
M. Ashraf
Biochemistry Laboratory, Department of Chemistry, Islamia University, Bahawalpur, Pakistan
S. Nisar
Biochemistry Laboratory, Department of Chemistry, Islamia University, Bahawalpur, Pakistan
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 4 | Page No.: 287-290







ABSTRACT
Seeds of Salvadora oleiodes (locally called Jhal or Pelun) were tested for their germination performance and membrane leakage properties during early hours of imbibtion. The optimal germination temperature determined was 30°C wherein 85 percent germination was obtained within 5 days of imbibition. When seeds with testa (intact) or without testa (‘naked’) were immersed in water and electrical conductance (E.C.) measured, higher E.C. values were obtained in ‘naked’ seeds than the intact ones. Potassium ions in leachates of 1 h imbibed seeds were 14-18 ppm/seed in intact and 133 ppm/seed in ‘naked’ seeds. Inorganic phosphorus contents (Pi) were 61 ppm/‘naked’ seed which remained un-detectable in leachates of intact ones. Less tham 1 ppm/seed Na was detected in leachates of ‘naked’ seeds. Amino acids and carbohydrate contents (33 μmol/seed and 68 μg/seed, respectively) were among organic solutes effluxed during early minutes of imbibition.
PDF Abstract XML References Citation
How to cite this article
M. Ashraf and S. Nisar, 1998. Germination Patterns and Electrolytes Leakage During Imbibition of Intact and ‘Naked’ Seeds of Salvadora oleiodes. Pakistan Journal of Biological Sciences, 1: 287-290.
DOI: 10.3923/pjbs.1998.287.290
URL: https://scialert.net/abstract/?doi=pjbs.1998.287.290
DOI: 10.3923/pjbs.1998.287.290
URL: https://scialert.net/abstract/?doi=pjbs.1998.287.290
INTRODUCTION
The process of Imbibition is well understood and involves repair and restoration of damaged cell membrane integrity during early minutes of contact between the seed and the surrounding water (Simon and Harun, 1972; Simon 1984; Bewley and Black, 1986). Membrane damage occurring during seed maturation and desiccation is higher in low vigour seeds and cell deaths may take place due to rapid uptake of water. It is this time during which solutes leak out of the cells. Many substances such as amino acids, organic acids, sugars, phenolics, phosphate and potassium ions, gibberellic acid and proteins are leaked and rate of leakage is not the same for each substance. The increasing in electrical conductivity (E.C.) in leachates of imbibing seeds is due to the increasing leakage of these electrolytes (Simon, 1984). The leakage is the most rapid in the first moments of imbibition and the rate slows down until 10-20 min and steady state condition is reached in the next half hour. Based on these measurements, Pandey (1988) has developed a useful, rapid, simple, inexpensive and highly predictive vigour test of French beans and Williams et al. (1995) have studied the effect of weathering on mung beans.
Intact seeds do not leak or leak little in the surrounding medium. When seed tasta is removed, more leakage is observed in the ‘naked’ seeds (Simon and Harun, 1972; Simon and Mathavan, 1986). Pea seeds after removal of seed testa show an immediate and rapid leakage of potassium and other electrolytes. Increased leakage of organic metabolites from testa-free or deteriorated seeds might indirectly enhance their capacity by encouraging the growth of rhizosphere microorganisms. The symptoms of low vigour and reduced viability could result from fundamental changes in membranes and macromolecules (Bewley and Black, 1986). Increased electrolytes leakage has also been observed in dehydrated isolated axes and an inverse relationship has been found between germination and electrolytes leakage (Senaratna and McKersie, 1983).
Salvadora oleiodes is an evergreen medicinal tree of saline soils (Kirtiker and Basu, 1991). Its seeds are blackish with a tough seed coat. The germination patterns at different temperature, optimum germination and percent seed germination has not been reported earlier. Neither is known about the imbibitional leakage and nature of solutes effluxed during early hour of imbibition. The reported work is an extension of our previous work on acceleration of seed germination of endangered and slow germinating/growing Cholistan desert plant species (Ashraf et al., 1998; Ashraf and Hussain, 1998). Both intact and ‘naked’ seeds have been employed to exhibit germination patterns and monitor the rates and nature of inorganic and organic species leaked during early hours of imbibition. The results indicate that K, inorganic phosphorus, amino acids and carbohydrates are among the dominant species effluxed duting early minutes of imbibition.
MATERIALS AND METHODS
Seeds of Salvadora oleiodes were collected directly from trees in June-July, 1996, from Lal Suhanra National Park Bahawalpur, Pakistan. All chemicals used were purchased from Merck and BDH. Conductivity meter, Milwaukee-CON 1000 was used for the measurement of electrolytes leakage during imbibition. Corning flame photometer was used for the determination of Na and K.
Germination tests were performed by placing the sets of 10 seeds on presoaked double layered filter papers in glass petri dish at the given temperature. Number of seeds germinated were noted after 24 h. Protrusion of radical marked the seed ‘germinated’.
10-20 seeds were immersed in 10 mL double distilled water for the given time and E.C. was measured by immersing the pre-calibrated electrode in the leachate. These leachates were then used for the measurement of K and Na ions by flame photometer after suitable dilutions, and for the determination of inorganic phosphorus by molybdenum method (Plummer, 1987). Amino acids and carbohydrate contents were measured by ninhydrin and anthrone methods (Plummer, 1987). Leachates after bench centrifugation were placed in 0.5 cm quartz cuvette and read at O.D254 nm for uv-absorbable material by uv-visible spectrophotometer (Jasco).
RESULTS
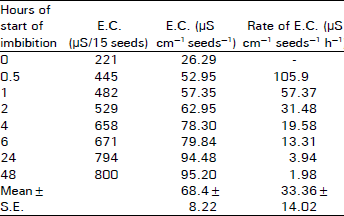
Salvadora oleiodes seeds are blackish weighing 24.28±2.3 mg/seed with moisture contents of 6.05 percent, nitrogen contents of 33.3±2.1 mg g–1 fresh weight and protein contents of 207.8±55.0 mg g–1 fresh weight (Nisar, 1997). A rapid increase in fresh weight of imbibing seeds is exhibited during the initial hours of imbibition. However, after 24 h, the process of uptake of water by the seed is slowed down (Fig. 1). Results of E.C. measurements show that dry seeds on imbibition exhibit E.C. of 26.29 μS cm–1 seed–1 which is doubled within 30 min of imbibition and reaches 4-fold value in the next 4 h (Fig. 1, Table 1). However, decrease in the rate of E.C. as measured in μS cm–1 seed–1 h–1 of 30 min imbibed seed is 105.9 with a 2-fold decrease in the rate in the next 30 min and about 4-fold decrease in 120 min of imbibition until the rate is 3.94 cm–1 seed–1 h–1 after 24 h imbibition (Fig. 1, Table 1).
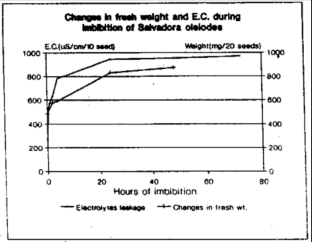 | |
| Fig. 1: | Changes in fresh weight and E.C. during imbibition of seeds of S. oleiodes. 20 seeds were placed in water and increase in fresh weight of imbibing seeds was monitored. For the measurements of E.C., seeds were immersed in 10 mL double distilled water and E.C. measured after given intervals with a precalibrated conductivity meter |
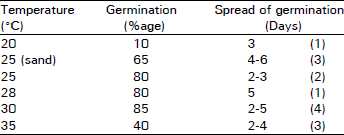
Seed germination: Maximum percentage germination was obtained at 30°C wherein 85 percent germination was achieved within 10 percent error (Table 2). At 25 and 28°C with or without sand (and or light, data not shown), 65-80 percent germination was recorded. However, as the temperature increased to 35°C, 40 percent seed germinated. These results show that 30°C is the optimal germination temperature for S. oleiodes and seed germination is not under photochemical control. Results also show that 85 percent germination is achieved within 4 days starting from 2nd day of imbibition and 80 percent germination is obtained in a single day after 2 days of imbibition at 25°C. Therefore, temperature range of 25-30°C is acceptable germination limit for these seeds. Testa-free ‘naked’ seeds did not show germination.
| Table 1: | E.C. of Salvadora oleiodes at 25°C during initial hours of imbibition. 15 seeds were immersed in 10 mL double distilled water and E.C. measurements of leachates were taken (n = 2). |
 | |
| Table 2: | Spread of germination of seeds at varying temperature regimes. Sets of 10 seeds were placed on presoaked double filter papers in glass petri-dishes at given temperatures. Figures in parenthesis () represent total number of days during which all seeds germinated. (n = 3. S.E is <10%). |
 | |
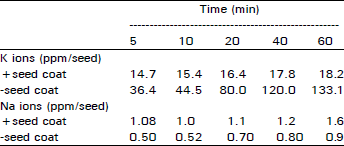
Inorganic solutes in leachates: Changes in the levels of potassium, sodium and inorganic phosphorus in leachates during eaely hours of imbibition were measured as given in Table 3, 4. Intact seeds efflux little K and Na ions in their leachates (Table 3). 14.7 ppm/seeds K is present after 5 min of imbibition which reaches 18.2 ppm/seed in an hour. However, Na levels are 1.08 ppm/seed after 5 min and reach 1.6 in 60 min of imbibition. When these studies were extended for seeds with removed testa, i.e., ‘naked’ seeds, pronounced changes were noticed in K ions compared with Na ions (Table 3). ‘Naked’ seeds effluxed 36.4 ppm/seed K ion in 5 min of imbibition which reached 80 ppm/seed in 20 min and 133.1 ppm/seed in 60 min, an overall increase of 4-fold. Na levels in ‘naked’ seeds decreased by half to 0.5 ppm/seed and remained less than 1 ppm/seed within 60 min duration of imbibtion. These results indicate that seed testa was also contributing to the loss of Na ions during imbibition.
Pi levels in leachates of intact seeds of 60-100 min of imbibition were not detectable (data not shown). However, ‘naked’ seed exhibited levels of 7 ppm/seed at 5 min of imbibition to 61 ppm/seed after 30 min of imbibition (Table 4). These levels indicate high turnover rate of inorganic phosphorus during early hours of imbibition (Bewley and Black, 1986).
| Table 3: | Determination fo Na and K ions in leachate of S. oleiodes with and without seed coats during the initial hours of imbibition. 10 seeds were immersed in 10 mL double distilled water for the given time. Date is mean of two independent experiments. S.E. is < 5% |
 | |
| Table 4: | The amount of inorganic phoshorus (Pi, ppm/seed), amino acids μmol/seed) and carbohydrates (μg/seed) in leachates during imbibition of ‘naked’ seeds. 10-20 seeds were immersed in 10 mL double distilled water and contents determined in the leachates. Results are mean of 2 3 independent experiments. Error is S.E.M. |
 | |
Measurement of uv-absorbable material in leachates: Data on uv-absorbance of leachates reveals a steady increase in the uv-absorbing material in leachate upto 7 h of imbibition of intact seeds (Fig. 2) indicating release of organic compounds from within the testa. However, changes in leachates of ‘naked’ seeds were too high to be determined but were practicable for the measurements of amino acids and carbohydrates contents which were taken as indicators if increased uv-absorbance values.
Organic solutes in leachates: Amino acid and carbohydrate contents of intact seeds were not detected though little increase in uv-absorbable material during early hour of imbibition has been seen (Fig. 2). However, leachates of ‘naked’ imbibing seeds exhibited increasing profiles of amino acids and carbohydrate contents and results are given in Table 4. Dry seeds at zero minute exhibited amino acid contents of 2 μmol/seed which increased 6-fold in 10 min and reached 33 μmol/seed in the next 30 minutes of imbibition. ‘Naked’ seeds leached 10 μg/seed carbohydrates at zero time and a 4-fold increase was seen in the next10min of imbibition. In 30 min of imbibition, 86 μg/seed carbohydrates have been detected (Table 4). These studies reveal that only testa free seeds leak amino acids and carbohydrates in the detectable limits though their rates are different during imbibition.
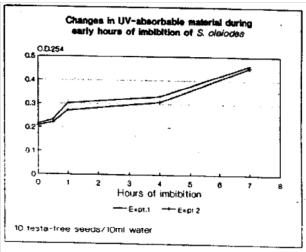 | |
| Fig. 2: | Changes in uv-absorbable material in leachates of imbibing seeds. 10 seeds were immersed in 10 mL double distilled water. Leachates were centrifuged and absorbance measured at 254 nm |
DISCUSSION
Desert plants seeds possess hard testa which is often associated with dormancy. When seeds are dispersed in sand, processes invoiving breaking of dormancy are completed. Like other trees, S. oleiodes seeds possess hard seed testa which is difficult to break and seeds may have some form of dormancy, though some seeds get germinated soon after dispersal. The present study was aimed, i) to obtain optimal germination temperature range for the seeds, ii) to abserve the imbibitional patterns of seeds with or without testa and, iii) to abserve the nature of electrolytes effluxed during early hours of seed imbibition.
ROptimal germination temperatures of S. oleiodes seeds are 25-30°C which have not been found different from other species of Cholistan desert (Nasim et al., 1996). The presence of hard seed coat helps in maintaining the seed vigour and viability throughout seed storage. Results show that seeds with seed testa exhibiting 80-85 percent germination leaked little on imbibition. However, testa-free ‘naked’ seeds exhibited maximum levels of amino acids and carbohydrates in their leachates and showed no germination. It suggests that the presence of seed testa restricts the efflux of organic (and inorganic) solutes which otherwise may help microorganisms to invade for nitrogen and carbon sources. Further, the deterioration and decay of the seed testa may have implications in desert soils to propagate rhizosphere during early hours of imbibition. Similar findings have been demonstrated by Simon (1984). Among inorganic ions effluxed during early hour of imbibition, K ions were present in leachates of both intact and ‘naked’ seeds though only 18.2 ppm/seed in 1 h leachates of intact seeds. Leachates of ‘naked’ seeds have shown 36.4 ppm/seed of k ions which rose 3.7 fold (133 ppm/seed) within 1 h of imbibition. Leakage of K ions looks a fundamental property of seeds since a number of other seeds have shown similar profiles furing early hours of imbibition (Simon, 1984; Ashraf et al., 1998). Scarified lotus seeds efflux 13% K during the initial 2 h of imbibition whilst celery seeds lose more than 60 percent originally present in teh seed within 1 h of imbibition (Simon and Mathavan, 1986). The presence of K ions is the major inorganic species responsible for increase in E.C. of leachates. As for as the presence of inorganic phosphorus (orthophosphate (Pi) species) is concerned, a 30 times increase in Pi content (from 2 to 61 μmol/sees) within 30 min of imbibition has been observed which indicates higher turnover rate of this species. Like K ions, Pi has also been increased during imbibition of other seeds (Simon, 1984: Bewley and Black, 1986). However, the contribution of Na ions is little both by the intact and ‘naked’ seeds in increasing E.C. of the leachates (Simon, 1984).
Amino acids and sugar contents in leachates of various seeds like lotus, wheat, pea, beans etc. have been determined (Simon, 1984; Simon and Mathavan, 1986; Ashraf et al., 1998). The data presented here is not different from others except that the retes of leakage of amino acids and sugars are different, that is , 16 times increase in amino acid contents occurs in 30 min of imbibition (from 2 to 33 μmol/seed) and this value for sugars is 8.6 fold (from 10 to 86 μg/seed). Differential behaviour of these electrolytes has already been studied and nature, vigour, physical damage to seeds and some genetic facrots like whether the seeds contain starch as reserved food or fats or proteins, all these factors determine the extent and rates of leakage of these solutes, including inorganic ones (Simon, 1984; Simon and Mathavan, 1986; Bewley and Black, 1986; Ashraf and Hussain, 1998). A detailed study is needed in demonstrating the individual species in carbohydrates and amino acids and fats which could act as markers for the germinability status of the given seed. What is the amount of other inorganic and organic molecules which have not been determined in the leachates, is unknown. Studies are ongoing on the quantification of other organic molecules and class of molecules effluxed in leachates of early imbibitional period of an under-developed seedling.
ACKNOWLEDGMENTS
Thanks are due to Mr. Abdul Rashid, Regional Agriculture Research Institure, Bahawalpur, for a king gift of seeds collected from Lal Suhanra National Park, Bahawalpur, Pakistan. This work was partly funded by Islamia University Research Grant.
REFERENCES
- Ashraf, M. and M.M. Hussain, 1998. Changes in amino acid and carbohydrate contents in leachates of preimbibed wheat seeds. Pak. J. Biol. Sci., 1: 148-151.
CrossRefDirect Link - Nasim, F.U.H., T.F. Shahzadi and M. Ashraf, 1996. A cold shock during imbibition improves germination of Acacia nilotica seeds. Pak. J. Bot., 28: 183-190.
Direct Link - Pandey, D.K., 1988. Priming induced repair in French bean seeds. Seed Sci. Technol., 16: 527-532.
Direct Link - Senaratna, T. and B.D. McKersie, 1983. Dehydration injury in germinating soybean (Glycine max L. Merr.) seeds. Plant Physiol., 72: 620-624.
CrossRefDirect Link - Simon, E.W. and R.R. Harun, 1972. Leakage during seed imbibition. J. Exp. Bot., 23: 1076-1085.
CrossRefDirect Link - Simon, E.W. and S. Mathavan, 1986. The time-course of leakage from imbibing seeds of different species. Seed Sci. Technol., 14: 9-13.
Direct Link - Williams, R.W., R.J. Lawn, B.C. Imrie and D.E. Byth, 1995. Studies on water damage in mungbean. II. Electrical conductivity of seed leachate as an assay of level of damage. Crop Pasture Sci., 46: 901-907.
CrossRefDirect Link