
N. Raja
ABI, Nat. Agric. Res. Centre, P.O NARC, Park Road, Islamabad, Pakistan
R.J. Rowbury
Biology Department (Darwin Building), University College London, Gower Street, London WC1E 6BT, U.K
D.G. Smith
Biology Department (Darwin Building), University College London, Gower Street, London WC1E 6BT, U.K
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 4 | Page No.: 256-258







ABSTRACT
ED1829ColV, I-K 94, a derivative of Escherichia coli K-12, were injured with hydrogen peroxide, acetic and formic acid for various length of time at acidic pH(s) in Nutrient Broth # 2. The acid injured organicism recovered significantly better in the media containing phosphate than the media without phosphate. Phosphate helped in plating efficiency of the inured bacterial population and somehow expedite the recovery process of the acid stressed organisms.
PDF Abstract XML References Citation
How to cite this article
N. Raja, R.J. Rowbury and D.G. Smith, 1998. Recovery of Injured Escherichia coli. Pakistan Journal of Biological Sciences, 1: 256-258.
DOI: 10.3923/pjbs.1998.256.258
URL: https://scialert.net/abstract/?doi=pjbs.1998.256.258
DOI: 10.3923/pjbs.1998.256.258
URL: https://scialert.net/abstract/?doi=pjbs.1998.256.258
INTRODUCTION
Chemical food preservation (sorbate, benzoate, propionate, acetate, and lactate) have been used widely during the greater part of this century. Most of these food additives inhibit the growth of unwanted organisms, usually without killing them and are ingested by humans without any apparent toxicity (Freese et al., 1973).
Acids alone or in combination with other physical and chemical agents in food industry, cause either damage to the existing microbial population or make conditions unsuitable for unwanted micro-organisms (Baird-Parker, 1980). In a situation when sublethal (acid) stress is applied to a bacterial population, this may cause injury to the microbial cells. The injured organisms could recover, if the acid-stress is removed, and they may form colonies on low stress-media but unable to form colonies on stressing -media (Przybylski and Witter, 1979).
Toxicity of organic acids was studied by Sinha (1986), where the wild type strain and four repair-deficient strains (uvrA6, uvrB5, recA56, and polA1) of E. coli, in their stationary growth phase, were exposed to organic acids (acetic acid, lactic acid and p-aminobenzoic acid) at pH 3.5. The sensitivity exhibited by the polA strain was much higher than its isogenic pol+ derivatives.
When Escherichia coli K-12 cultures were treated with acidic 0.3 M sodium acetate buffer, the extent of both death and injury of acid-treated cells increased with decreasing pH. Injured cells were able to recover their colony-forming ability in the recovery medium. The recovery was not affected by the presence of inhibitors of protein, cell wall, DNA, or RNA synthesis (Przybylski and Witter 1979).
Alkali-injured cells were found to recover when incubated at 37°C for 2 h in 0.05 M phosphate buffer of pH 8.0, whereas no such liquid holding recovery was observed in recA and lexA mutants. Recovery in phosphate buffer was not affected by metabolic inhibitors (Musarrat and Ahmad 1988).
Dukan et al. (1997) reported that HOCI stressed Escherichia coli resulted in recovery of the in phosphate buffer. The HOCI-stressed population is in large part due to growth of a few culturable cells at the expense of damaged cells. However, comparison of the growth rates of the stressed culturable population and of untreated bacteria growing in filtrate showed significantly faster growth of the stressed cells, a fact not fully compatible with the hypothesis that recovery is only the simple growth of survivors. They suggested, therefore, that in addition to growth of the few culturable stressed cells, there is repair and growth of some mildly injured viable but nonculturable cells.
MATERIALS AND METHODS
Bacteria strain: The strain used was ED1829ColV,I-K94, a derivative of E. coli K-12, which has been described in Hicks and Rowbury (1986).
Media: Oxoid Nutrient Broth No. 2 (NB) was used at 25 g L–1 as growth medium. Its pH was adjusted by the addition of HCI, to required values, where needed. For study of recovery of injured organisms, medium was made up in 0.1 M pH 7.5 phosphate buffer.
Growth conditions: Overnight cultures were prepared by growing the bacterial cells in NB of required pH, at 37°C with aeration. For exponential cultures, the overnight grown cells were diluted into fresh NB of required pH and incubated with shaking, at 37°C, for 2-3 h.
Sensitivity of the organism to putative inhibitory agents: ED1829ColV,I-K94 was grown exponentially for 2-3 h and then treated with various inhibitory agents in NA at a required pH for an appropriate length of time with shaking, at 37°C.
Recovery of injured organisms: ED1829ColV,I-K94 organism were treated with putative inhibitory agents for various length of time, with aeration at 37°C. Samples (1 mL) were withdrawn and diluted into 99 mL of the recovery medium with and without 0.1 M phosphate buffer of pH 7.5, and the cultures were incubated statically at room temperature or 37°C for 2 h prior to plating 0.1 mL of appropriate dilutions on Na plates to determine percentage recovery.
| Table 1: | Effect of incubation in different media on recovery of ED 1829ColV, I-K94 cells after 10 min of exposure to 10 mM hydrogen peroxide at pH 3.0 |
 | |
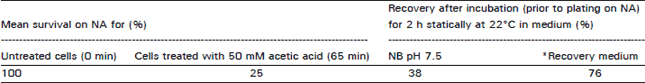
| Table 2: | Effect of incubation in different media on recovery of ED 1829ColV, I-K94 cells after 65 min of exposure to acetic acid at pH 4.5 |
 | |
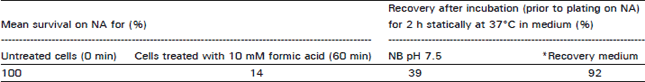
| Table 3: | Effect of incubation in different media on recovery of ED 1829ColV, I-K94 cells after 60 min of exposure to formic acid at pH 4.5 |
 | |
| Na: Nutrient agar, NB: Nutrient broth pH 7.5. *Recovery medium, NB: prepared in 0.1 M phosphate buffer of pH 7.5. Experiment was repeated thrice with consistent results | |
RESULTS
Recovery of non-habituated injured cells: The presence of phosphate in the recovery media, as explained in materials and methods, enhanced the recovery of the injured Ed1829CoIV, I-K94 cells which were treated with hydrogen peroxide and weak acids like acetic and formic acids.
The recovery of organisms, injured by exposure to hydrogen peroxide, was 4 time more in the recovery medium with phosphate then in medium without phosphate at room temperature (Table 1). Similarly the organism injured by acetic acid recovered significantly more, at-room temperature, in recovery media having phosphate than those of cells from medium without supplementation of phosphate (Table 2). A similar trend was exhibited by formic acid injured organisms (Table 3).
The results gave a clear indication that phosphate have some stimulatory effects on the recovery of stressed organisms by the fatty acids or hydrogen peroxide. This could be either due to repair of injured cells or both under the condition when the stress has been alleviated.
DISCUSSION
Recovery of injured cells: A stress may not be severe enough to destroy all bacteria present in a system may, instead, inflict sublethal injury to a proportion of the population. These injured cells can form colonies on low stress media but not on stressing media. However, if these innured organisms are removed from stressful environments they may revive so as to be able to from colonies on all media.
It has preciously been reported by Przybylski and Witter (1979), that the cells of Escherichia coli K-12, injured by sub-lethal acidification tend to recover in trypticase soy broth and in potassium phosphate buffer. Almost complete recovery (95%) was obtained afetr 120 min of incubation in 0.04 M potassium phosphate buffer of pH 8.0.
Musarrat and Ahmad (1988), observed that alkali-injured cells could recover on incubation in 0.05 M potassium phosphate buffer of pH 8.0, at 37°C, after 2 h. Recovery in phosphate buffer was not affected by metabolic inhibitors.
Recovery of injured ED1829CoIV, I-K94+ cells was observed, on incubation, in a complete medium like nutrient broth to a small extent; however, the process of recovery of the injured cells was somehow expedited in NB prepared in 0.1 M potassium phosphate buffer of pH 7.5 (Table 1, 2 and 3). The recovery medium used in the present study was totally supportive to growth and contained an energy source unlike the above mentioned previous two studies. It appears that phosphate buffer somehow supports the recovery of injured cells but its mechanisms is not clear. This could be either due to repair of injured cells or their growth at the expense of dead organisms or both under the condition when the stress has been alleviated which is in accordance with the findings of Dukan et al. (1997).
ACKNOWLEDGMENTS
The author acknowledges the facility and assistance provided by Professor Rowbury, R.J and David G. Smith, Department of Biology, CUL, London, U.K.
REFERENCES
- Dukan, S., Y. Levi and D. Touati, 1997. Recovery of culturability of an HOCl-stressed population of Escherichia coli after incubation in phosphate buffer: Resuscitation or regrowth? Applied Environ. Microbiol., 63: 4204-4209.
Direct Link - Freese, E., C.W. Sheu and E. Galliers, 1973. Function of lipophilic acids as antimicrobial food additives. Nature, 241: 321-325.
CrossRefDirect Link - Hicks, S.J. and R.J. Rowbury, 1986. Virulence plasmid-associated adhesion of Escherichia coli and its significance for chlorine resistance. J. Applied Bacteriol., 61: 209-218.
CrossRefDirect Link - Musarrat, J. and M. Ahmad, 1988. pH induced damage and repair in E. coli. Mutat. Res./DNA Repair Rep., 193: 219-227.
CrossRefDirect Link - Sinha, R.P., 1986. Toxicity of organic acids for repair-deficient strains of Escherichia coli. Applied Environ. Microbiol., 51: 1364-1366.
Direct Link - Przybylski, K.S. and L.D. Witter, 1979. Injury and recovery of Escherichia coli after sublethal acidification. Applied Environ. Microbiol., 37: 261-265.
Direct Link