
K.M. Makkouk
Virology Laboratory, Germplasm Program, International Center for Agricultural Research in the Dry Areas (ICARDA), P.O. Box 5466, Aleppo, Syria
S.G. Kumari
Virology Laboratory, Germplasm Program, International Center for Agricultural Research in the Dry Areas (ICARDA), P.O. Box 5466, Aleppo, Syria
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 4 | Page No.: 303-306







ABSTRACT
Antisera against purified particles of tobravirus isolates from Libya (LYV66-91) and Algeria (AlgRl 0) were produced and used for their serological comparison with other isolates of pea early-browning tobravirus (PEBV), including one of the broad bean yellow band serotypes (PEBV:BBYB) and with the PRN strain of tobacco rattle tobravirus (TRV:PRN). DASELISA, DAC-ELISA and dot-blot BLISA showed that the Algerian and the Libyan isolates represent two different serotypes. In DAS-BLISA, the Libyan isolate was similar to the Dutch isolate of PEBV (isolate El 16). In DAC-ELISA, the Algerian isolate reacted strongly with PEBV:BBYB and the TRV:PRN antibodies but not with those to the Libyan or the Dutch isolates of PBBV. Using dot-blot ELISA, the Libyan isolate was serologically closely related to the Dutch isolate of PBBV and distinct from the Algerian isolate of PEBV, PEBV:BBYB and TRV.PRN.
PDF Abstract XML References Citation
How to cite this article
K.M. Makkouk and S.G. Kumari, 1998. Further Serological Characterization of Two Tobravirus Isolates from Algeria and Libya. Pakistan Journal of Biological Sciences, 1: 303-306.
DOI: 10.3923/pjbs.1998.303.306
URL: https://scialert.net/abstract/?doi=pjbs.1998.303.306
DOI: 10.3923/pjbs.1998.303.306
URL: https://scialert.net/abstract/?doi=pjbs.1998.303.306
INTRODUCTION
Pea early-browning tobravirus (PEBV) was originally described from Western Burope (Bos and van der Want, 1962; Gibbs and Harrison, 1964) as a soil-(nematode-) and seed-borne virus able to cause severe disease in pea and often symptomlessly infecting other plant species, particularly legumes. Russo et al. (1984) isolated a virus from faba bean in Southern Italy, broad bean yellow band tobravirus (BBYBV), which they considered a distinct tobravirus, serologically unrelated to PEBV. Soon thereafter, Robinson and Harrison (1985) provided evidence that BBYBV is a serotype of PEBV, designated PEBV:BBYB, rather than a distinct tobravirus.
Within the framework of the ICARDA research on viruses of cool season food legumes in West Asia and North Mrica (WANA) (Makkouk et al., 1988) and through a linkage project with the Research Institute for Plant Protection (IP0- DLO) in Wageningen, two tobravirus isolates were recently recovered from faba bean (Viciafaba) in Libya and Algeria. They were studied at IPO-DLO and found to be different from each other. The Libyan isolate (LYV66-91) was closely related to the Dutch type strain of PEBV (El 16) (Bos et al., 1993), and the Algerian isolate (AlgR10) was related to, but biologically different from the Italian type strain of PEBV:BBYB serotype of PEBV (Mahir et al., 1992). LYV66- 91 reacted in ISEM with antiserum to El 16 but not with that to BBYB (Bos et al., 1993), whereas AlgR10 reacted with antiserum to BBYB only (Mahir et al., 1992).
This paper describes the production of antisera against the LYV66-91 and A1gR10 isolates as diagnostic reagents for flirthersurveying in the region, for germplasm testing for seed infection and for resistance screening. It flirther characterizes the serological relationship between these isolates and strains of PEBV and TRV, and evaluates the useflilness of the antisera produced for sensitive detection of the two isolates using different ELISA variants.
MATERIALS AND METHODS
Virus isolates and maintenance: The Libyan isolate (LYV66- 91) was originally obtained by mechanical inoculation from a faba bean leaf sample collected in 1991 near Tajura, Libya, and later identified as PEBV (Bos et al., 1993). The Algerian isolate (AgR10) was derived from a sample of faba bean leaf provided by A. Oufiroukh, Institute National de Protection des Vegetau (INPV), El-Harrach, Algiers, Algeria, identified earlier as a deviant isolate of PEBV:BBYB serotype of PEBV (Mahir et al., 1992). Both isolates were preserved in desiccated leaves and stored over calcium chloride (CaCl2) in the !PO collection and at ICARDA. The LYV66-9 1 isolate was propagated and maintained in Nicotrana clevelandii, and AlgR10 in faba bean or N. rustica by serial mechanical inoculation.
Virus isolates used for comparative serological tests were PEBV (E413) provided by Dr. L. Bos, The Netherlands; the PRN strain of tobacco rattle virus (TRV:PRN) and PBBV:BBYB supplied by Dr. D.J. Robinson, Dundee, 5CR’, Scotland.
Virus punfication: LYV66-91 and AlgR10 were purified from infected N. clevelandli and N. rustica, respectively, using a method slightly modified from those described by Cooper and Mayo (1972) and Lister and Bracker (1969). Both isolates were purified by using the same procedure.
Systemically-infected leaves were harvested 15-20 days after plant inoculation and ground at room temperature in 0.1 M potassium phosphate, pH 7.4, containing 0.01 M citric acid (trisodium) and 0.1 per cent thioglycollic acid (1 g tissuel 2 ml buffer). The homogenate was kept at -20°C for 2-5 days, thawed overnight at 4°C, and then heated at 50°C for 10 mm. The sap was expressed through cheesecloth, mixed with a quarter volume of a 1:1 mixture of chloroform:butanol, stirred at room temperature for 15 mm and then centrifliged at 10,000 rpm for 10 min (Sorvall SS-34). The virus particles were concentrated by precipitation with 8 percent polyethylene glycol plus 1 percent NaCI at 4°C for 3 h, then centrifligation at 12,000 rpm for 20 mm and the resulting pellets were resuspended overnight at 4°C in 0.02 M potassium phosphate (PP), pH 7.4. The preparation was then centrifuged at 12,000 rpm for 10 min and 50 ml of the supernatant fluid were placed on 15 ml ‘cushions’ of 20 percent (WIV) sucrose in PP buffer and centrifliged at 33,000 rpm for 90 mm (Beckman TY35). The pellets were resuspended in a small volume of PP buffer arid 1 mL aliquots were layered onto sucrose gradients in the same buffer. Sucrose gradients were prepared by using the freezing and thawing method (Davis and Pearson, 1978). The gradients were centrifliged at 30,000 rpm for 50 min (Beckman SW 41), followed by fractionation in an ISCO 640 density-gradient fractionator. Virus-containing fractions were combined, diluted 10-15 times in PP buffer, centrifliged at 33,000 rpm for 90 min, and the pellets obtained were resuspended in a small volume of the same buffer. Virus yields were estimated spectrophotometrically at 260 nm using an extinction coefficient value of 3, as established earlier for PEBV (Harrison, 1970).
Antisera production and antisera used: Antisera for LYV66- 91 and A1gR10 were produced by giving New Zealand white rabbits five weekly intramuscular injections each containing 0.1-0.3 mg virus. Purified virus preparations were emulsified with an equal volume of Freund’s complete adjuvant for the first injection and with Freund’s incomplete adjuvant for subsequent injections. One booster injection was given one month after the fifth injection. The rabbits were bled ten times at weekly intervals starting one week after the fifth injection.
Polyclonal antisera used for comparative serological tests were kindly supplied by Dr. L. Bos (Dutch type strain of PBBV isolated from pea (El 16); (Bos and Van der Want, 1962) and Dr_ D.J. Robinson, 5CR’, Dundee, Scotland (TRV:PRN and PEBV:BBYB). In addition, the Fab fragment of TRV:PRN antibodies was provided by Dr. D.J. Robison.
Serological tests
Double-AntibodySandwichELISA(DAS-ELISA).
Immunoglobulins (1gG) were fractionated from LYV66-91 AlgR10 and El 16 antisera using the caprylic acid method (Steinbuch and Audran, 1969). Conjugation of immunoglobulin with alkaline phosphatase and DAS-ELISA procedures were those of Clark and Adams (1977). Plates were coated with I μg/ml of gammaglobulin and the conjugate dilution used was III 000. Absorbance at 405 nm was measured 1 h after addition of the substrate.
When the Fab fragemnt of TRV:PRN was used in DAS-ELISA, the plates were coated with a 1:1000 dilution of such fragment followed by appling the antigen extract. The detecting antibody (TRV:PRN IgG) was mixed with protein A conjugated to alkaline phosphatase and the mixture was diluted 1:1000 in conjugate buffer. Absorbance at 405 nm was measured 12 h after addition of the substrate.
Direct Antigen Coating ELISA (DAC-ELISA). The DAC-ELISA procedure was that of Lommel et al. (1981). All antisera (LYV66-91, AlgR10, El 16, PEBV:BBYB and TRV:PRN) were diluted 1:1000 and the goat anti-rabbit alkaline phosphatase conjugate was diluted 1:2000. Absorbance at 405 nm was measured 30 mm after addition of the substrate.
Dot-blot ELISA. This test was conducted by using chromogenic substrates nitro blue tetrazolium (NBT) and bromo-chloro-indolyl phosphate (BCIP) (Makkouk et al., 1993). All antisera were diluted 1:1000 and the goat anti-rabbit alkaline phosphatase conjugate was diluted 1:2000. Samples were applied on the nitrocellulose membrane (NCM) using a Manifold I System (Schleicher & Schuell). Reactions were evaluated visually 15 min after the addition of the substrate.
Results and Discussion
Virus purification: Following the purifcation procedure described above, particles of LYV66-91 and AlgRIO gave two opalescent bands after centrifugation in sucrose density gradients (Fig. 1). The presence of the virus in both bands was confirmed by its reactivity with PEBV:BBYB antiserum (AlgR10) or to El 16 antiserum (LYV66-91) using the dot-blot test. The A260/280 ratio of the purified preparation of AlgRIO was 1.16, which is close to the value for PBBV:BBYB reported by Russo et al. (1984), The M601280 ratio for LYV66-91 was 1.13 which is close to the value for PEBV reported by Harrison (1973). Assuming an extinction coefficient for PEBV of 3, the yield of purified virus was calculated to be 4.6 mg/kg of leaf for AI∼10 and 7.5 mg kg–1 for LYV66-91 (average of three preparations).The UV absorbtion profiles (Fig. 1) for the two isolates, also suggested that the virus bands are very well separated from non-virus components. In addition, the ratio of the long IL) to short (S) particles in the Algerian isolate was higher than that of the Libyan isolate. This result was consistent in the three preparations made independently. However, since the Algerian isolate was purified from N. rustica and the Libyan isolate from N. clevelandii, it is not clear whether the difference in LIS particle ratio is dependent on host or virus strain.
Serology: Results obtained from DAS-ELISA (Table 1) indicated that the Libyan and Algerian isolates of PEBV hardly cross react, if at all. The Libyan isolate of PEBV (LYV66-91) reacted strongly with antisera to the homologous virus or to El 16 but not with that to AlgR10. In contrast AIgRIO reacted strongly with its homologous antiserum but not with LYV66-91 or El 16 antisera. These reactions suggest that the Libyan isolate is serologically closely related to the Dutch PEBV isolate but the Algerian isolate is not. Results obtained also suggested that the antisera produced against the Algerian and Libyan isolates are of good quality as indicated by the high specificity and the low heterologous and healthy reactions observed.
When five tobravirus antisera were used in a comparative study (Table 2), reactions obtained showed again that the Libyan isolate is serologically closely related to the Dutch isolate but not to the AlgR10, PBBV:BBYB or the type strain of TRV (TRV-PRN). In contrast, the Algerian isolate reacted strongly with the homologous antiserum and with antisera to PEBV:BBYB and TRV-PRN and no reaction was obtained against antisera to the Libyan isolate or the Ducth strain of PEBV (El 16). Dot-blot ELISA also clearly showed that the Algerian and the Libyan isolates of PEBV strongly differ serologically (Fig. 2).
| Table 1: | Detection of LYV66-91 and AlgR10 in leaf extracts and purified preparations when tested by DAS-ELISA |
 | |
| *A405 nm alues were recorded after 1 h of substrate incubation | |
| Table 2: | Differentiation among PEVB isolates by their reaction with six polyclonal antibodies when tested by DAC-ELISA, and DAS-ELISA using the Fab fragment in case of TRV:PRN |
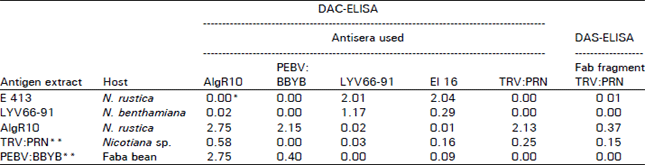 | |
*A405 values were recorded after 30 min. of substrate incubation for DAC-ELISA and 24 h for DAS-ELISA (TRV:PRN, Fab fragment), and after substrating healthy values, which ranged between 0.11-0.37 for Nicotiana spp. And 0.12-0.38 for faba bean. **Extracted tissue for these viruses was sent to us by mail from SCRI, and possibly partially degraded before use. | |
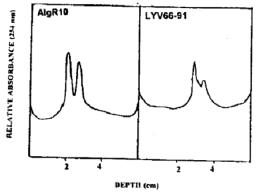 | |
| Fig. 1: | UV (254 nm) absorption profiles of purified virus preparations obtained from N. rustica infected with Algerian isolate (AlgR10) or N. cleyelandii infected with Libyan isolate (LYV66-91) after centrifugation on sucrose gradients at 30,000 rpm for 50 min. (Beckman SW41). |
However, the Algerian isolate of PEBV was serological close to PEBV:BBYB but not identical. AlgR10 antiserum reacted strongly with the homologous antigen and with PEBV:BBYB and not as strongly with TRV-PRN. Likewise, PEBV:BBYB antiserum reacted with AlgR10 and PEBV:BBYB antigens but not with TRV-PRN.
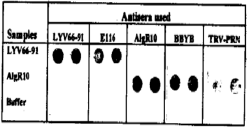 | |
| Fig. 2: | Reaction of purified preparations (1 μg/ml) of AlgR10 and LYV66-91 with five polyclonal antibodies when tested by Dot-blot ELISA. |
These reactions suggest that AlgR10 has some serological relatedness to the type strain of TRV, but PEBV:BBYB does not. Some differences were also reported in their host reactions: PEBV:BBYB and AlgR10, both infect faba bean systemically but they differ appreciably in their reaction on N. rustica, N. tabacum, and Petunia hybriaa (Mahir et al., 1992) and the symptoms observed in faba bean were less severe than those described by Russo et al. (1984) for BBYBV. The Algerian isolate produced a very clear systemic line pattern on N. rustica and N. tabacum White Burley, whereas the Libyan isolate did not produce symptoms on these species (Bos et al., 1993; Mahir et al., 1992).
ACKNOWLEDGMENTS
The authors would like to thank Dr. L. Bos, IPO: DLO, The Netherlands and Dr B. Harrison, SCRI, Scotland for their critical review of the manuscript and their valuable comments.
REFERENCES
- Bos, L., M.A.M. Maliir and K.M. Makkouk, 1993. Some properties of pea early-browning tobravirus from faba bean (Vicia faba L.) in Libya. Phytopath. Medit., 32: 7-13.
Direct Link - Bos, L. and J.P.H. van der Want, 1962. Early browning of pea, a disease caused by a soil- and seed-borne virus. Tijdschrift Over Plantenziekten, 68: 368-390.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Cooper, J.I. and M.A. Mayo, 1972. Some properties of the particles of three tobravirus isolates. J. Gen. Virol., 16: 285-297.
CrossRefDirect Link - Davis, P.B. and C.K. Pearson, 1978. Characterization of density gradients prepared by freezing and thawing a sucrose solution. Anal. Biochem., 91: 343-349.
CrossRefPubMedDirect Link - Gibbs, A.J. and B.D. Harrison, 1964. A form of pea early-browning virus found in Britain. Ann. Applied Biol., 54: 1-11.
CrossRefDirect Link - Lister, R.M. and C.E. Bracker, 1969. Defectiveness and dependence in three related strains of tobacco rattle virus. Virology, 37: 262-275.
CrossRefPubMedDirect Link - Lommel, S.A., A.H. McCain and T.J. Morris, 1981. Evaluation of indirect enzyme-linked immunosorbent assay for the detection of plant viruses. Phytopathology, 72: 1018-1022.
CrossRefDirect Link - Mahir, M.A.M., M. Fortass and L. Bos, 1992. Identification and properties of a deviant isolate of the broad bean yellow band serotype of pea early-browning virus from faba bean (Vicia faba) in Algeria. Netherlands J. Plant Pathol., 98: 237-252.
CrossRefDirect Link - Makkouk, K.M., L. Bos, O.I. Azzam, S. Koumari and A. RizkaUab, 1988. Survey of viruses affecting faba bean in six arab countries. Arab J. Plant Protect., 6: 53-61.
Direct Link - Makkouk, K.M., H.T. Hsu and S.G. Kumari, 1993. Detection of three plant viruses by dot-blot and tissue-blot immunoassays using chemiluminescent and chromogenic substrates. J. Phytopathol., 139: 97-102.
CrossRefDirect Link - Robinson, D.J. and B.D. Harrison, 1985. Evidence that broad bean yellow band virus is a new serotype of pea early-browning virus. J. Gen. Virol., 66: 2003-2009.
Direct Link - Russo, M., D. Gallitelli, C. Vovlas and V. Savino, 1984. Properties of broad bean yellow band virus, a possible new tobravirus. Ann. Applied Biol., 105: 223-230.
CrossRefDirect Link - Steinbuch, M. and R. Audran, 1969. The isolation of IgG from mammalian sera with the aid of caprylic acid. Arch. Biochem. Biophys., 134: 279-284.
CrossRef