
M. Hanif
School of Pure Applied Biology, University of Wales, Cardiff, UK.
M. S. Davies
School of Pure Applied Biology, University of Wales, Cardiff, UK.
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 4 | Page No.: 252-255







ABSTRACT
The seedlings of Triticum aestivum cvs. Lyallpur 73 and Lu-26-S were grown in aerated 0.1 Rorison nutrient solution for 72 hours. Then the kinetics of cells cycle in the root meristem were determined by a pulse labelling experiment with low specific activity tritiated thymidine following a 70 hours exposures to three concentrations of NaCl (0, 30, 70 mM NaCl). Treatments with 30 and 70 mM NaCl increased the length of cell cycle in cv Lu-26-S but that of Lyallpur 73 was decreased. The growth fraction (refers to percentage of meristematic cells which are actively cycling as opposed to these that are not cycling in the meristem) was increased in Lyallpur 73 from 7.9 to 11.11% over the range of 0-70 mM NaCl whereas that of Lu-26-S was decreased in response to increasing NaCl concentration.
PDF Abstract XML References Citation
How to cite this article
M. Hanif and M. S. Davies, 1998. The Influence of NaCl on the Cell Cycle and Growth Fraction in the Root Meristems of Triticum aestivum cvs. Lyallpur 73 and Lu-26-S. Pakistan Journal of Biological Sciences, 1: 252-255.
DOI: 10.3923/pjbs.1998.252.255
URL: https://scialert.net/abstract/?doi=pjbs.1998.252.255
DOI: 10.3923/pjbs.1998.252.255
URL: https://scialert.net/abstract/?doi=pjbs.1998.252.255
INTRODUCTION
A fundamental property of meristematic cells is their ability to grow and divide. The mitotic cell cycle is divided into a number of distinct phases. Nuclear DNA is replicated in the DNA synthetic (S) phase, preceded by a presynthetic interphase stage (G1) and followed by a postsynthetic interphase (G2) which, in turn, immediately precedes the onset of mitosis (M). Cells is G1 thus have the unreplicated (2C) DNA content and those ion G2 the 4C content. A standard way of measuring the cell cycle components in plant meristems is by the percentage labelled mitosis (PLM) method. Meristems are given a short exposure (pulse) with thymidine, radioactively labelled with tritium (3H-TdR) and then chased with cold (non-radioactive) TdR before being grown in the treatment solution. Roots are sampled at regular intervals thereafter for a period exceeding the likely duration of the cell cycle. The relationship between the percentage of cells in mitosis containing radioactive label and the time elapsed since labelling is determined. The method thus monitors the flow of a population of cells from S-phase through G2 and into and out of mitosis. Generally a double-peaked curve is found. The distance between peaks of labelled mitosis is taken to be equivalent to the duration of the cell cycle, the width of the first peak at half of the maximum height point is equal to the duration of S-phase plus the period of exposure to label; the interval from the start of labelling to 50% of the height of the ascending arm of the first peak is equal to the dyration of G2+1/2M; M is from where the initial limb rises to where it plateaus; G1 is obtained by difference (Quastler and Sherman, 1959). Scadeng and MacLeod (1976) proposed a modification of the method for calculating the duration of M from PLM data as below:
Where L.l is the mean labelling and M.l is mean mitotic index. Mean values for L.l and M.l are calculated over a period of time equal to the TC sratting from the start of labelling. The duration of G1 (TG1) is determined by the formula:
The cycle can be affected profoundly by environmental factors such as temperature, light and toxicity of metal (e.g., Van’t Hof and Ying, 1964; Francis and Barlow, 1988; Nabil 1996). Often some strains or varieties do better than other in coping with stress whether induced artificially or naturally. Predictably, the cell cycle lengthens in response to stress.
The aim of the work reported here were to determine the effects of varying NaCl concentrations in the nutrient solution on the length of cell cycle and its component phases and growth fraction in the root meristems of two cultivars of wheat.
MATERIALS AND METHODS
On experiment day 0 seeds of two Pakistani wheat cultivars Triticum aestivum (2n = 6x = 42) cvs. Lyallpur 73 and Lu-26-S were soon under the standard conditions (Hanif and Davies, 1998) in a growth room at a constant temperature of 20+0.5°C with illumination provided by white fluorescent tubes at a photon fluence rate of 153 (mol m–2 s–1) for 16 hours per day.Seed were imbibed for 24 hours in sandwich boxes containing 1 dm3 0.1 strength Rorison’s nutrient solution before sowing.
Ungerminated seeds were then removed and 15 seeds per cultivar were transferred to each of 6 rafts per sandwich box and were allowed to grow for a further 48 hours in Rorison’s nutrient solution. The rafts were then transferred into the various NaCl treatment solutions. The NaCl treatments for the these wheat cultivars were 0, 30 and 70 mM NaCl supplied in background of 0.1 strength Rorison’s nutrient solution. The different NaCl treatments were selected on sensitivity of roots of these cultivars to NaCl. There were two replicate sandwich boxes with six rafts per box per cultivar per NaCl concentration. After 70 hr. exposure to the NaCl treatments, the seedlings were transferred into an aqueous solution of radioactive thymidine 3H-TdR (concentration 37 KBq cm–3, specific activity 185 GBq m mol–1). After 1 hour the rafts were transferred to "cold" -non radioactive thymidine (concentration 10–5 M) for 1 hr. and then rinsed in deionised water before putting back into the original NaCl treatment. All the solutions were equilibrated to the growth room temperature before transfer. Five seedlings per cultivar per NaCl treatment were sampled at each time point. The first sample was taken 1 hr. after start of the pulse label with 3H-TdR and then at two hourly intervals thereafter, up to 36 hr. Samples were fixed in 3:1 v/v ethanol:glacial acetic acid and stored at 4°C. Five roots at each sampling time, for each cultivar per treatment were Feulgen stained and squash preparations were made of the apical root meristem as outlined previously (Hanif and Davies, 1998). Permanent autoradiographs were made at a room temperature of 18°C. Ilford K2 photogaraphic emulsion was mixed with an equal volume of distilled water, a drop of glycerol was added and the mixture was maintained at 40°C in a water bath. Slides from each treatment were dipped in the emulsion to drain off under condition of 40 percent relative humidity and at a room temperature of 25°C under llford F904 filters. The slides were stored in light-proof boxes in the presences of a desiccant (sillica gel) in the dark for 14 days at 4°C.
Autoradiographs were developed in llford Phenisol developer for 6 mins precisely, rinsed for approximately 30 seconds in distilled water and fixed in llford Hypam fixer at 20°C for 6 mins and were then raised in distilled water. The slides were then dehydrated in an ethanol series (30, 50, 70, 80 and 2x100% for 5 mins at room temperature). Slides were then cleared in xylene for 10 mins and mounted in DPX. On each of the developed autoradiographs, the percentage of mitoses which were labelled was determined. One thousand nuclei per slide were scored for these PLM determinations. The mitotic index (M.l) and labelling index were also measured on each slide. Labelling index (L.I) is the percentage of cells which had incorporated 3H-TdR expressed as a percentage of total cells scored. The M.I was calculated as the percentage of cells in mitosis expressed as a percentage of total cells scored. For each time point the relationship between the percentage of labelled mitoses and time elapsed since the start of pulse labelling (up to 37 hr.) was poltted and the duration of the cell cycle and the component phases calculated. For growth fraction the above programme was repeated but after 70 hr. exposure to NaCl treatments, the seedlings were transferred to the same treatment solution described above but containing 3H-TdR (concentration 9.25 KBq cm–3, specific activity 74 GBq m mol–1). The seedling were harvested at the end of 26 hr. continuous exposure to the 3H-TdR, rinsed with water and were then fixed in 3:1 (v/v) ethanol:glacial acetic acid. Permanent Feulgen stained autoradiographs were made as described above. Surprisingly, there was a complete absence of labelled cells on all slides. In case the absence of labelling was due to problem with the autoradiographs, further autoradiographs were made after removal of the photographic emulsion (Bianchi et al., 1964). However, again no apparent incorporation of label into any of the cells of the root meristerms occurred.
RESULTS AND DISCUSSION
The plots of percentage labelled mitoses against time elapsed since the start of labelling are shown in Figures 1-6. The duration of the cell cycles and its component phases are given in Table 1. The overall duration of the cell cycle in the meristems of roots of two cultivars in wheat, measured as the interval between the successive peaks of the PLM plots in the control (O NaCl) treatments were 14 hours in Lyallpur 73 and 12 h in Lu-26-S (Table 1). However, at 30 mM NaCl the cell cycle duration of Triticum aestivum cv. Lu-26-S was increased to 20 hr. but that of cv. Lyallpur 73 hr. was decreased to 12 hr. at 70 mM NaCl, the cell cycle of Lu-26-S was extended to 16 hr. but of Lyallpur 73 further decreased to 10 hr. (Table 1).
Despite these appreciable effects of NaCl on the length of the cell cycle in Lu-26-S, NaCl concentration had comparatively little effects on the duration of S-phase. Increases in cell cycle of Lu-26-S at 30 mM were mediated largely through an increase in the duration of M phase (4-fold)m G1 (3-fold) but decreases in G2 (4-fold). At 70 mM NaCl in the cultivar M phase was till 3 times that of the control and G1 was 1.7 times that of the control. S-phase was unchanged over the range of treatments; 30 mM and 70 mM NaCl in this cultivar. The data for Lyallpur 73, revealed two patterns. At 30 Mm NaCl, M phase showed 2.5-fold decrease compared with the control, G2 phase (3 fold-decrease) and S-phase a 3-fold decrease but G1 showed 2-fold increase. At 70 mM the further decrease in the cell cycle was mediated through a decrease in the duration of G1 (from 8.5 to 2.5 hr.) but M increased as did G2. Thus in this cultivar the duration of component phase showed no consistent trend with variation in To-for example as Tc shortened, G1 increased from 4.5 to 8.5, then declined to 2.5 hr., while M shortened and then increased. As there was no labelling of any cells of the cultivars investigated, perhaps a prolonged exposure of roots to a radioactive thymidine in continuous labelling experiments was more liable to perturb normal cell division activity than a short exposure used in pulse labelling experiments-although no firm evidence for this exists. However, the complete absence of labelled cells argues against a minor perturbation. It is more likely that, due to unknown factors, no cells were cycling in the root meristem of the cultivars. The growth fraction was calculated by dividing cell cycle duration by the respective cell doubling time estimates and expressing as a percentage value. The cell doubling times of the cultivars were very long and thus measures of growth fraction were very low and therefore must be treated with caution. Nevertheless, they can be used for the comparative purpose here. In cultivar Lyallpur 73 there was little difference between at 0 and 30 mM NaCl but an increase in growth fraction to 11 percent occurred a t 70 mM NaCl. In the cultivar Lu-26-S, the growth fraction decreased from 6.8 percent at 0 mM NaCl to 3.3 percent at 70 mM NaCl.
In the PLM method a double peaked curve is usually found; the distance between the peaks is measure of the duration of the cell cycle. In the wheat cultivars these PLM curves generally showed the initial major peak followed by a smaller peak.
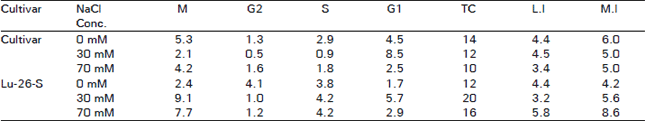
| Table 1: | Duration (h) of the cell cycle (TC) and its component phases estimated from the percentage labelled mitoses data in cells of root meristem of Triticum aestivum cvs. (Lyallpur 73 and LU-26-S) |
 | |
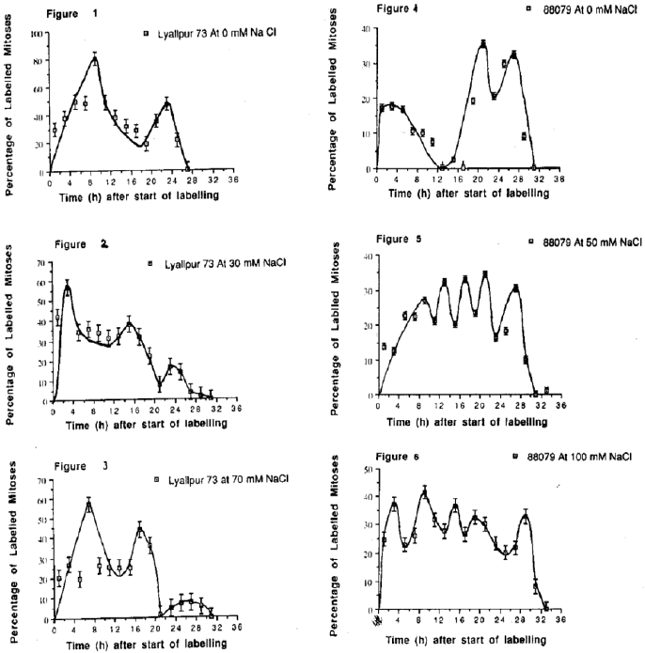 | |
| Fig. 1-6: | The relationship between the percentage labelled mitoses (SE±) and elapsed after the start of 1 hr. Pulse label with 3 H-thymidine, in squash preparations of root meristems of Tritium aestivum cvs. Lyallpur 73 and Lu-26-S following a 70 hr. Exposure to solutions containing 0, 30 and 70 mM NaCl supplied in a background of 0.1 strength Rorison’s nutrient solution. |
In PLM curves such as these not all points fall on a smooth curve and subjective judgment must be used in deciding the precise location of the curve. Although the peak to peak distance was generally unequivocal, some difficulty was encountered in deciding where precisely to draw the curve. Thus the peak to peak distance were accurate and reliable and thus so were the cell cycle duration estimates. The measure of the duration of the component phases showed no consistent trend with variation in Tc- for example as Tc shortened G1 increase from 4.5 to 8.5 hr. then decline to 2.5 hr. while M shortened then increased. In Lu-26-S showed a 1.7 fold increased (from 12 hr. to 20 hr.) over the range 0-30 mM NaCl but then decreased to 16 hr. at 70 mM. Again variation in Tc showed no consistent relationship with the duration of any particular phase, but both G1 and M varied in the same pattern as Tc. The effect of NaCl on cell cycle measure here was not consistent with its effect on root growth and mitotic index. Increasing NaCl concentration markedly reduced root growth and the mitotic index in these cultivars but shortened the cell cycle duration in Lu-26-S at 70 mM compared with that in the control. This finding was in contrast to other work of the effect of stress on cell division in roots. For example, the zinc induced reduction in root length in contrasting cultivars of F. rubra (Powell et al., 1986) was accompanied by corresponding increases in the duration of mitotic cycle in the root meristem, through an increase in the duration of G1. Thomas (1992) found that inhibition of root growth by Manganese was accompanied by corresponding increase in the duration of cell cycle mediated largely through as increase in G1. Creber et al. (1993) had also demonstrated that low temperature increases in cell cycle duration in Dactylis glomerata were mediated mainly through an increase in G1. It was possible that NaCl treatment reduced root growth, not primarily in these cultivars through an effect on the cell cycle but by reducing proportion of meristematic cells which were cycling (growth fraction).
ACKNOWLEDGMENTS
M. Hanif thanks the Government of Pakistan for the award of Central Overseas Training Scheme Scholarship for Ph.D. in U.K.
REFERENCES
- Bianchi, N., A. Lima-de-Faria and H. Jaworska, 1964. A technique for removing silver grains and gelatin from tritium autoradiographs of human chromosomes. Hereditas, 51: 207-211.
CrossRefDirect Link - Creber, H.M.C., M.S. Davies and D. Francis, 1993. Effects of temperature on cell division in root meristems of natural populations of Dactylis glomerata of contrasting latitudinal origins. Environ. Exp. Bot., 33: 433-442.
CrossRefDirect Link - Hanif, M. and M.S. Davies, 1998. Effects of NaCl on meristem size and proximity of root hairs to the root tips in Secale cereale (cv. K2) and Triticum aestivum (cv. Chinese spring). Pak. J. Biol. Sci., 1: 15-18.
CrossRefDirect Link - Powell, M.J., M.S. Davies and D. Francis, 1986. The influence of zinc on the cell cycle in the root meristem of a zinc-tolerant and a non-tolerant cultivar of Festuca rubra L. New Phytol., 102: 419-428.
CrossRefDirect Link - Quastler, H. and F.G. Sherman, 1959. Cell population kinetics in the intestinal epithelium of the mouse. Exp. Cell Res., 17: 420-438.
CrossRefDirect Link - Scadeng, D.W.F. and R.D. MacLeod, 1976. The effect of sucrose concentration on cell proliferation and quiescence in the apical meristem of excised roots of Pisum sativum L. Ann. Bot., 40: 947-955.
Direct Link - Van't Hof, J. and H.K. Ying, 1964. Relationship between the duration of the mitotic cycle, the rate of cell production and the rate of growth of Pisum roots at different temperatures. Cytologia, 29: 399-406.
CrossRefDirect Link