther extrusion and plants were transferred to controlled environment chambers when anthers appeared in 50 percent of the spikes. Plants were briefly acclimated at 22/17°C day/night and then were randomly assigned among six treatments consisting of three temperature regimes, 15/10, 25/20, or 35/30°C, combined with two osmotic stress levels, -0.01 MPa from the nutrient solution or -0.40 MPa from polyethylene glycol (PEG 8000). One half of the PEG was added when temperature treatments were applied and the other half was added 4 days later to all osmotically stressed plants except those at 35/30°C, which were wilted severely. The concentration of PEG required was individually calculated for each temperature regime (Michel and Kaufmann, 1973).
All chambers had 16/8-hr light/dark regimes with 420 μmol m–2 s–1 PAR during light periods. Humidity was not controlled and ranged from 40 to 50 %RH during the light period and from 70 to 80 percent RH during the dark period under all temperature regimes.
Photosynthesis, leaf area and plant mass measurements: Plant photosynthetic parameters, leaf areas and mass of vegetative parts and grain were measured when temperature and osmotic treatments were imposed and at weekly intervals afterwards until maturity.
Photosynthetic rates and stomatal conductances were measured with a LI-6200 system (Li-Cor, Inc., Lincoln, NE, USA). Readings were taken on flag leaves of main stems of at least three plants in each treatment in three replications on each date.
After photosynthetic parameters were measured, plants were separated into viable and nonviable leaf blades by visual assessment, stems plus sheaths and spike components. Area of viable leaves was measured with a LI3000 meter (Li-Cor,lnc.). Vegetative parts were dried at 70°C to constant weight, usually for 72 hr and weighed. Spikes were dried at 40°C constant weight, usually for 7 d and threshed manually or with a mechanical thresher (Precision Manufacturing company, Lincoln, NE, USA) and the grain was redried at 40°C and weighed.
Mean grain filling rate from anthesis to physiological maturity was calculated as the average incremental change in grain mass per day during each 7-d sampling interval. The grain filling duration was calculated as the ratio of final grain weight to mean grain filling rate (Sofield et al., 1977).
Experimental design and data analyses: A completely randomized design was used. Pots were randomly assigned to the six combination of temperature and osmotic stress and treatments were replicated three times. Differences among treatment means were compared by least significant differences (LSD) at the 0_05 or 0.10 probability level when sample sizes were equal. Least square means were used when sample sizes were not equal.
RESULTS AND DISCUSSION
Plants grown at 15/10°C without osmotic stress exhibited few signs of senescence except for partial chlorosis of lower most leaves and spikes. Most leaves were still viable when the grain was physiologically mature with a moisture content of ca. 25 percent at 70 days after anthesis. Plants at 25/20°C senesced faster and leaves and spikes were completely chlorotic after 49 days. At 35/30°C, all plant parts matured within 7 days after anthesis.
Osmotic stress hastened senescence less at low temperature than at high temperature. The flag and penultimate leaves of plants at 15/10°C with osmotic stress remained viable and the stem was chlorophyllous when the grain reached physiological maturity after 63 d. All leaves of plants at 25/20°C senesced after 28 days and stems were chlorotic after 35 days. Plants at 35/30°C with only one half of PEG osmoticum lost all viability within 4 days. Flag leaf photosynthetic rates decreased under all temperatures during the first 7 days of treatment when plants were not osmotically stressed (Fig. 1). The decline was significantly slower at 15/10°C and 25/20°C than at 35/30°C. Activity remained high at 15/10°C and persisted until 70 days after anthesis. Similar rates were maintained at 25/20°C for 35 d, when activity began to decrease to low levels at 49 days. Plants at 35/30°C lost all activity within 7 days.
Osmotic stress decreased photosynthetic rates at all temperature regimes except the highest during the first 7 days (Fig. 1). Rates continued lower with osmotic stress than without osmotic stress at 15/10°C, but photosynthesis persisted for 63 days. Combining osmotic stress with 25/20°C ended photosynthetic activity after 21 days. At the highest temperature, all activity ceased within 7 days regardless of osmotic stress.
Stomatal resistance was low throughout the experiment when plants were grown at 15/10°C without osmotic stress (Fig. 2). Increasing the temperature to 25/20°C increased, stomatal resistance slightly after 42 days as leaves began to senesce, but values were still low. The highe temperature, 35/30°C, raised stomatal resistance within days.
Combining osmotic stress with 15/10°C temperature did not alter stomatal resistance until 49 days, when values increased slightly (Fig. 2). When osmotic stress was combined with 25/20°C treatment, however, stomata! resistance increased after 14 days and with 35/30°C treatment, it increased markedly the first 7 days.
Viable leaf area at 15/10°C remained constant during the initial 14 days, decreased until 28 days and then stayed nearly steady until 70 days after anthesis when plants were not osmotically stressed (Fig. 3). At 25/20°C, leaf area decreased the first 7 days, remained constant the next 7 days and then fell until leaves lost viability after 42 days. High temperature accelerated senescence greatly and caused all the leaves to lose viability within 7 days.
Osmotic stress at low temperature decreased leaf area rapidly for 14 days, after which senescence slowed, and some viable leaf area was still present 63 days after anthesis (Fig. 3). At the intermediate temperature, osmotic stress decreased most of the viable leaf area within 7 days and all of it after 21 days. All leaves senesced completely within 7 days at the high temperature regardless of osmotic stress.
Plant (shoot) mass increased equally under all temperature treatments for the first 7 days (Fig. 4). Growth continued steadily at the low temperature without osmotic stress to a maximum after 70 days of treatment but ended at the intermediate temperature after 14 days and at the high temperature after 7 days. Osmotic stress slowed the plant growth rate and shortened the growth duration to 63 days at 15/10°C. No additional growth was added after the first 7 days when osmotic stress was combined with 25/20 or 35/30°C treatments.
Grain mass increased slowly but steadily and reached the highest final yield after 70 days when plants were grown at 15/10°C without osmotic stress (Fig. 5). Increasing the temperature to 25/20°C enhanced grain growth the first 28 days, but no additional accumulation occurred after that time. Plants at 35/30̊C produced little grain.
Osmotic stress did not consistently affect early grain growth at 15/10°C (Fig. 5). Grain growth slowed after 35 days and ended after 63 days at the low temperature. At the intermediate temperature, osmotic stress stopped grain growth after 14 days and at the high temperature, caused rain growth to end immediately.
Mean kernel weight was the only yield component that was affected by temperature and osmotic stress treatments (Fig. 6). Spike number and kernels per spike were already set when treatments were imposed and were not altered. Kernel growth at 15/10°C without osmotic stress followed the same slow pace as grain yield for 49 days, increasing the temperature to 25/20°C likewise inceased kernel growth the first 14 days, but accumulation of assimilates slowed until 35 days and then ended after 42 days, Kernel growth was slight at 35/30°C and ended soon ter anthesis.
Early growth of kernels was affected inconsistently by osmotic stress at 15/10°C (Fig. 6). At 25/20°C, however, growth ended after only 14 days when plants were osmotically stressed. The severe injury to kernel growth from the 35/30°C treatment was not increased by osmotic stress.
The mean rate of grain filling over each 7-days period increased from the low to the intermediate temperature and then decreased at the high temperature without osmotic stress. The positive response between 15/10 and 25/20°C, but not the negative response to 35/30°C was prevented by osmotic stress.
Mean grain filling duration decreased by nearly one-half from the low to the intermediate temperature and again from the intermediate to the high temperature when plants were not osmotically stressed. Osmotic treatment shortened the long duration of grain filling at the low temperature but did not significantly alter the briefer durations at the intermediate and high temperatures.
Wheat growth and production are severely affected by high temperature and water stress during maturation (Ehdaie et al., 1988). Drought is commonly considered to be the major factor limiting wheat productivity in the world (Gusta and Chen, 1987). It is increasingly evident, however, that high temperature, particularly during the grain filling stage, also greatly reduces yield (Al-Khatib and Paulsen,1990; Harding et al., 1990b; Wardlaw et al., 1980). High temperature also combines with drought to diminish grain development more than either stress alone. The present study demonstrates that high temperature and osmotic stress interact even more markedly to alter plant photosynthesis and leaf area duration, the primary determinants of wheat productivity.
Low temperature enabled plants to maintain photosynthesis and the supply of assimilates for grain filling largely independently of osmotic stress. Considering that current photosynthesis provides most of the assimilates for high yield (Evans and Rawson, 1970; Evans et al., 1975). It is apparent that wheat can tolerate substantial osmotic stress if the temperature is favorable. Thus, low temperature has at least two benefits: a direct effect that maintains high rate of photosynthetic activity and an indirect effect that minimizes adverse consequences of osmotic stress.
Net photosynthesis, mesophyll conductivity and chlorophyll content, a measure of leaf viability, are highly correlated in wheat (Planchon, 1979). Comparisons of some wheat and triticale species indicated that mesophyll conductivity was the major limiting factor to CO2 exchange by photosynthesis. The response of wheat to high temperature and osmotic stress treatments in our experiments does not support extrapolating genotypic effects to environmental effects. Reduced photosynthetic rates from high temperature, in particular, were not associated with low stomatal conductivity, a result also obtained in other research (Al-Khatib and Paulsen,1990).
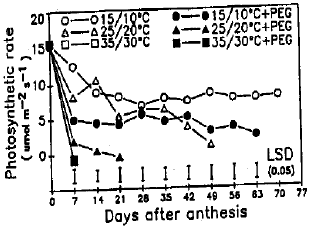 | |
| Fig. 1: | Photosynthetic rate of spring wheat as affected by various temperatures and PEG 800 with time |
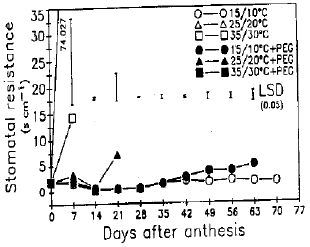 | |
| Fig. 2: | Stomatal resistance of spring wheat as affected by various temperatures and PEG800 with time |
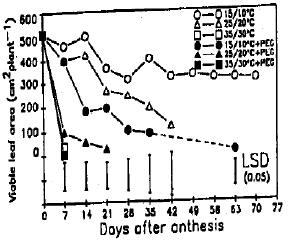 | |
| Fig. 3: | Viable leaf area of spring wheat as affected by various temperatures and PEG800 with time |
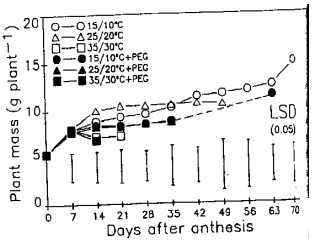 | |
| Fig. 4: | Plant mass of spring wheat as affected by various temperatures and PEG 800 with time |
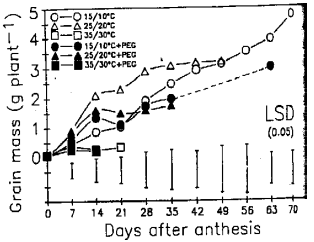 | |
| Fig. 5: | Grain mass of spring wheat as affected by various temperatures and PEG800 |
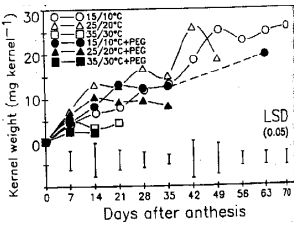 | |
| Fig. 6: | Kernel weight of spring wheat as affected by various temperatures and PEG800 with time |
Direct effects of high temperature on photosystem-11 and acceleration of senescence processes were undoubtedly more important (Harding et al.,1990a; Sayed et al., 1989a). When high temperature was combined with osmotic stress, however, stomatal conductivity may well have limited photosynthesis and other plant processes.
The interaction between temperature and osmotic stress is undoubtedly complex, but it may center around the response of stomatal conductivity (Hsiao, 1973). Low temperature enabled plants that were osmotically stressed to maintain nearly normal water contents thus, stomatal conductivity stayed high and photosynthetic rate and leaf area decreased only slightly as plants matured. The grain filling rate remained steady because translocation of assimilates is relatively insensitive to osmotic stress and continues even after photosynthesis is inhibited (Johnson and Moss, 1976).
High temperature induced the interplay between osmotic stress and stomata' resistance that is typically observed (Frank et al., 1973; Schonfeld et al., 1988; Shimshi et al., 1982). Increasing the temperature to moderate levels in the absence of osmotic stress affected stomata minutely until plants approached maturity and probably changed conductivity little if any. Imposing osmotic stress at the intermediate temperature induced stomatal closure within 14 d, however, which decreased photosynthesis and probably diminished leaf cooling by transpiration (Gates, 1968; Tashiro and Wardlaw, 1990).
Leaf temperature is determined by an energy budget that includes latent heat loss by transpiration (Gates, 1968). The cooling effect of transpiration is directly proportional to the difference in water vapor pressure between the mesophyll and the free air surrounding the leaf and to the diffusion conductivity of vapor from inside to outside the leaf (Gusta and Chen, 1987). The combination of osmotic stress and the intermediate temperature affected both components, reducing water vapor pressure of tissue in parallel studies and stomatal conductivity in the present studies. Under these circumstances, leaf temperature may increase 5 to 15°C above air temperature and greatly disrupt cellular processes (Gates, 1968).
Inducing stomata! closure osmotically in light initiated degradative processes and suggested strongly that aperture controls senescence of detached leaves (Tashiro and Wardlaw, 1990). Control may be mediated by the action of stomatal closure on cyclic phosphorylation or on a growth regulator to induce senescence, which was measured by change in leaf chlorophyll content. Exposing plants simultaneously to osmotic stress and moderately elevated temperature probably initiated a similar sequence that accelerated senescence processes more than either stress alone.
Responses of wheat to temperature and ost-hotic stress may have important agronomic applications for cultivating the crop in different environments. The relationships explain why wheat can be successfully cultivated even in areas of moderate rainfall by careful selection of seeding date so plants mature when the temperature is low. Conversely, the relationships explicate low yield in humid, wet tropics, where the temperature remains high throughout the growing season. Low water to irrigation in major production regions that are hot and arid also can be attributed to the effect of high temperature. Adverse effects of high temperature, which prevent plants from using moisture efficiently for growth, limit responses to supplemental water and reduce the yield that can be obtained.
REFERENCES
- Al-Khatib, K. and G.M. Paulsen, 1989. Enhancement of thermal injury to photosynthesis in wheat plants and thylakoids by high light intensity. Plant Physiol., 90: 1041-1048.
CrossRefDirect Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Fischer, R.A. and R. Maurer, 1978. Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust. J. Agric. Res., 29: 897-912.
CrossRefDirect Link - Frank, A.B., J.F. Power and W.O. Willis, 1973. Effect of temperature and plant water stress on photosynthesis diffusion resistance and leaf water potential in spring wheat. Agron. J., 65: 777-780.
Direct Link - Gates, D.M., 1968. Transpiration and leaf temperature. Ann. Rev. Plant Physiol., 19: 211-238.
CrossRefDirect Link - Harding, S.A., J.A. Guikema and G.M. Paulsen, 1990. Photosynthetic decline from high temperature stress during maturation of wheat II. Interaction with source and sink processes. Plant Physiol., 92: 654-658.
CrossRefDirect Link - Harding, S.A., J.A. Guikema and G.M. Paulsen, 1990. Photosynthetic decline from high temperature stress during maturation of wheat: I. Interaction with senescence processes. Plant Physiol., 92: 648-653.
PubMedDirect Link - Hsiao, T.C., 1973. Plant response to water stress. Ann. Rev. Plant Physiol., 24: 519-570.
CrossRefDirect Link - Lu, D.B., R.G. Sears, G.M. Paulsen, 1989. Increasing stress resistance by in vitro selection for abscisic acid insensitivity in wheat. Crop Sci., 29: 939-943.
Direct Link - Nicolas, M.E., R.M. Gleadow and M.J. Dalling, 1984. Effects of drought and high temperature on grain growth in wheat. Aust. J. Plant Physiol., 11: 553-566.
Direct Link - Planchon, C., 1979. Photosynthesis, transpiration, resistance to CO2 transfer and water efficiency of flag leaf of bread wheat, durum wheat and triticale. Euphytica, 28: 403-408.
CrossRefDirect Link - Sayed, O.H., M.J. Earnshaw and M.J. Emes, 1989. Photosynthetic responses of different varieties of wheat to high temperature II. Effect of heat stress on photosynthetic electron transport. J. Exp. Bot., 40: 633-638.
CrossRefDirect Link - Schonfeld, M.A., R.C. Johnson, B.F. Carver and D.W. Mornhinweg, 1988. Water relations in winter wheat as drought resistance indicators. Crop Sci., 28: 526-531.
CrossRefDirect Link - Shimshi, D., M.L. Mayoral and D. Atsmon, 1982. Responses to water stress in wheat and related wild species. Crop Sci., 22: 123-128.
CrossRefDirect Link - Sofield, I., L.T. Evans, M.G. Cook and I.F. Wardlaw, 1977. Factors influencing the rate and duration of grain filling in wheat. Aust. J. Plant Physiol., 4: 785-797.
CrossRefDirect Link - Tashiro, T. and I.F. Wardlaw, 1990. The effect of high temperature at different stages of ripening on grain set, grain weight and grain dimensions in the semi-dwarf wheat banks. Ann. Bot., 65: 51-56.
Direct Link - Wardlaw, I.F., 1967. The effect of water stress on translocation in relation to photosynthesis and growth I. Effect during grain development in wheat. Aust. J. Biol. Sci., 20: 25-40.
Direct Link