
M.M.A. El-Sawah
Microbiology Department, Faculty of Agriculture, Mansoura University, Mansoura, Egypt
Eman H. Ashour
Microbiology Department, Faculty of Agriculture, Mansoura University, Mansoura, Egypt
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1240-1244







ABSTRACT
Lactase was produced by Streptococcus salivarius subsp. thermophilus grown on deproteinized whey. Maximum lactase production in 6.85 percent whey supplemented with certain nutrients was 54.5 U/ml. Lactase was purified by ammonium sulfate and acetone fractionation and gel filtration. It was immobilized on chitin and chitosan by covalent binding. Lactase immobilized on chitosan had the highest activity for hydrolysis of whey lactose. The maximum amount of glucose produced by the immobilized lactase was 20 percent of lactose during conversion of 71 percent lactose in 6 h. Xanthomonas campestris was able to convert glucose, galactose, mixture of them, lactose, or whey to xanthan, where the yield was 86, 67, 83, 2, and 1.8 percent respectively, after 72 h in batch fermentation. When deproteinized whey, which was hydrolyzed by immobilized lactase, was used for xanthan production. The synthesis of icanthan was generally as good as with comparable conventional media, the yield reached to 65 percent.
PDF Abstract XML References Citation
How to cite this article
M.M.A. El-Sawah and Eman H. Ashour, 1999. Xanthan production from whey treated with immobilized lactase. Pakistan Journal of Biological Sciences, 2: 1240-1244.
DOI: 10.3923/pjbs.1999.1240.1244
URL: https://scialert.net/abstract/?doi=pjbs.1999.1240.1244
DOI: 10.3923/pjbs.1999.1240.1244
URL: https://scialert.net/abstract/?doi=pjbs.1999.1240.1244
INTRODUCTION
In recent years there is a great deal of interest in using industrial wasts as nutrient sources in bioconversions. Whey is a by-product of dairy industry. In Egypt, about 500,000 tons of whey are produced per year, most of this amount is run to drain without use constituting serious pollution problem (Facia, 1983). Whey is a fluid containing very low quantities of milk solids and high concentration of lactose. A typical sweet dairy whey contains 7 percent solids and is composed of about 70 percent lactose, 12 percent protein and smaller amount of organic acids, minerals and vitamins (Glass and Hedrick, 1977). In respect that its containing nutritionally valuable substances, whey is a good candidate to be utilize as an economic substrate by certain microorganisms for production of essential and beneficial compounds such as ethanol, lactic acid, amino acids and SCP (Mann, 1986).
Lactase could be obtained from several microorganisms including lactose fermenting molds, yeasts and bacteria. Kluyveromyces fragilis (Mahoney et al., 1975) and Streptococcus thermophilus (Rao and Dutta, 1981) are the best sources of the enzyme. The later, which is widely used as a starter organism for yogurt manufacture, is a food-safe organism and promising source of lactase. Recently, there is an effort to produce this enzyme in a large amount because of its application for hydrolyzing lactose in milk products to elevate problems associated with whey disposal.
Xanthomonas campestris is industrial interesting for its ability to produce xanthan gum, an extracellular high molecular weight polysaccharide, which is used in a variety of applications as a stabilizing, viscosifying, emulsifying, thickening and suspending agent (Becker et al., 1998). Many researchers have studied the fermentation conditions required for optimal gum production (Moraine and Rogovin, 1973; Kennedy, 1982; Torrestiana et al., 1990; Garca-Ochoa et al., 1992; De Vuyst and Vermeire, 1994; Rajeshwari et al., 1995). In industrial production of xanthan gum, whey may offer a cheap cultivation medium. Due to the low level of lactase present in X. campestris (Frank and Somkuti, 1979; Walsh, 1984 and Fu and Tseng, 1990), the bacterium is not able to grow well and produce xanthan gum in lactose medium. Xanthan gum can be successfully produced in whey-containing medium, if qualities of X. campestris are genetically engineered or if lactose is first hydrolyzed to glucose and galactose to utilize whey more efficiently.
The goal of this article away from genetic manipulation is the later approach which is more economical to achieve this production by cultivation the original strain (X. campestris) in whey pretreated with immobilized lactase.
MATERIALS AND METHODS
Bacterial strains: Streptococcus salivarius subsp. thermophilus EMCC 10509 was obtained from the microbial culture collection center of Cairo MIRCEN, Fac. Agric., Ain Shams Univ., Egypt and maintained on YPG medium which containing (g/L); yeast extract, 5.0; peptone, 10.0; glucose, 5.0; NaCI, 5.0 and agar 20.0. The pH was adjusted to 7.2-7.4. Xanthomonas campestris NRRL B-1459 was used throughout this study. The strain was maintained on slants with YPGM medium (Rajeshwani et al., 1995).
Whey Preparation: Whey from bovine was supplied as sweet dried whey from Sigma Co., USA. It contained (w/w) 73 percent lactose, 10.8 percent protein and 0.75 percent phosphorus. Deproteinized whey solutions were prepared by acidification (pH 4.5) and heat treatment (90°C for 30 min). Then, the preparation was cooled and filtered to remove the precipitated protein.
Lactase Production: Lactase was produced in optimum conditions as described by Menshawy (1997). The medium employed for lactase production contained 5.48 percent sweet dried whey (Sigma Co.) in water together with "supplementary nutrients" [0.5 % of NaH2PO4; 0.5 percent of yeast extract, and 0.3 percent of (NH4)2SO41. The initial pH was adjusted to 7.0. The strain was grown in shake flasks (120 rev/min) for 24 h at 40°C.
Extraction and Purification of Lactase: Harvested cells were washed twice with 0.067 M potassium phosphate buffer, pH 6.8. Washed cells were suspended in 25 ml of 0.067 M potassium phosphate buffer, pH 6.8, containing 0.5 mM MgSO4 and 0.1 mM MnCl2. The suspension was treated with toluene (2 percent v/v) at 37°C for 10 min. Broken cells were removed and discarded by centrifugation at 15 000 rpm for 30 min at 4°C and the supernatant was used as an enzyme source. Cell free extract (1000 ml) was collected and precipitated by the addition of one volume of acetone at 4°C. Then, the precipitate was collected by centrifugation and the residual acetone was evaporated using an air stream. The precipitate was dissolved in phosphate buffer at pH 7.0 and dialyzed at 4°C against the same buffer. Solid (NH4)2SO4 up to 60 percent saturation was added slowly to the dialyzed solution, the precipitate which formed was collected by centrifugation at 15000 rpm for 20 min in refrigerated centrifuge, resuspended in phosphate buffer, pH 7.0 and dialyzed extensively against the same buffer at 4°C. The partially purified enzyme was applied on a Sephadex G-100 column to remove the (NH4)2SO4, then active fraction was collected and used as a source of lactase.
Preparation of Immobilized Lactase: Chitosan beads were prepared by shaking 0.5 g chitosan in 10 ml of 0.1 M HCI containing 2.5 percent glutaraldehyde for 2 h at 30°C. The beads were precipitated by the addition of 0.1 M NaOH to neutrality. The beads were collected by filtration and washed with water. The wet chitosan beads were mixed with 5.0 ml of the lactase solution (1000 units). After being shaken for 1 h at 30°C, the unbound lactase was removed by washing with distilled water. In case of chitin, 0.5 g chitin was shaked in 10 ml of 2.5 percent glutaraldehyde in 0.1 M acetate buffer (pH 5.5) for 2 h. The solid material was filtered and washed. The treated chitin was mixed with 5.0 ml of the lactase solution (1000 units) for 1 h at 30°C. The unbounded lactase was removed by washing with distilled water (Ohtakara, 1988).
Assay of Immobilized Lactase Activity: Lactase (approximately g of protein) were suspended in 50 mmol/l of phosphate buffer (pH 6.9) containing 4.56 percent whey lactose at 40°C with magnetic stirring. After 10 min of incubation or at various times a portion of the reaction mixture was removed and the amount of lactose hydrolyzed was assayed. One unit of immobilized lactase was defined as the amount of immobilized lactase required to hydrolyze 1 μmmol of lactose per min at 40°C and pH 7.0.
Whey Hydrolysis with Immobilized lactase: Whey permeate, adjusted to pH 7.0, was hydrolyzed using immobilized lactase. Hydrolyzed whey permeate was supplemented with a basal medium (per liter K2HPO4, 7 g; KH2PO4, 2 g; NH4NO3, 0.6 g; FeSO4.7H20, 0.01 g; MgC12.6H20, 0.1 g and MnC12, 0.001 g; Na-citrate, 2 g and 0.125 percent tryptone). The pH of the solution was finally adjusted to 7.0 with 2N H2SO4 and 2N KOH. 50 ml of whey medium was dispensed into 250 ml Erlenmeyer flasks, sterilized, then, 5 ml of 24 h old inoculum was added (for preparing inocula, a fresh culture grown on the YPGM agar slant at 28°C for 48 h was transferred to 500 ml Erlenmeyer flasks containing 100 ml of the YPGM broth and incubated at 28°C and 250 rec/min for 24 h).
Xanthan Production: In shake-flask (250 rev/min on a rotary shaker) experiments, cultures were grown at 30°C using 250-ml Erlenmeyer flasks containing 50 ml of liquid xanthan production medium (a basal medium amended with different sugar sources) and inoculated by 10 percent (v/v) inoculum. Inocula was prepared as previously mentioned.
Sugar determination: Glucose was determined by glucose oxidase peroxidase. Galactose was determined by galactose oxidase peroxidase. Lactose was determined by Nickerson et al. (1976) method.
Xanthan Determination: Cultures were centrifuged at 10,000 xg for 10 min. To aliquots of the supernatant ethanol was added to 70 percent concentration with agitation followed by standing in cold for 3 hr. The precipitates were collected by centrifugation and redissolved in aliquots of distilled water. Modified Anthrone method (Trevelyan and Harrison, 1952) was used for quantitative determination. The standard curve was constructed by using pure xanthan.
Viscosity Measurement: Viscosity was measured using a rotational viscometer at a constant shear rate of 10.71S–1 and 30°C.
pH Determination: The pH value was measured using laboratory pH-meter with glass electrodes (Knick-Digital-pH meter 646).
Protein determination: Protein was determined by the method of Lowry et al. (1951), using crystalline bovine serum albumin as standard.
Lactase activity determination: The activity of lactase was determined at 35°C using 45 mM lactose as substrate (in 50 mM phosphate buffer, pH 7.0). Glucose evolution was analyzed using the glucose oxidase/peroxidase system (Bergmeyer and Bernt, 1974). One unit of lactase activity is defined as the amount of enzyme that hydrolyses 1 mmmol lactose/min at 35°C.
RESULTS AND DISCUSSION
Lactase production: Lactase production by S. salivarius subsp. thermophilus, grown in optimum conditions in different concentrations of whey supplemented with essential nutrients, was investigated. Data represented in Fig. 1 show that after 24 h of cultivation, total lactase activity (54.5 μ/ml) was obtained with 6.85 percent whey (5.0% lactose). Consumption of whey lactose reached 93.44 percent after 24h. The enzyme level and remaining lactose, in whey medium, is within the range of values reported previously for S. salivarius subsp. thermophilus (Rao and Dutta, 1981; Menshawy, 1997).
Purification of lactase: Purification parameters are summarized in Table 1. A 29.25-fold purification of the lactase was achieved with an overall recovery of 59.29 percent.
| Table 1: | Summary of the partial purification of lactase from S. salivarius subsp. thermophilus |
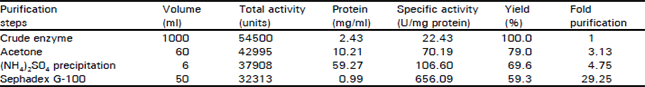 | |
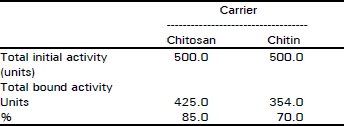
| Table 2: | Immobilization of lactase by covalent binding on chitosan and chitin |
 | |
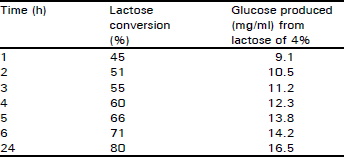
| Table 3: | Lactose hydrolysis by lactase at 37°C. Experimental conditions: 5.48% deproteinized whey (4% lactose), pH 7.0, 15 U lactase/ml |
 | |
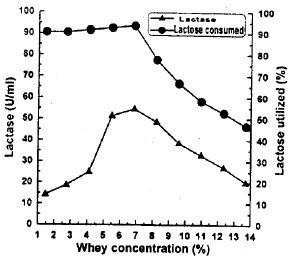 | |
| Fig. 1: | Lactase production and lactose utilization during growth in different concentrations of whey. |
The results are in line with Rao and Dutta (1981).
Immobilization of lactase: The immobilization of S. salivarius subsp. thermophilus lactase by covalent binding though glutaraldehyde onto chitin and chitosan was carried out (Table 2). A considerably good loading efficiency (354 units/g) for lactase immobilized on chitin, but low immobilization yield was detected (70 percent). Lactase immobilized on chitosan by covalent binding showed higher activity (425 units/g) and better immobilization yield (85%). Therefore, chitosan was used for lactase immobilization for the purpose of whey hydrolysis. Immobilization of lactase on chitosan might be useful in processing milk by-products into valuable sweeteners, or into nutritional media for fermentation processes. Its possible application to solving many problems related to the use of lactose should also be considered (Leuba and Widmer, 1977; Greenberg and Mahoney, 1981).
Whey lactose hydrolysis: Quite high activity for the hydrolysis of lactose was obtained by immobilized lactase. Table 3 is indicated the concentrations of glucose produced during hydrolysis of 5.48 percent whey (4.0 percent lactose) with the lactase immobilized onto chitosan. A maximum of 20 percent glucose of the total lactose was obtained by 71 percent conversion of lactose, which is slightly less than with the free lactase. The results are in line with Huffman and Harper (1985) and Mozaffar et al. (1986).
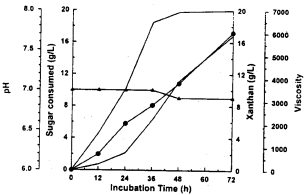 | |
| Fig. 2a: | X.campestris behavior in xanthan production medium amended with glucose. Sugar consumed (∇), Xanthan (●), viscosity (○), pH (▴) |
Xanthan production: The basal medium was supplied with glucose, galactose, or both of them (mixture of glucose and galactose 50/50) and used as xanthan production medium by X. campestris. The results given in Fig. (2a-c) revealed that glucose and galactose either alone or in combination were utilized for xanthan synthesis. Glucose was consumed more rapidly than galactose. The results also showed simultaneous utilization of both sugars. After 72 h of batch fermentation, 17.22 (g/L) of xanthan accumulated in the broth. This maximum yield recorded 86 percent after 72 h by using glucose medium.
When lactose used in the production medium as a sole carbon source, slight amount of lactose was assimilated by the tested organism. Only 0.4 g of xanthan accumulated in the liter broth after 72 h of batch fermentation (Fig. 2d).
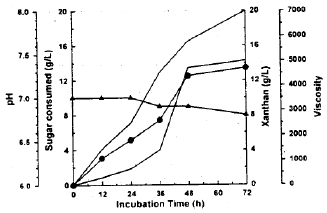 | |
| Fig. 2b: | X. campestris behavior in xanthan production medium amended with galactos. Sugar consumed (∇), Xanthan (●), viscosity (○), pH (▲) |
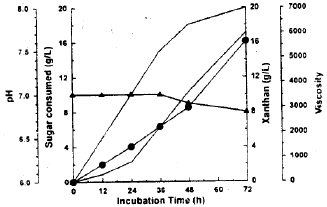 | |
| Fig. 2c: | X. campestris behavior in xanthan production medium amended with glucose+galactose. Sugar consumed (∇), Xanthan (●), viscosity (○), pH (▲) |
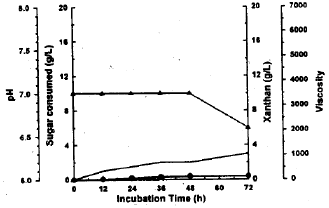 | |
| Fig. 2d: | X. campestris behavior in xanthan production medium amended with lactose. Sugar consumed (∇), Xanthan (●) viscosity (○), pH (▲) |
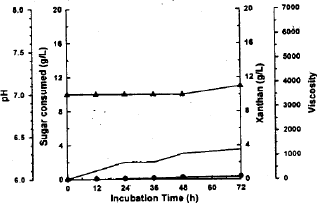 | |
| Fig. 2e: | X. campestris behavior in xanthan production medium amended with whey. Sugar consumed (∇), Xanthan (●), viscosity (○), pH (▲) |
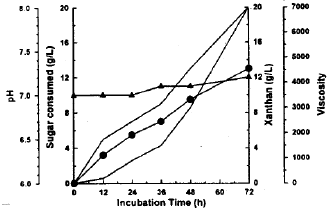 | |
| Fig. 2f: | X. campestris behavior in xanthan production medium amended with pretreated whey. Sugar consumed (∇), Xanthan (●), viscosity (○), pH (▲) |
The low level of lactase present in X. campestris may explain this behavior. It was expected according to Stauffer and Leeder (1978) who found that no significant amount of xanthan gum is produced when X. campestris is grown in lactose medium. Frank and Somkuti (1979), Walsh (1984) and Fu and Tseng (1990) reported that X. tampestris was not able to grow in a lactose medium and to synthesize high xanthan yield because of the low concentration of produced lactase. Moreover, Drahovska and Turna (1995) found that original strain X. campestris 1069 produced 10.5 g of xanthan gum in liter of glucose medium after 16 h and no detectable amount of xanthan in lactose medium.
Slight amounts of lactose from whey were assimilated by X. campestris in whey medium, the yield percent was very low (1.75 percent) and only 0.35 g xanthan /L after 72 h (Fig. 2e). This result in agreement of the previous data about xanthan production in lactose medium, therefore, an attempt to use lactose-based substrate (such as whey) for gum production by X. campestris would be difficult. Some attempts have been previously made by several groups to construct lactose utilizing X. campestris strains for xanthan production. Fu and Tseng (1990) were able to select a strain which can utilize lactose for xanthan production, but the strain was not stable.
Therefore in this study, a mobilizable lactase was introduced into whey firstly to hydrolyse lactose to fermentable sugar, glucose and galactose. In that case, the bacterium was able to grow and successfully produce xanthan from pretreated whey in amounts near to those produced by galactose-grown cells, reached to 12.99 g xanthan/L after 72 h (Fig. 2f) and was 40-fold higher than in a medium containing whey. The results obtained are an indication for the possibility usage of pretreated whey by immobilized lactase in the process of industrial production of xanthan.
REFERENCES
- Becker, A., F. Katzen, A. Puhler and L. Ielpi, 1998. Xanthan gum biosynthesis and application: A biochemical/genetic perspective. Applied Microbiol. Biotechnol., 50: 145-152.
CrossRefDirect Link - De Vuyst, L. and A. Vermeire, 1994. Use of industrial medium components for xanthan production by Xanthomonas campestris NRRL-B-1459. Applied Microbiol. Biotechnol., 42: 187-191.
CrossRefDirect Link - Drahovska, H. and J. Turna, 1995. Construction of lactose-utilizing Xanthomonas campestris with a mini-mu derivative. Applied Environ. Microbiol., 61: 811-812.
Direct Link - Frank, J.F. and G.A. Somkuti, 1979. General properties of beta-galactosidase of Xanthomonas campestris. Applied Environ. Microbiol., 38: 554-556.
Direct Link - Fu, J.F. and Y.H. Tseng, 1990. Construction of lactose-utilizing Xanthomonas campestris and production of xanthan gum from whey. Applied Environ. Microbiol., 56: 919-923.
PubMedDirect Link - Garca-Ochoa, F., V.E. Santos and A.P. Fritsch, 1992. Nutritional study of Xanthomonas campestris in xanthan gum production by factorial design of experiments. Enzyme Microbiol. Technol., 14: 991-996.
CrossRefDirect Link - Glass, L. and T.I. Hedrick, 1977. Nutritional composition of sweet-and acid-type dry wheys. II. Vitamin, mineral and calorie contents. J. Dairy Sci., 60: 190-196.
CrossRefDirect Link - Kennedy, J.F., P. Jones, S.A. Barker and G.T. Banks, 1982. Factors affecting microbial growth and polysaccharide production during the fermentation of Xanthomonas campestris cultures. Enzyme Microb. Technol., 4: 39-43.
CrossRefDirect Link - Leuba, J.L. and F. Widmer, 1977. Immobilization of the β-galactosidase from Aspergillus niger on chitosan. Applied Biochem. Biotechnol., 2: 257-271.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mahoney, R.R., T.A. Nickerson and J.R. Whitaker, 1975. Selection of strain, growth conditions and extraction procedures for optimum production of lactase from Kluyveromyces fragilis. J. Dairy Sci., 58: 1620-1629.
CrossRefDirect Link - Moraine, R.A. and P. Rogovin, 1973. Kinetics of the xanthan fermentation. Biotechnol. Bioeng., 15: 225-237.
CrossRefDirect Link - Mozaffar, Z., K. Nakanishi and R. Matsuno, 1986. Continuous production of galacto-oligosaccharides from lactose using immobilized β-galactosidase from Bacillus circulans. Applied Microbiol. Biotechnol., 25: 224-228.
CrossRefDirect Link - Nickerson, T.A., I.F. Vujicic and A.Y. Lin, 1976. Colorimetric estimation of lactose and its hydrolytic products. J. Dairy Sci., 59: 386-390.
CrossRefDirect Link - Rajeshwari, K.V., G. Prakash and P. Ghosh, 1995. Improved process for xanthan production using modified media and intermittent feeding strategy. Lett. Applied Microbiol., 21: 173-175.
CrossRefDirect Link - Rao, M.R. and S.M. Dutta, 1981. Purification and Properties of Beta‐galactosidase from Streptococcus thermophilus. J. Food Sci., 46: 1419-1423.
CrossRefDirect Link - Stauffer, K.R. and J.G. Leeder, 1978. Extracellular microbial polysaccharide production by fermentation on whey or hydrolyzed whey. J. Food Sci., 43: 756-758.
CrossRef - Torrestiana, B., L. Fucikovsky and E. Galindo, 1990. Xanthan production by some Xanthomonas isolates. Lett. Applied Microbiol., 10: 81-83.
CrossRefDirect Link - Trevelyan, W.E. and J.S. Harrison, 1952. Studies on yeast metabolism. 1. Fractionation and microdetermination of cell carbohydrates. Biochem. J., 50: 298-303.
PubMedDirect Link - Walsh, P.M., M.J. Haas and G.A. Somkuti, 1984. Genetic construction of lactose-utilizing Xanthomonas campestris. Applied Environ. Microbiol., 47: 253-257.
PubMed