
N. Altaf
Nuclear Institute for Agriculture and Biology, Jhang Road, Faisalabad
J. Iqbal
Nuclear Institute for Agriculture and Biology, Jhang Road, Faisalabad
Botany Department, Punjab University, Quaid-e-Azam Campus, Lahore-Pakistam
M. S. Ahmad
Nuclear Institute for Agriculture and Biology, Jhang Road, Faisalabad
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 4 | Page No.: 321-325







ABSTRACT
Culture responses of 9 lentil explants were compared in various media having different ingredients. The most promising were the scrapped axillary portions of basal nodes for enhanced regeneration. The optimum size of the explant was also determined. The callogenesis response varied with the explant and ranged between 0.3-0.5 cm.
Among the sugars tested, sucrose was the best for callussing. Auxin 2,4-D induced callussing but cytokinin enhanced formationof shoot primordia in explants. Both BA and K promoted multiple shoot formation but K and GA gave more shoot buds. Cytokinins with auxins gave callusing. Callus was also induced in explants when grown in K (10 mg L–1) + GA (1 mg L–1) containing medium in the dark. When callus was shifted to light it started regeneration as green dots. The addition of seed extract to K and GA medium considerably enhanced regenerations.
PDF Abstract XML References Citation
How to cite this article
N. Altaf, J. Iqbal and M. S. Ahmad, 1998. Influence of Explant Source and Medium Ingredients on
Culture Responses of Microsperma Lentil Cultivar Masoor-85. Pakistan Journal of Biological Sciences, 1: 321-325.
DOI: 10.3923/pjbs.1998.321.325
URL: https://scialert.net/abstract/?doi=pjbs.1998.321.325
DOI: 10.3923/pjbs.1998.321.325
URL: https://scialert.net/abstract/?doi=pjbs.1998.321.325
INTRODUCTION
Lentil is a cold season annual legume and second major pulse crop after chickpea in Pakistan and it is fifth most important pulse of the world (FAO, 1988), Pakistani lentil is microsperma and average yield of national lentil is 408 kg ha–1 which is very low as compared to that of the world average of 812 kg ha–1 (FAO, 1992). According to the (GOP, 1996), the production is almost static since 40 years except a dip in production in 1984 because of rains.
A number of factors are responsible for its low yield and high production cost. These include domestic market lentil prices, annual rainfall, and cultivation on marginal rainfed areas. Fifty percent of the cultivated area for lentil lies in Punjab and North West Frontier Province. Lack of soil moisture at the time of planting is the most serious problem. Another one lentil has weak stem which make it lodging susceptible in heavy rains. Another problem for lentil is the slow growing habit as compared to that of weeds. Weeds take about 30-40% of plant nutrients applied to the crop (Dryden and Krishnamurthy, 1977).
The specific objectives of present work is in vitro culturing of lentil tissues leading to recovery of regenerated shoots, rooting of these shoots, establishment of plants in soil and selection of somaclonal variants.
This paper describes the influence of explant source origin and some medium ingredients to have sufficient amount of shoot regenerating callus under different culture conditions.
MATERIALS AND METHODS
Lentil genotype Masoor-85 was used. For explant source, healthy seeds were sterilized either with 0.5% HgCl2 for 20 min or with NaOCI (7%) for 30 min. Subsequently seeds were washed with sterile distilled water. Seeds were allowed to germinate on MS medium (Murashige and Shoog, 1962) with 3% sucrose. contaminated seeds, Seeds germinated at 25°C±1°C within 3-4 days. All part of the germinating seedling i.e. shoot apex, leaf, stem, epicotyl, hypocotyl, cotyledonary node, cotyledonary node with cotyledons, root and seed were used as explant for in vitro culturing. The explants were cultured with basal cut end facing the agar medium.
Initally two media MS (Murashige and Skoog, 1962) and B5 (Gamborg et al., 1968) were tested for growing lentil explants. However, MS was found better as explants remained fresh, healthy and vigorous on this medium. The pH of medium was adjusted to 5.7 with KOH or HCl. Difco Bacto agar (1%) was used to solidity the medium. Medium was autoclaved at 20 psi for 10 min. The filter sterilized growth regulators (IAA, IBA, GA) were added after autoclaving the medium.
Growth room had fluorescent light intensity of 3000-4000 lux with light/dark cycle of 16/8 h. For optimum temperature selection for callussing, explants were grown at 18, 20, 22 and 25°C in medium containing K+GA in the dark. The optimum response was obtained at 20-22°C.
Comparative growth rate of different explants: To evaluate the comparative growth rate of various explants were grown in MS liquid medium containing K and 2,4-D (each 2 mg L–1) + Glutamine 5 mg L–1 and the increase in the weight of the explants was recorded after six week growth.
Study for the optimum size of the explants for callus culture: To find out an appropriate size of the explant for callussing, explants varying in size from 0.2 to 0.6 cm were cultured in callussing medium MS containing K + 2,4-D (each 2 mg L–1) for one month and the optimum size for callus formation for each explant was recorded.
Requirements for callussing, growth and shoot regenerations: Medium requirements for induction of callus and formation of shoot primordia in different explants both under light (16/8 h, 4000 lux) and complete dark conditions were determined.
Effect of IBA exposure to lower stem segments and morphogenetic responses: The plants of two month age (Masoor-85) were used grown under sterile conditions in jar. Three segments of stem within two inch distance above the cotyledons were cultured in MS medium containing IBA from 0 to 1.5 mg L–1. The cultures were kept in dark for five days and then transferred to MS basal medium of 1/10th strength and kept in 16/8 h light dark cycle. The growth responses of stem sections were recorded after one month growth.
Influence of different types of cut made on explant, for callussing and organogenesis: All explants listed above were grown in complete dark at 20+2°C on MS containing K (10 mg L–1) + GA (1 mg L–1) to observe the influence of various types of cuts on callussing and morphogenesis. The response were noted after three weeks of culturing.
Effect of different sugars on formation of callus mass formation: In another experiment shoot apical callus was produced in MS + K, 4; GA, 1; IAA, 1: glutamine, 50 mg L–1 and 3 % sucrose, under 16/8 h light/dark cycle under 4000 lux at 20±2°C. The callus after 2 subcultures (12 week) was grown on the same medium constituents, but sucrose was replaced by other sugar i.e. maltose and fructose. The weight of callus was recorded after 2 month growth.
Effect of maltose, sucrose, mannitol, fructose yeast and malt extract on callussing and subsequent formation of shoot primordia: This experiment was conducted on callus initially obtained from cotyledonary node region with half cotyledons attached to it in MS medium containing BA (5 mg L–1) + 2,4-D (0.5 mg L–1) and fructose 3%. After six weeks, the callus masses, as whole, were transferred to various medium containing either; maltose or sucrose or mannitol or fructose each 3 per cent and additives yeast and malt extract (each 0.1 g L–1). The subculturing medium had reduced salts (1/2 MS) with BA (5 mg L–1) and without 2,4-D. After six week the emerged shoot primordia were counted.
RESULTS
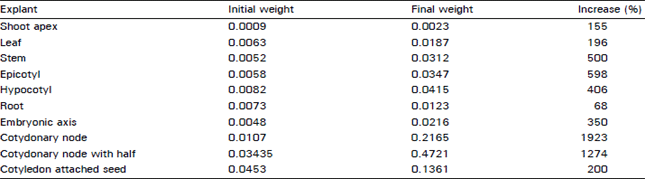
Callussing response of different explants: The percent increase in weight (g) due to callussing of different explants of M-85 after 6 weeks of culture in MS+K and 2,4-D (each 2 mg L–1) with glutamine (5 mg L–1) liquid medium are given in Table 1. The maximum cell proliferation was in the cotyledonary nodes (1923%) and the minimum were the root explants (68%). The next lower percent increase in mass was in the cotyledonary nodes with half cotyledons attached (1274%) and the second next lower gain in weight was in the epicotyl explants (598%). This experiment clearly demonstrated that the first basal nodes and the region adjacent to it had the maximum cell division capability, especially if the explants were from the germinating seedlings.
Optimum size of the explant for callussing: The optimum size of the various explants for callus culture in K + 2,4-D (each 2 mg L–1) are recorded in Table 2 on the basis of vigour and callus mass production leaflet with petiole 0.3 cm, shoot apex 0.2 cm, epicotyl, hypocotyl and root 0.5 cm, cotyledonary node and cotyledonary node with 1/6 cotyledon 0.4 cm, stem with node 0.3 cm were the optimum size of different explants for optimum callussing.
Growth regulator requirements for callussing and shoot regeneration: Growth regulator requirements of light or dark grown cultures of different explants from four day old seedlings were studied for callus induction and formation of shoot primordia, after six week of culture (Table 3). Agar solidified media were used. The best callus response was from the seeds. Cotyledonary nodes developed shoot primordia along with callussing. In the basal nodes, BA with fructose produced more shoot primordia as compared to BA with sucrose. Leaf explants in glutamic acid containing media gave fresh and healthy calli.
Morphogenetic response of the lower stem segments to IBA: The morphogenetic responses of the stem segments (two inches above cotyledons) were recorded; when they were first exposed to IBA(0.01-1.5 mg L–1) for 5 days in dark and then transferred to MS basal medium (Table 4). At 0.01 mg L–1 IBA, shoots developed from buds and roots from the lower stem segment near to cotyledon. The buds developed at IBA concentrations ranging from 0.01 to 0.1 mg L–1 and those elongated into shoots. IBA at 1.5 mg L–1 mostly induced root formation in segment of the lower stem section. However, one to two roots developed from each segment. It is concluded from the data that exposure of stem segment to IBA at these concentrations for five day did not inhibit bud and shoot growth. It also helped in root induction.
Influence of types of cut to explants on callus induction, shoot and root formation: It was regularly observed that cotyledonary stem region from the germinating seedlings had the highest growing capabilities. The surgical cut made on explants from this region had definite influence on callussing and regeneration (Table 5). All these culture were grown in dark in K (10 mg L–1) + GA (1 mg L–1) as in light, the cultures were not as responsive to callus formation, as in dark. When the cotyledons were kept intact with embryonic axes, there was no callus formation. If the cotyledons were cut, then the explant started callussing, otherwise the cotyledons became enlarged with swelling. However, scrapping of the axillary portions with intact cotyledons attached gave rise to vigorous regenerating callus. The shoot and root poles enlarged into shoot and roots.
Effect of different sugars on callus mass: The callus from shoot apex explant was obtained in MS (K, IAA, GA,4,1,1 mg L–1) under light conditions. Shoot apex callus after 2 subcultures, was further subcultured in various sugars combinations for a period of two month in the same medium. It was clear from the results that sucrose alone was the best for callus subculturing as maximum callus weight was obtained with sucrose supplement (Table 6).
| Table 1: | Percent increase in weight (g) due to callusing of different explants of M-85 after 6 week of culture in MS+K and 2,4-D each (2 mg L–1) + Glutamine (5 mg L–1) liquid medium |
 | |
| Table 2: | Optimum size of different explants for optimum callussing* |
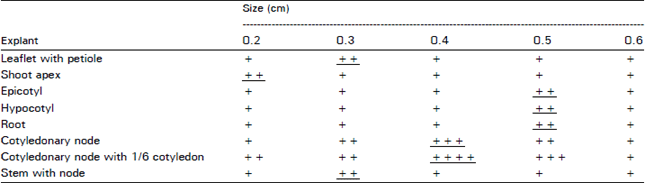 | |
*In MS + K + 2,4-D (each 2 mg L–1) for one month Callussing response: + poor, ++ slow, +++ good, ++++ very good. The underline mark indicate optimum response | |
| Table 3: | Growth regulator requirements for callus growth and shoot regenerations of Light (L) or Dark (D) grown on cultures of various explants from 4-day seedling of Masoor-85 |
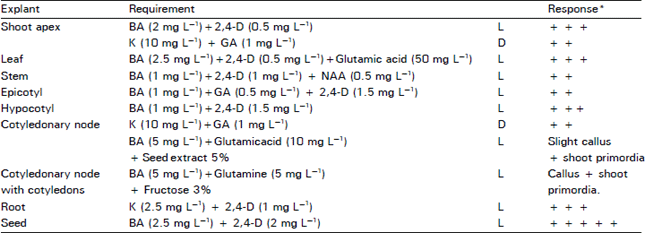 | |
| *Response was after 6 week growth; + + Fair callussing; + + + Good callussing; + + + + + best callus response | |
| Table 4: | Effect of IBA (0-1.5 mg L–1) in dark for five day on morphogenetic responses of stem segment |
 | |
| Masoor-85 plants (2 months). 3 sections of plant part within 2" distance above the cotyledons | |
| Table 5: | Influence of types of cut, made on explants, on callus induction and other morphogentic responses (after 3 week growth) |
 | |
| Culture medium MS + K (10 mg L–1) + GA (1 mg L–1) in complete dark L - longitudinal, T-transverse, EA-embryonal axis, Cot - cotyledons, C - callus, S - Shoot, R - root | |
| Table 6: | Effect of different sugars on growth of callus* (0.5 g) taken from second subculture (2 month old) |
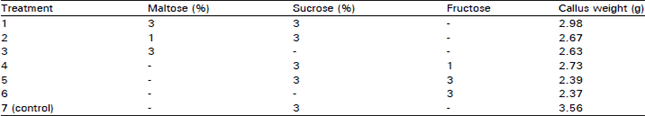 | |
| Culture medium MS (K, IAA, GA, 4, 1, 1 mg L–1) + Glutamine 50 mg L–1. *Explant was shoot apex. | |
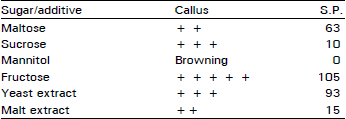
| Table 7: | Effect of sugar (3%) and yeast and malt extract (0.1 g L–1) on callus growth* (explant cotyledonary node) and formation of shoot primordia (S.P.) |
 | |
| *1/2 MS + BA 5 mg L–1 | |
Effect of maltose, sucrose, mannitol, fructose, yeast and malt extract on callus growth and formation of shoot primordia: This experiment was done on cotyledonary node as explant. The total callus growth after six week in 1/2 MS + BA (5 mg L–1) + 2,4-D (0.5 mg L–1) + 3% fructose without removing any material of callus portion was tested on various additives as yeast, malt extract (each 0.1 g L–1) and sugars (3%) enhanced the callussing process and number of shoot primordia (Table 7). The culture mass increase was maximum with fructose and the number of shoot primordia formed were also maximum in fructose. Maximum elongated shoots were grown in the initial culture but the number of shoot formed decreased with the subsequent subculturing in fructose. However, the cultures continued forming shoot primordia uptill two or three subcultures. These shoot buds were dormant and elongation of shoots from these buds was problematic. Yeast extract also increased the number of shoot primordia. The callus mass did not increase much in maltose, although more shoot primordia were formed as compared to sucrose. Mannitol did not promote any callussing or shoot primordia formation. It induced browning of callus. The fructose in the presence of BA was found better as compared to K and GA combinations (data not reported).
DISCUSSION
Two factor which control and govern the success of plant cultures are explant origin, size and culture environment. It was noticed that growth responses of explants from the germinating seedlings (3-4 days) were much better than that of the older seedlings. Both the type and size of explant were important along with the culture protocol. Too small or too large explants were not very suitable for cultures. In this work, the basal stem portion along with the scrapped basal nodes and some part of cotyledons with normal polarity from the fresh germinating seedlings had maximum callussing and cell proliferation capacity. The developmental stage of the explant can be of tremendous importance. The age of the stock plant, the physiological age of the explant and its developmental stage as well as its size can determine the success of a procedure (Franclet et al., 1987).
Williams and McHughen (1986) working with macrosperma lentil in cultivar "Eston" had reported shoot meristem callus regeneration with K and GA. However, epicotyl and cotyledon calli did not regenerate in this medium. Polanco et al. (1988) used three Spanish cultivers, verdina, pardina and castellana and observed that from amongst shoot tip, leaf and node explants the best morphogenetic responses were obtained from nodes and the poorest from the leaves. The explant type has a definite influence on callus regenerations as cotyledonary node explants in soybean (Jackson and Hobbs, 1990). Surgical cut or injuring the cotyledons of eight genotypes of soyabean were examined, cutting expressed higher ability for somatic embryogenesis than the whole cotyledons (Nadolska-Orczyk and Orczyk, 1994). In mungbean regeneration, the number of attached cotyledons and removal of a portion of one or both cotyledons affected shoot formation (Gulati and Jaiwal, 1994). Cotyledonary region has a very good regeneration potential not only in seed legumes but in other species as well. In Camellia reticulata L., plantlets were regenerated by direct and indirect somatic embryogenesis from immature zygotic embryos, cotyledonary sections and embryonic axis cultures (Plata and Vieitez, 1990).
Since fructose enhanced shoot primordia formation, various sugars were supplemented individually to see if they were having any effect on cell growth during callus subculturing. It was found that fructose and maltose had no better effect on callussing than sucrose. However, callussing and shoot primordia formation could be increased with the addition of seed extract in all callus inducing media including K and GA. The later medium had regeneration potential for more than one year as long as the callus continued growing in it. Almost all the explants from the germinating seedlings formed callus in K and GA and also regenerated buds. The number of regenerated primordia depended on callus surface. For the larger callus surface, there were more regenerations and the surface increased with the increase in mass by subculturing passages. Fructose gave maximum shoot buds in comparison to yeast extract, sucrose, maltose and malt extract respectively.
However, mannitol had no positive effect on lentil callus which turned brown. If the seeds of Masoor-85 were incubated in high cytokinin for 5 to 6 weeks, the basal nodes started proliferating shoot primordia and in some cases, apical meristem started multiplication and produced shoot buds which was a rare event in case of lentil cultures. Masoor-85 scrapped cotyledonary nodal explants along with 1/6th cotyledons attached on K and GA medium containing lentil seed extract gives regenerating callus in the dark. The mass of the callus can be increased by subculturing as more regenerations are associated with more callus mass. Infact, shoot bud number enhanced by scrapping.
Shoot buds from lower stem sections when grown elongated in MS basal medium but after they had been exposed to IBA for 5 days in the dark. However, in these cultures, the basal stem parts had rooting capacity.
One of the aims of this work was to observe the growth regulator interaction with meristem in Masoor-85 and it was found that K and BA both are capable of inducing shoot buds, the 2,4-D increased cell proliferations in explants by callussing. However, K and GA induced slow growing callus in Masoor-85 explants when kept in dark conditions. The dark period was essential for callussing with K and GA. This callus had regeneration ability when shifted to light. The callus continued proliferating with callus mass and shoot buds. Callus and shoot buds were also enhanced by the addition of lentil seed extract to K and GA medium. Addition of seed extract improved callusing and regeneration in Masoor-85 as it had been reported to improve regenerations in case of Phaseolus vulgaris L., where plants were regenerated on a medium containing bean seed extract of an undefined composition (Crocomo et al., 1976).
REFERENCES
- Crocomo, O.J., W.R. Sharp and J.E. Peters, 1976. Plantlet morphogenesis and the control of callus growth and root induction of Phaseolus vulgaris with the addition of a bean seed extract. Zeitschrift Pflanzenphysiologie, 78: 456-460.
CrossRefDirect Link - Franclet, A., M. Boulay, F. Bekkaoui, Y. Fouret, B. Verschoore-Martouzet and N. Walker, 1987. Rejuvenation. In: Cell and Tissue Culture in Forestry, Volume 1: General Principles and Biotechnology, Bonga, J.M. and D.J. Durzan (Eds.). Chapter 14, Springer, Dordrecht, The Netherlands, ISBN: 978-90-481-8300-5, pp: 232-248.
Direct Link - Gamborg, O.L., R.A. Miller and K. Ojima, 1968. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res., 50: 151-158.
CrossRefPubMedDirect Link - Gulati, A. and P.K. Jaiwal, 1994. Plant regeneration from cotyledonary node explants of mungbean (Vigna radiata (L.) Wilczek). Plant Cell Rep., 13: 523-527.
CrossRefDirect Link - Jackson, J.A. and S.L.A. Hobbs, 1990. Rapid multiple shoot production from cotyledonary node explants of pea (Pisum sativum L.). In vitro Cell. Dev. Biol., 26: 825-835.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nadolska-Orczyk, A. and W. Orczyk, 1994. New aspects of soybean somatic embryogenesis. Euphytica, 80: 137-143.
CrossRefDirect Link - Plata, E. and A.M. Vieitez, 1990. In vitro regeneration of Camellia reticulata by somatic embryogenesis. J. Hortic. Sci., 65: 707-714.
CrossRefDirect Link - Polanco, M.C., M.I. Pelaez and M.L. Ruiz, 1988. Factors affecting callus and shoot formation from in vitro cultures of Lens culinaris Medik. Plant Cell Tissue Org. Cult., 15: 175-182.
CrossRefDirect Link - Williams, D.J. and A. McHughen, 1986. Plant regeneration of the legume Lens culinaris Medik. (lentil) in vitro. Plant Cell Tissue Org. Cult., 7: 149-153.
CrossRefDirect Link