
F. Ahmet Ayaz
Department of Biology, Karadeniz Technical University, 61080 Trabzon, Turkey
Huseyin Inceer
Department of Biology, Karadeniz Technical University, 61080 Trabzon, Turkey
Sema Hayirlogliu-Ayaz
Department of Biology, Karadeniz Technical University, 61080 Trabzon, Turkey
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1139-1142







ABSTRACT
Soluble (non-reducing) and non-soluble (reduced) protein profiles of nine taxa of Vicia species are V. sepium L., V. peregrine L., V. hybrida L. and V. sativa L. subsp. nigra (L.) Ehrh, from Sect. Vicia, V. cassubica L. and V. cracca L. subsp. cracca from Sect. Cracca, V. tetrasperma (L.) Schreb. and V. hirsute (L.) S. F. Gray. from Sect. Ervum and V. bithynica L. from Sect. Faba were studied. . The non-reducing protein profile from the seeds exhibited a major set of proteins with molecular masses in the range of 66-34.7 kDa and the reduced protein profile showed major proteins with molecular masses in the range of 66-45 kDa. Spectrophotometric determinations of non-reduced protein amount was found significantly different (p =0.05) in the seeds of V. hirsute, V. sativa subsp, nigra and V. bithynica while the remaining species were not determined significantly different. As for reduced protein amount, the amount of reduced protein of V. hirsute was found significantly different among the species (p=0.05). The highest level of non-reduced and reduced proteins were determined for the seed of V. hirsute.
PDF Abstract XML References Citation
How to cite this article
F. Ahmet Ayaz, Huseyin Inceer and Sema Hayirlogliu-Ayaz, 1999. An Electrophoretic Analysis of the Seed Proteins of some Vicia L. Species from Northeast Anatolia (Turkey). Pakistan Journal of Biological Sciences, 2: 1139-1142.
DOI: 10.3923/pjbs.1999.1139.1142
URL: https://scialert.net/abstract/?doi=pjbs.1999.1139.1142
DOI: 10.3923/pjbs.1999.1139.1142
URL: https://scialert.net/abstract/?doi=pjbs.1999.1139.1142
INTRODUCTION
The systematic methodology mainly based on morphology has been improved by the incorporation of physiology, ecology or biochemical characters. Severel biochemical analysis, especielly of proteins, meke it possible to establish differences at various taxonomic levels (Vaughan, 1983). One of the biochemical methods more extensively used for taxonomic purposes has been the electrophoretic analysis of the proteins found in seed and storage orgaras. These proteins are physilogically stable and easly to handle (Ladizinsky and Hymowitz, 1979). Seed proteins have received extensive attention in literature (Derbyshire et al., 1976; O'Kenedy et al., 1979; Lalonde et al., 1984; Aliaga-Morell et al., 1987; Gifford, 1988; Collada et al., 1988). Considerable insight has been dained as to their structure and synthesis during seed development and to their role as storage proteins following seed inhibition (Higgins, 1984). Seed proteins have previously been classified according to their exraction by a series of solvents (Osborne, 1924; Shewry and Miflin, 1985). The storage proteins in the majority of plant species studied are of the globin type. Two classes have been described: 11 S or legumin type' globulins and 7 S or vicilin type' (Derbyshire et al., 1976; Casey et al., 1986). Legumins, i.e. 11/12 S storage proteins, are the major seed proteins the legumens Vicia faba L. (Bailey and Boulter, 1970), Pisum sativum L. and in other Vicia species (Derbyshire et al., 1976). Similar proteins with the significant legumin characters (acidic subunits of Mr =c. 30-40 KD, basic subunits of Mr = c. 20 KD, linked together by S-S bridges and forming hexametric molecules in the storage tisues) have also been recognized as major storage proteins in many dicot and monocot species (De Klerk and Engelen, 1985; Fischer and Schopfer, 1988; Hasegawa et al., 1978; Jensen, 1984; Jensen and Buttner, 1981; Konishi et al., 1985; Simon et al., 1985). In this investigation we analysed the occurence of the soluble and insoluble proteins in some Vicia species collected from Northeast Anatolia (Turkey) were surveyed. No studies have previously been reported on electrophorotic separation of the storage proteins of given 9 Vicia taxa from Turkey.
MATERIALS AND METHODS
Seed materials: The seeds used in this study for protein extraction were collected from Northeast Anatolia (Turkey) are V. sepium L., V. peregrine L., V. hybrida L. and V. sativa L. subsp. nigra (L.) Ehrh, from Sect, Vicia, V. cassubica L. and V. cracca L. subsp. cracca from Sect. Cracca, V. tetrasperma (L.) Schreb. and V. hirsute (L.) S. F. Gray. from Sect. Ervum and V. bithynica L. from Sect. Faba.
Protein extraction: A modification of Gifford (1988) method (Gifford et al., 1982) was used for the protein exraction from the seeds. Seed coats were removed prior exraction. Endosperm plus embryo were ground in Tris-glycine-buffer (0.01 M Tris; 0.08 M glycine), pH (.2+2 percent NaCI (Jensen and Berthold, 1989) in a ratio of 1 g seed material to 10 ml buffer. After 30 min stirring the slurry was centrifugated at 16.000 g for 20 min at 10°C. The supernatants, containing the soluble proteins, were removed. The pelled was thoroughly reexracted twice with a duble volume of buffer to exract all soluble proteins. The pelled containing the reduced storage proteins was suspented in TGP buffer (0.01 M Tris, 0.08 M glycine) pH 8.2+2 percent NaCI and an equal volume of 62 mM Tris-HCl (pH 6.8) buffer with 3.05 percent (w/v) SDS and 10.7 percent (w/v) glycerol. The suspension was boiled for 8 min. After centrifugating the supernatant was applied for SDS-PAGE.
Electrophoresis: Proteins in the supernatant were quantified by the medhot of Laemmli (1970) and electrophoretically separated according to Jensen and Lixue (1991) in SDS-PAGE. Fifty microgram of protein were applied to each slot and at 4°C under a constant of intensity of a mA per slot for 3.5 h. Reaching a final voltage of 300 V. Gels were stained with silver nitrate (Sambrook and Russell,1989). The standard proteins (MW-SDS-702, Sigma Chemical Co.) were employed are trypsin inhibitor (20.1 kDa) trypsinogen (24 kDa), carbonik anhydrase (29 kDa), glyceraldhyde-3-phosphate dehydrogenase (34,7 kDa), egg albumin (45 kDa) and bovine albumin (66 kDa).
RESULTS AND DISCUSSION
Seeds of some Vicia L. species were extracted for SDS-soluble seed proteins and their electrophoretic behaviour was determined by SOS-PAGE. The saver stained gray protein profiles obtained after separating these proteins in their reduced (non-soluble) (+ME) and nonreduced (soluble) (-ME) states are shown in Fig. 1. The total soluble and non-soluble protein amount of the seeds were given in Table 1. The non-reducing gel (Fig. 1a) shows that the soluble protein profile from the seeds are different and has a major set of proteins with molecular masses in the range of 66-34.7 kDa. Whereas the nonsoluble protein profile has major proteins with molecular masses in the range of 66-45 kDa (Fig. 1b).
Table 2 shows presence or absence of non-reducing and reducing seed proteins in Vicia species separated by SDS-PAGE. There were dramatic changes in the protein composition of the seeds were classified according to extraction procedures (see Materials and Methods). The non-reduced protein amount was found significantly different (p = 0.05) in the seeds of V. hirsute, V. sativa and V. bithynice while the remaining species were not determined significantly different. As for reduced protein amount, the amount of reduced protein of V. hirsute was found significantly different among the species (p = 0.05k The highest level of non-reduced and reduced protein were determined for the seed of V. hirsute.
When the proteins from seeds of Vicia of nine taxa was electrophoresed, the non reducing protein profiles (Fig. 1a) of the various species had a common protein band with an estimated molecular weight of 34.7 kDa and 24 kDa. The reducing protein profiles (Fig. 1b) of the various species had a common protein band with an estimated molecular weight of 34.7 kDa . The 66 kDa protein band as in non reducing protein profiles were identified for the species V. cracca subsp. cracca, V. hybrida, V. sativa subsp. ngra, V. bithynica and V. peregrine. The non reducing protein profiles of some species except V. sepium, V. tetresperrna, V. hirsute and V. hybrida and V. sepium, V. tetrasperma and V. peregrine were found to be specific for 45 kDa and for 29 kDa protein bands, respectively while V cassubica, V. bithynica and V. peregnine for 20 kDa and V. cracca subsp. cracca, V. bithynice and V. peregrine for 14 kDa protein bands were found specific (Table 2).
The reducing protein profiles exhibited most of the protein bands at 34.7 kDa in all Vicia species except that V. sepium and V. hirsute, The 66 kDa protein band as in reducing protein profiles was determined in V. sepium, 45 kDa in V. tetrasperma, V. bithynice and V. peregrine, 29 kDa in V. sepium, V. tetrasperma and V. bithynica and 20 kDa in V. terrasperma and V. sativa subsp. nigra (Fig. 1b). In many seeds, storage proteins are localized within an amorphous matrix contained in single membrane bound organelles called protein bodies (Lott, 1980), Commonly, these proteins are globulins (Derbyshire et al., 1976). However, in some seeds inclusions of crystalloid storage ,proteins are also found embeded in the matrix.
| Table 1: | Protein content of non-reducing and reducing fractions of Vicia L. seeds. Results are the means of three replicates of 5 seeds of each. For comparisons among means the analysis of variance was used |
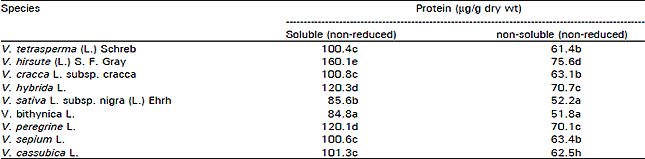 | |
| Means in columns followed by different letters are different at the p = 0.05 level | |
| Table 2: | Presence (+) or absence (-) of non-reducing and reducing seed proteins in some Vicia L. species collected from Northeast Anatolia (Turkey) |
 | |
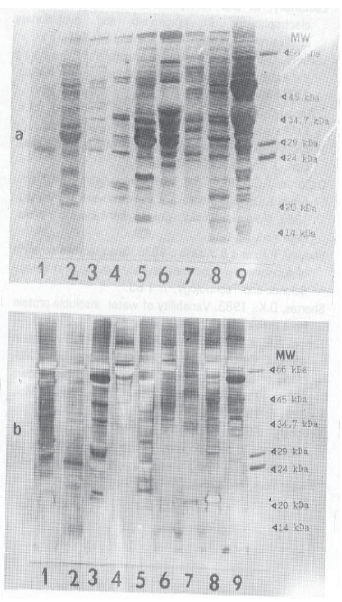 | |
| Fig. 1: | SDS-PAGE protein profiles from seeds of Vicia L.; a) non-reducing protein profile (Tris-glycine-buffer extracts + 2 percent NaCI), b) reducing protein profiles (Tis-glycine-buffer extracts + 2 percent NaCI and Tris-SDS-glycerol). 1; V. sepium L., 2; V. cassubica L., 3; V. tetrasperma (L.) Schreb, 4; V. hirsuta (L.) S. F. Gray, 5; V. cracca L. subsp. cracca., 6; V. hybrida L., 7; V. sativa L. subsp. nigra (L.) Ehrh., 8; V. bithynica L. 9; V. peregrina L. (45, 34.7, 20 and 14 kDa molecular weights in standard couldn't clearly be seen because of the low density in staining) |
Ultrastructural studies have shown that protein bodies localized in the storage parenchyma of seeds of several Pinus species contain crystalloid protein inclusions.
Although legumins have been reported to be the predominant storage protein in seed plants, exceptions are known (Danielsson, 1949). In gymnosperms, legumins has also been detected in the seeds of Ginkgo biloba (Jensen and Berthold, 1989), but not in Macrozamia commur (Blagrove et al., 1984). In Abies species, all major legumin proteins were characterised at 55 kDa (Jensen and Lixue, 1991). Most of the species of Poaceae family store predominantly prolamine. The oat accumulates 11S globulins and rice store glutelins (Kasarda et al., 1976; Shewry and Miflin, 1985). In some species of Leguminasae, such as Vicia faba L. and Pisum sativum L., the 11 and 12S proteins were described as the major seed proteins (Derbyshire et al., 1976). 30-40 and 20 kDa proteins were recognized in many dicot and monocot species (De Klerk and Engelen, 1985; Fischer and Schopfer, 1988; Hasegawa et al., 1978; Jensen, 1984; Jensen and Buttner, 1981; Konishi et al., 1985; Simon et al., 1985). Similarly, we determined most of the soluble proteins between the ranges at 30-40 and 20 kDa. This paper does not include the classification of protein types in the given species, but more detailed analyses will be required to detect protein types and classes bay increasing the number of species in the genus upon this work.
REFERENCES
- Aliaga-Morell, J.R., F.A. Culianez-Macia, G. Clemente-Marin and G. Primo-Yufera, 1987. Differentiation of rice varieties by electrophoresis of embryo protein. Theor. Applied Genet., 74: 224-232.
CrossRefDirect Link - Bailey, C.J. and D. Boulter, 1970. The structure of legumin, a storage protein of broad bean (Vicia faba) seed. Eur. J. Biochem., 17: 460-466.
CrossRefDirect Link - Blagrove, R.J., G.G. Lilley and T.J.V. Higgins, 1984. Macrozin, a novel storage globulin from seeds of Macrozamia communis L. Johnson. Funct. Plant Biol., 11: 69-77.
CrossRefDirect Link - Collada, C., R.G. Caballero, R. Casado and C. Aragoncillo, 1988. Different types of major storage seed proteins in Fagaceae species. J. Exp. Bot., 39: 1751-1758.
CrossRefDirect Link - Danielsson, C.E., 1949. Seed globulins of the Gramineae and Leguminosae. Biochem. J., 44: 387-400.
Direct Link - Derbyshire, E., D.J. Wright and D. Boulter, 1976. Legumin and vicilin, storage proteins of legume seeds. Phytochemistry, 15: 3-24.
CrossRefDirect Link - Fischer, W. and P. Schopfer, 1988. Isolation and characterization of mustard (Sinapis alba L.) seed storage proteins. Plant Biol., 101: 48-56.
CrossRefDirect Link - Gifford, D.J., J.S. Greenwood and J.D. Bewley, 1982. Deposition of matrix and crystalloid storage proteins during protein body development in the endosperm of Ricinus communis L. cv. Hale seeds. Plant Physiol., 69: 1471-1478.
Direct Link - Gifford, D.J., 1988. An electrophoretic analysis of the seed proteins from Pinus monticola and eight other species of pine. Can. J. Bot., 66: 1808-1812.
Direct Link - Higgins, T.J.V., 1984. Synthesis and regulation of major proteins in seeds. Annu. Rev. Plant Physiol., 35: 191-221.
Direct Link - Jensen, U. and C. Lixue, 1991. Abies seed protein profile divergent from other Pinaceae. Taxon, 40: 435-440.
CrossRefDirect Link - Jensen, U. and H. Berthold, 1989. Legumin-like proteins in gymnosperms. Phytochemistry, 28: 1389-1394.
CrossRefDirect Link - Konishi, Y., Y. Fumita, K. Ikeda and C. Okuno, 1985. Isolation and characterization of Globulin from seeds of Amaranthus hypochondriacus L. Agric. Biol. Chem., 49: 1453-1459.
CrossRefDirect Link - Lalonde, L., D.W. Fountain, A. Kermode, F.B. Ouellette, K. Scott, J.D. Bewley and D.J. Gifford, 1984. A comparative study of the insoluble storage proteins and the lectins of seeds of the Euphorbiaceae. Can. J. Bot., 62: 1671-1677.
Direct Link - Ladizinsky, G. and T. Hymowitz, 1979. Seed protein electrophoresis in taxonomic and evolutionary studies. Theor. Applied Genet., 54: 145-151.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Shewry, P.R. and B.J. Miflin, 1985. Seed storage proteins of economically important cereals. Adv. Cereal Sci. Technol., 7: 1-83.
Direct Link - Simon, A.E., K.M. Tenbarge, S.R. Scofield, R.R. Finkelstein and M.L. Crouch, 1985. Nucleotide sequence of a cDNA clone of Brassica napus 12S storage protein shows homology with legumin from Pisum sativum. Plant Mol. Biol., 5: 191-201.
CrossRefDirect Link