
Mohammad Adil
Department of Pharmacology, Poona College of Pharmacy, Bharati Vidyapeeth Deemed University, Erandwane, Paud Road, Pune, 411 038, Maharashtra, India
Asjad Visnagri
Department of Pharmacology, Poona College of Pharmacy, Bharati Vidyapeeth Deemed University, Erandwane, Paud Road, Pune, 411 038, Maharashtra, India
V. Shiva Kumar
Department of Pharmacology, Poona College of Pharmacy, Bharati Vidyapeeth Deemed University, Erandwane, Paud Road, Pune, 411 038, Maharashtra, India
Amit D. Kandhare
Department of Pharmacology, Poona College of Pharmacy, Bharati Vidyapeeth Deemed University, Erandwane, Paud Road, Pune, 411 038, Maharashtra, India
Pinaki Ghosh
Department of Pharmacology, Poona College of Pharmacy, Bharati Vidyapeeth Deemed University, Erandwane, Paud Road, Pune, 411 038, Maharashtra, India
Subhash L. Bodhankar
Department of Pharmacology, Poona College of Pharmacy, Bharati Vidyapeeth Deemed University, Erandwane, Paud Road, Pune, 411 038, Maharashtra, India
Pharmacologia
Year: 2014 | Volume: 5 | Issue: 6 | Page No.: 222-234







ABSTRACT
Background: Arsenicals induced testicular cancer is a serious debilitating epidemic affecting all social strata in developing as well as developed countries. Naringin possess antioxidant and free radical scavenging properties. Hence, the present investigation was design to elucidate the effect of naringin against sodium arsenite induced testicular toxicity in laboratory rats. Materials and Methods: Male Sprague-Dawley rats were pretreated with either vehicle (control), naringin (20, 40 and 80 mg kg-1, p.o.) and Co-enzyme Q10 (10 mg kg-1, p.o) for 7 days. Testicular toxicity was induced by sodium arsenite (10 mg kg-1, p. o., for two consecutive days). Drug treatment was continued up to 15th day. On 16th day reproductive organo-somatic indices, biochemical and histolopathogical determinations were carried out. Results: Rats pretreated with naringin (40 and 80 mg kg-1, p.o.) showed significant and dose dependant increased (p<0.01 and 0.001, respectively) in relative organ weight (epididymis, seminal vesicle and testis) and sperm count. It also showed significant increased (p<0.01 and 0.001, respectively) in level of serum protein, albumin, AST, ALT and activity of testicular endogenous enzymes (nitric oxide (NO) content superoxide dismutase (SOD) and reduced glutathione (GSH). The elevated levels of serum creatinine and glucose, testicular MDA and NO were significantly inhibited (p<0.01 and 0.001, respectively) by the naringin (40 and 80 mg kg-1, p.o.). It significantly (p<0.01) restored the alteration in serum lipid profile. Elevated levels of pro-inflammatory cytokines (Tumor necrosis factor-alpha (TNF-α) and Interleukin-1beta (IL-1β) were significantly and dose dependently (p<0.01 and 0.001, respectively) inhibited by the naringin (40 and 80 mg kg-1, p.o.) treatment. It also restored the histological aberration induced by the sodium arsenite. Conclusion: Naringin exerted its protective effects against sodium arsenite induced testicular toxicity via modulation of endogenous antioxidant enzyme, inhibition of release of oxido-inflammatory mediators such as MDA and NO as well as pro-inflammatory cytokines.
PDF Abstract XML References Citation
How to cite this article
Mohammad Adil, Asjad Visnagri, V. Shiva Kumar, Amit D. Kandhare, Pinaki Ghosh and Subhash L. Bodhankar, 2014. Protective Effect of Naringin on Sodium Arsenite Induced Testicular Toxicity via Modulation of Biochemical Perturbations in Experimental Rats. Pharmacologia, 5: 222-234.
URL: https://scialert.net/abstract/?doi=pharmacologia.2014.222.234
URL: https://scialert.net/abstract/?doi=pharmacologia.2014.222.234
INTRODUCTION
Arsenic is an inorganic compound which exists in environment in abundant amount and it has ability to cause multiple causative effects in animals as well as in human (Bates et al., 1992; Liu et al., 2001). There are various natural and man-made sources through which arsenic release in the environment like agricultural pesticides industries, coal fired power plants, glassware manufacturing, wood preservatives and smelting industries, cosmetic industries, etc. (Tchounwou et al., 2002). Water-soluble trivalent arsenite form (As III) and pentavalent arsenate form (As V) are the two most common forms of arsenic, however inorganic trivalent arsenic has more potential to cause toxicity than pentavalent arsenic (Bertolero et al., 1987). Water, food and drugs are the common ways through which exposure of arsenic easily occur and this arsenic is quickly absorbed through gastrointestinal tract results in the toxicity of multiple organs including liver, lung, testis, spleen, kidney, etc. (Pant et al., 2004; Sarkar et al., 2003). This toxic effect is mediated through an array of vicious processes including obstacles in DNA repair, changes in ubiquitous enzymatic activities, alteration in gene expression and involvement of oxidative stress (Das et al., 2009; Yousef et al., 2008).
Impaired male reproductive function is a result of arsenic intoxication and reflected by suppression of spermatogenesis and androgenesis, reduction of the weight of testes, imbalance of sex hormones and accessory sex organs (Pant et al., 2004; Sarkar et al., 2003). Exact mechanism behind the arsenic induced testicular toxicity is not fully elucidated however, studies carried out in various animal models of testicular toxicity revealed the mechanism behind arsenic induced toxicity (Kandhare et al., 2011b). Various finding favours the role of oxidative stress and inflammation with the increment of generation of reactive oxygen species and production of pro-inflammatory cytokines (Chang et al., 2007; Ghosh et al., 2012a, b, c; Singh et al., 2010).
It has been documented that diseases which are related to oxidative stress can be easily treated by applying nutritional antioxidants (Kamble et al., 2013; Kandhare et al., 2011a, 2013a). Isolated bioactive moieties from plant are known to exert their protective effects by scavenging free radicals and modulating antioxidant defense system (Kandhare et al., 2012e; Raygude et al., 2012a; Visnagri et al., 2014). Naringin (4’,5,7-trihydroxy flavonone 7-rhamnoglucoside), a plant bioflavonoid is naturally present in grape fruits and other citrus species with the ability of positive effect on human health (Jagetia and Reddy, 2002; Jagetia et al., 2003; Kandhare et al., 2012b). Naringin has free radical scavenging property that attenuate induced molecular menac such as reduction of cellular and chromosomal aberrations (Jagetia et al., 2003). Various pharmacological actions like of naringin antiulcer, antidiabetic, renoprotective, cardioprotective, anti-inflammatory, anticancer properties ( Ali et al., 2004; Benavente-Garcia and Castillo, 2008; Choe et al., 2001; Gonzalez-Gallego et al., 2007; Martin et al., 1994; Punithavathi et al., 2008; Rajadurai and Prince, 2007; Singh et al., 2004) have been reported. Naringin caused inhibition of lipopolysacharides which induced squeezing of tumour necrosis factor-á in macrophages of mice (Kawaguchi et al., 1999). Naringin possess preventive effect on lipoproteins, lipid as well as enzymes related with metabolism of lipid and lipid peroxides in isoprenaline-induced myocardial infarction in rats (Rajadurai and Prince, 2007). Kandhare et al. (2012a) reported that naringin possess neuroprotective effects in streptozotocin (STZ) induced neuropathic pain (Kandhare et al., 2012b). Moreover, naringin ameliorates acetic acid induced colitis through modulation of endogenous oxido-nitrosative balance and DNA damage (Kumar et al., 2013). Various plant bioflavonoids including curcumin, rutin and hesperidin have been shown to play ameliorating effect in sodium arsenite induced biochemical perturbations in testis of rats (Akondi et al., 2011; Taepongsorat et al., 2008; Yousef et al., 2008). Previous report of Akondi et al. (2011) reported that naringin exert its protective effect on testicular tissue via inhibition of oxidative stress in type I diabetes mellitus (Akondi et al., 2011). On the basis of above outcomes, the objective of present study was to study the effect of naringin in sodium arsenite induced testicular toxicity by assessing various biochemical, molecular and histological parameters.
MATERIALS AND METHODS
Animals: Adult male Sprague-Dawley rats (200-250 g) were purchased from the National Institute of Biosciences, Pune (India). They were housed at 24±1°C, with relative humidity of 45–55% and12:12 h dark/light cycle. The animals had free access to standard pellet chow (Pranav Agro industries Ltd., Sangli, India) and water throughout the experimental protocol. All experiments were carried out between 09:00 and 17:00 h. The experimental protocol was approved by the Institutional Animal Ethics Committee (IAEC) of Poona College of Pharmacy, Pune (CPCSEA/83/2012) and performed in accordance with the guidelines of Committee for Control and Supervision of Experimentation on Animals (CPCSEA), Government of India on animal experimentation.
Drugs and chemicals: Naringin was purchased from Sigma Chemical Co. (St Louis, MO, USA). Co-enzyme Q10 was supplied by Medicines Pvt. Ltd., (Mumbai, India) and sodium arsenite powder (Otto Chemicals, India). 1,1’,3,3’-Tetraethoxypropane, crystalline beef liver catalase, reduced glutathione (GSH), 5,5’-dithiobis (2-nitrobenzoic acid) were purchased from S.D. Fine Chemicals, Mumbai, India. Sulphanilamides, naphthalamine diamine Hcl and phosphoric acid were obtained from LobaChemie Pvt. Ltd., Mumbai, India. Rat Tumor necrosis factor-alpha (TNF-α) and Interleukin-1beta (IL-1β) ELISA kits were purchased from Thermo Scientific (Rockford, IL Campus, USA).
Experimental design: Rats were randomly divided into following groups of 6 animals each as follows:
| Group I: | Normal [N]: Received double distilled water (10 mg kg-1, p.o. for 15 days) |
| Group II: | Arsenic Control [As]: Received sodium arsenite (10 mg kg-1, p.o. for two consecutive days) and treated with distilled water (10 g kg-1, p.o. for 15 days) |
| Group III: | Arsenic+Naringin (20 mg kg-1) [As+N (20)]: Received sodium arsenite (10 mg kg-1, p.o. for two consecutive days) and treated with naringin (20 mg kg-1, p.o. for 15 days) |
| Group IV: | Arsenic+Naringin (40 mg kg-1) [As+N (40)]: Received sodium arsenite (10 mg kg-1, p.o. for two consecutive days) and treated with naringin (40 mg kg-1, p.o. for 15 days) |
| Group V: | Arsenic+Naringin (80 mg kg-1) [As+N (80)]: Received sodium arsenite (10 mg kg-1, p.o. for two consecutive days) and treated with naringin (80 mg kg-1, p.o. for 15 days) |
| Group VI: | Arsenic+Co-enzyme Q10 (10 mg kg-1) [As+CoQ10 (10)]: Received sodium arsenite (10 mg kg-1, p.o. for two consecutive days) and treated with Co-enzyme Q10 (10 mg kg-1, p.o. for 15 days) |
Testicular damage was induced in rats of the second and third groups by sodium arsenite at a dose of 10 mg kg-1, orally for two consecutive days (Das et al., 2009). Co-enzyme Q10 was prepared as aqueous solution with addition of Tween 80 (1%), while sodium arsenite was dissolved in normal saline. The three different dosage of naringin i.e., 20, 40 and 80 mg kg-1 were selected on the basis of previous study (Kandhare et al., 2012b). Everyday naringin solution was freshly prepared in distilled water. An appropriate stock solution was prepared to administer selected dosage of 20, 40 and 80 mg kg-1 orally for 15 days. Naringin and Co-enzyme Q10 were administered for 7 consecutive days before arsenite administration and continued up to 15 days. On 16th day rats were anesthetized by anesthetic ether and blood was withdrawn by retro orbital puncture. Each blood sample was collected into separate vials for determination of serum parameters. After blood collection the animals were sacrificed by cervical dislocation and testis were rapidly removed and stored at -80°C for biochemical parameters.
Serum biochemistry: The serum was separated by centrifugation using Eppendorf Cryocentrifuge (model No. 5810, Germany), maintained at 4°C and run at speed of 7000 rpm for 15 min. Serum protein, albumin, creatinine, Aspartate Aminotransferase (AST), Alanine transaminase (ALT), glucose, cholesterol, HDL (High-Density Lipoprotein), LDL (Low-Density Lipoprotein) and triglyceride were measured by spectrophotometer (UV-Visible spectrophotometer, Jasco V-530, Japan) using reagent kits according to procedure provided by manufacturer (Accurex Biomedical Pvt. Ltd., Mumbai, India).
Biochemical estimation of testis homogenate
Tissue homogenate preparation: Animals were sacrificed at the end of study and testis were immediately isolated. Tissue homogenate was prepared with 0.1 M tris-HCl buffer (pH 7.4) and supernatant of homogenates was employed to estimate superoxide dismutase (SOD), reduced glutathione (GSH), lipid peroxidation (MDA content) and nitric oxide (NO content) as described previously (Kandhare et al., 2012a, b, c; Visnagri et al., 2012).
Quantification of testicular TNF-α and IL-1β: The quantification of testicular TNF-α and IL-1β was performed in testis homogenate with the help and instructions provided by rat TNF-α and IL-1β ELISA kit manufacturer (Thermo Scientific, Rockford, IL Campus, USA).
Histopathological analysis of testis: Specimen of other testis was stored in 10% formalin for 24 h. The specimen was dehydrated and placed in xylene for 1 h (3 times) and later in ethyl alcohol (70, 90 and 100%) for 2 h, respectively. The infiltration and impregnation was carried out by treating with paraffin wax twice, each time for one h. Tissue specimens were cut into sections of 3-5 μm thickness and stained with hematoxylin and eosin (H and E). The sections were mounted on slide by use of Distrene Pthalate Xylene (DPX) as mounting medium. Sections were examined under a light microscope to inspect the histopathology features of specimen and infiltration of cells. The various changes in histological features were graded as Grade 0 (not present or mild) l; Grade 1 (mild); Grade 2 (moderate); and Grade 3 (severe).
Statistical analysis: Data were expressed as mean±standard error of mean (SEM). Data analysis was performed using Graph Pad Prism 5.0 software (Graph Pad, San Diego, CA, USA). Data of biochemical parameters were analyzed using one way analysis of variance (ANOVA) and Dunnett’s test was applied for post hoc analysis. A value of p<0.05 was considered to be statistically significant.
RESULTS
Effect of naringin and Co-enzyme Q10 on body weight and relative organ weights in male rats: There was significant (p<0.001) reduction in body weight, relative weight of epididymis, seminal vesicle and testis of arsenic control group as compared to normal group. On the other hand, there was significant (p<0.01 and 0.001) and dose dependant (40 and 80 mg kg-1) increase in the body weight, relative weight of epididymis, seminal vesicle and testis in naringin treated group as compared to arsenic control group. In co-enzyme Q10 (10 mg kg-1) treated group significant (p<0.001) increase in the body weight, relative weights of epididymis, seminal vesicle and testis as compared to arsenic control group was observed (Table 1).
Effect of naringin and Co-enzyme Q10 on sperm count in male rats: Sperm count in arsenic control rats was significantly decreased (p<0.001) after arsenite administration as compared to normal rats. Sperm count in naringin treated rats was significantly (p<0.01 and 0.001, respectively) and dose dependently (40 and 80 mg kg-1) increased as compared to arsenic control rats.
 |
 |
The reduced sperm count after arsenite administration was significantly (p<0.001) increased in Co-enzyme Q10 (10 mg kg-1) treated rats as compared to arsenic control rats (Table 1).
Effect of naringin and Co-enzyme Q10 on serum protein, albumin, AST and ALT in male rats: There was significant (p<0.001) decrease in serum protein, albumin, AST and ALT level in arsenic control animals as compared to normal animals. Treatment with naringin significantly (p<0.01 and 0.001) and dose dependently (40 and 80 mg kg-1) increased the serum protein, albumin, AST and ALT level as compared to arsenic control animals. Treatment with Co-enzyme Q10 (10 mg kg-1) significantly (p<0.001) increased the serum protein, albumin, AST and ALT level as compared to arsenic control group (Table 2).
Effect of naringin and Co-enzyme Q10 on serum creatinine and glucose level in male rats: There was significant (p<0.001) increase in serum creatinine and glucose level in arsenic control group as compared to normal group. Treatment of naringin significantly (p<0.05 and 0.001, respectively) and dose dependently (40 and 80 mg kg-1) decreased serum creatinine and serum glucose levels as compared to arsenic control animals. Co-enzyme Q10 (10 mg kg-1) treated rats showed significant (p<0.001) reduction in serum creatinine and serum glucose levels as compared to arsenic control group (Table 2).
Effect of naringin and Co-enzyme Q10 on lipid profile in male rats: There was significant (p<0.001) increase in serum cholesterol, triglyceride and LDL-C level in arsenic control group as compared to normal group. The increased level of cholesterol and LDL-C due to arsenic administration was significantly (p<0.05 and 0.01) and dose decrease (40 and 80 mg kg-1) by naringin treatment as compared to arsenic control group. There was significantly (p<0.01 and 0.001) and dose dependant (40 and 80 mg kg-1) reduction in the serum triglyceride level by naringin treatment as compared to arsenic control animals. Treatment with Co-enzyme Q10 (10 mg kg-1) significantly decreased the level serum cholesterol (p<0.001), triglyceride (p<0.001) and LDL-C (p<0.01) as compared to arsenic control group (Table 3).
The level of serum HDL-C in arsenic control rats was significantly (p<0.001) decreased after arsenite administration as compared to normal rats. Serum HDL-C level in naringin (80 mg kg-1) treated rats was significantly (p<0.01) increased as compared to arsenic control rats. When compared with arsenic control rats, naringin (20 and 40 mg kg-1) treated rats failed to produce any significant change in serum HDL-C. The reduced serum HDL-C level was significantly (p<0.05) increased by Co-enzyme Q10 (10 mg kg-1) treated rats as compared to arsenic control rats (Table 3).
Effect of naringin and Co-enzyme Q10 on testicular SOD and GSH in male rats: Testicular SOD and GSH levels of arsenic control rats were significantly (p<0.001) decreased after administration of sodium arsenite as compared to normal rats.
 |
 |
The testicular SOD level in naringin treated rats was significantly (p<0.05 and 0.001) and dose dependently (40 and 80 mg kg-1) increased as compared to arsenic control rats. Whereas naringin treatment showed significant (p<0.01 and 0.001) and dose dependant inhibition (40 and 80 mg kg-1) in the arsenite induced reduction in testicular GSH level when compared with arsenic control rats. When compared with arsenic control rats, rats treated with co-enzyme Q10 (10 mg kg-1) showed significant (p<0.001) increase in testicular SOD and GSH level (Table 4).
Effect of naringin and Co-enzyme Q10 on testicular MDA and NO level in male rats: Lipid peroxidation (MDA) and NO levels were significantly increased (p<0.001) in arsenic control rats after administration of sodium arsenite as compared to normal rats. The testicular MDA and NO levels in naringin treated rats were decreased significantly (p<0.01 and 0.001) and dose dependently (40 and 80 mg kg-1) as compared to arsenic control rats. When compared with arsenic control rats, Co-enzyme Q10 (10 mg kg-1) treated rats also showed significant (p<0.001) inhibition in this elevated level of testicular MDA and NO (Table 4).
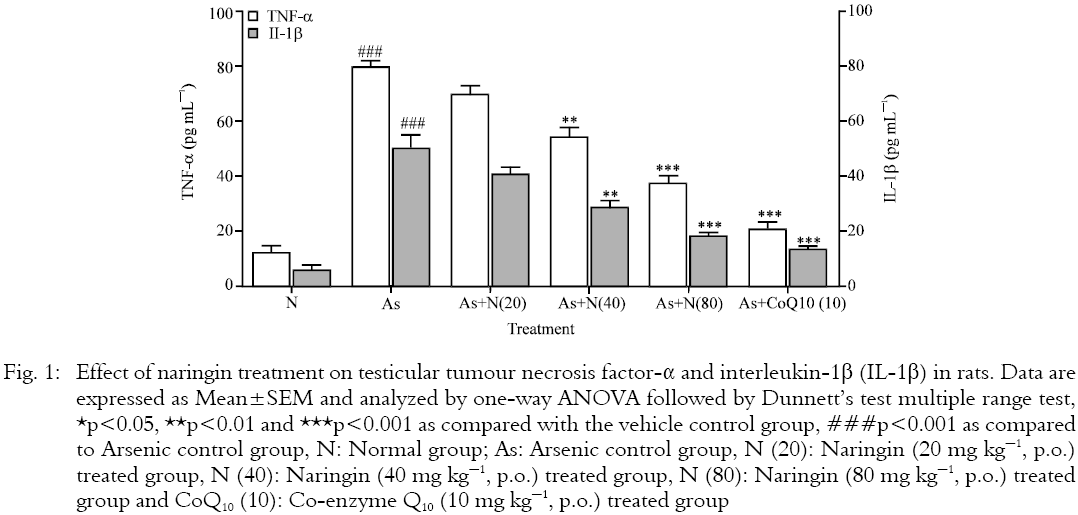
Effect of naringin on and Co-enzyme Q10 testicular TNF-α and IL-1β level in male rats: There were significant (p<0.001) increase in the testicular TNF-α and IL-1β levels in arsenic control rats as compared with normal rats. These elevated testicular TNF-α and IL-1β levels were significantly (p<0.01 and p<0.001) and dose-dependently (40 and 80 mg kg-1) inhibited by naringin treatment when compared with arsenic control rats. Treatment with Co-enzyme Q10 (10 mg kg-1) showed significant (p<0.001) reduction in testicular TNF-α and IL-1β levels as compared to arsenic control group (Fig. 1).
Effect of naringin and Co-enzyme Q10 on histopathological analysis of testis in male rats: Histopathological investigation of testis from normal rat showed well structured features including the appearance of normal spermatogenesis and spermiogenesis cycles, presence of uniformly arranged longitudinal to transversely placed seminiferous tubules with minimal interstitial spaces (Fig. 2a). It showed normal leydig cells with presence of few congested blood vessels (Grade 1). However, sections of testis from arsenic control animals showed ruptured follicles and few follicles showing reduction in the number of spermatozoa and degenerated inter follicular septa (Fig. 2b). It also exhibits the prominent features like necrosis in seminiferous tubules (Grade 4), interstitium edema (Grade 3), maturation arrest, failure in spermiation and vacuolization of spermatid nuclei (Grade 4). Histopathological investigation of testis tissue from Co-enzyme Q10 (10 mg kg-1) treated rat showed normal spermatogenesis characterized by the presence of mature secondary spermatids at the lumenal edges of epithelia but presence of moderate interstitium edema (Grade 1) with degenerative changes (Grade 1) in histological findings of testis (Fig. 2c). Naringin (80 mg kg-1) treatment improved testicular aberration induced after arsenic toxicity. It showed the restoration of normal morphological structures of seminiferous tubules but presence of degenerated (Grade 1) and reduced number of Leydig cells.
 |
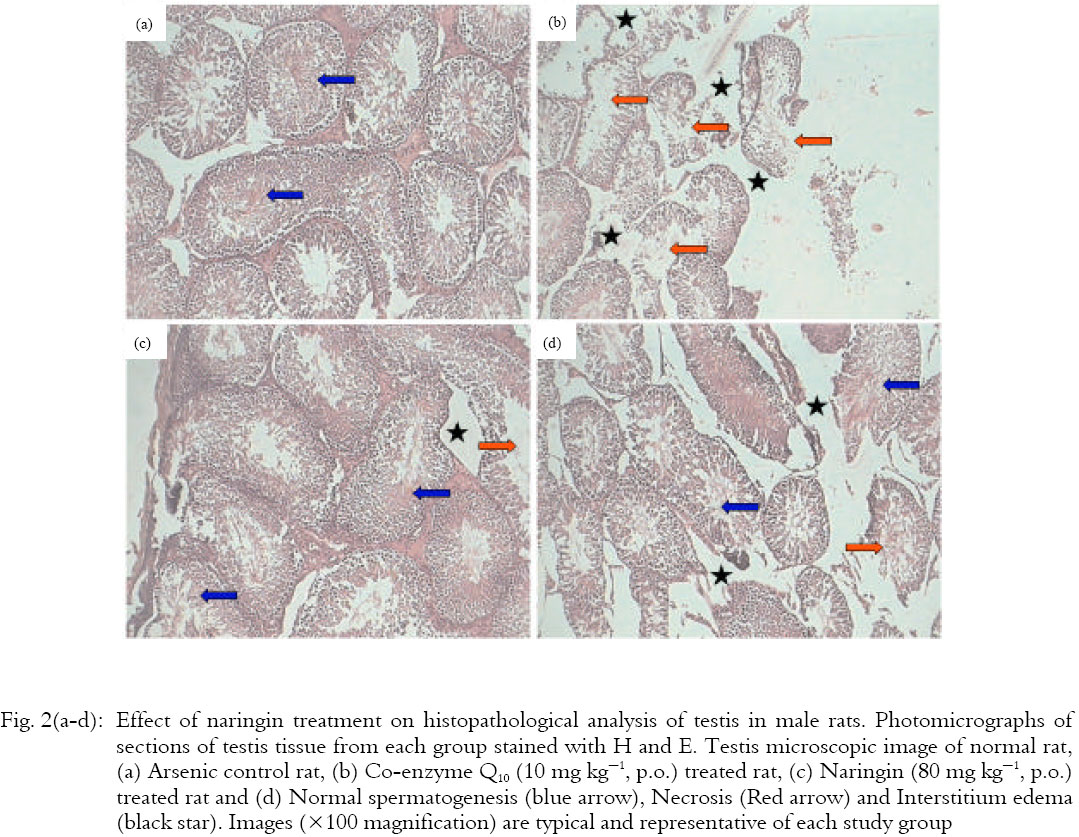 |
The numbers of spermatids were increased, tubular basement membrane thickness was reduced and the intertubular space was decreased (Fig. 2d). Restoration of active spermatogenesis in most of seminiferous tubules w ere seen in naringin (80 mg kg-1) treated animals. The interstitial tissue edema (Grade 2) and necrosis (Grade 1) were decreased and histology showed minimal injury of testicular tissue.
DISCUSSION
Arsenic has wide range of application in manufacturing industries in an array of domain across insecticides, preservatives and dyes. Products from the industries are used by all strata of society and an imperative possibility of arsenic mediated pathobiology lurks in the background. The indispensible use of their products in the daily life promulgates the needs for agents possessing adequate property of quenching the toxic aberrations precipitative due to its chemical properties. It is present ubiquitously in the environment but in minuscule proportion (Ramanathan et al., 2003). IARC (International agency for research and cancer) has christened arsenic as group-I carcinogen reflecting the impact levied by arsenic toxicity on global health (IARC, 2004). However, well directed research initiatives have not produced or identified molecules or targets that could be used as cynosure of international efforts to alleviated arsenic induced toxicity.
Exposure to high levels of arsenic are liable to develop lung, testis, skin, bladder cancer and occlusive vascular disease thus withered populations are at elevated risk for these conditions (Bates et al., 1992; Smith et al., 1992). Arsenic is metabolized by recurring reduction and oxidative methylation. Various arsenic metabolic intermediates such as monomethylarsonous acid and dimethylarsinous acid were also found in urine, which are highly toxic than inorganic arsenicals (Kobayashi et al., 2005). Arsenic exposure leads to commencement of oxidative stress, increased lipid peroxidation, depletion of antioxidant defences and increased production of pro-inflammatory cytokines (Fouad et al., 2011). There are various facets of arsenic induced toxicity in compassing oxidative, nitrostative and molecular perturbation due to mono, di and tri methylated forms which have enhanced toxic potential as compared to the parent molecules.
The presence investigation was undertaken to evaluate the effect of naringin to reverse arsenic induced toxicity in laboratory rats. The literature is punctuated with evidences proving antioxidants Reactive Oxygen Species (ROS) scavenging properties of naringin as the principle components of arsenic toxicity is oxido-nitrostative stress leading to DNA damage hence the selection of naringin in this animal model is justifiable (Kandhare et al., 2012b, c; Kumar et al., 2013). Oxidative stress has a plethora of components which comprise various enzymes involved in the electron transport chain (Kandhare et al., 2013b, c).
The administration of sodium arsenite leads to decrease in the absolute weight of the testis, epididymis and seminal vesicle which indicate the toxic effects of arsenite on these reproductive organs, this observation is in agreement with a earlier finding (Morakinyo et al., 2010). Sperm count and sperm morphology are key features of male fertility, as these are the principal markers in testicular spermatogenesis and epididymal maturation (Sarkar et al., 2003). Moreover, undesirable effect of arsenite on sperm count may be accountable due to low testosterone concentration and it has been reported that high level of testosterone in testis is crucially required for usual spermatogenesis, maintenance of sperm morphology and normal composition of seminiferous tubules (Sharpe et al., 1988, 1992). In present investigation, the dwindling of sperm count by sodium arsenite treatment is corroborated with the findings of others (Jana et al., 2006; Sarkar et al., 2003). Treatment with naringin significantly inhibited alteration in absolute weight of testis, epididymis and seminal vesicle and improved the sperm count. The increase in sperm count is also evident by the restoration of active spermatogenesis in seminiferous tubules observed in histopathological investigation of testis of naringin treated animals. The results of present investigation are in line with the finding of previous researcher (Akondi et al., 2011).
The demur in plasma total protein due to induction with sodium arsenite was essentially due to diminished albumin fraction. The inhibitory effect of sodium arsenite on total protein is in accordance with the finding of previous researchers (Kaltreider et al., 2001; Nandi et al., 2005) reported that arsenic inhibited the aptitude of glucocorticoid and its receptor to turn on genes, which plays an essential role in protein metabolism. The reduced level of plasma protein in rodents due to environmental pollutants could be endorsed to changes in protein as well as free amino acid metabolism and their synthesis in the liver. Treatment of naringin showed prominent effect by increasing the plasma total protein and albumin levels. In addition, the decrease in concentration of serum protein may be due to excessive loss of proteins, which leads to decrease in plasma proteins. Sodium arsenite caused reduction in the activities of serum AST, ALT levels (Yousef et al., 2008). However, treatment of naringin significantly restored the AST and ALT levels in serum.
The imbalance in glucose production and utilization leads to hyperglycaemia. The present study reveals the increase in glucose levels of sodium arsenite administered rats. It has been reported that methylated metabolites of arsenic are responsible for induction of diabetes mellitus (Tseng, 2004). Moreover, Oxidative Stress (OS) has been considered as a major link to insulin resistance and β-cell dysfunction which thwart the release of insulin in to circulation along with damage of insulin secreting β cells after prolonged exposure to arsenic (Hayden, 2002). Treatment of naringin significantly reduced the blood glucose level which is in accordance with the previous study (Kandhare et al., 2012b). The present study indicated that administration of sodium arsenite caused noteworthy increase in serum creatinine, which is due to decrease in the clearance from urine and leads to minimize an ability of the kidneys to filter waste products from the blood and excrete them in the urine naringin treatment show significant decrease in serum creatinine.
The present data showed that plasma cholesterol, triglyceride and LDL were significantly increased by sodium arsenite administration, while HDL levels were decreased, which are in accordance with (Mann et al., 2005) who reported that liver X receptor (LXR) lead to hypercholesterolemia. It is also reported that lipid and carbohydrate metabolism are primarily regulated by LXR genes (Steffensen and Gustafsson, 2004). Oral treatment of naringin promisingly reduced the serum cholesterol and triglyceride levels and increased HDL levels. The finding of present investigation is in accordance with the results of previous researcher where oral administration of naringin restored the altered level of serum lipid in paracetamol induced hepatotoxicity (Javed et al., 2013).
ROS generation leads to damage of macromolecules and DNA (Raygude et al., 2012b). In a typical organ system, the overall metabolism of formation of ROS involves damage to the physiological quenching of the superoxide radical anion (O2¯). Superoxide anion is converted to H2O via glutathione peroxidase or catalase (Patil et al., 2012c; Visnagri et al., 2013). Another pathway for diminution of superoxide free radical anion is via lipid conjugation (Kandhare et al., 2012d). This pathway involves β-oxidation by lipid peroxides leading to formation of malonaldehyde and 4-hydroxynoneol. This physiological balance is disregulated under exogenous stress like arsenic leading to elevation of MDA and reduction of GSH and SOD.
MDA is a reproducible index of oxidative stress in biological system produced due to lipid peroxidation (Gosavi et al., 2012a; Patil et al., 2012b). This yard stick has been reported to mirror loss of testicular function and sperm damage owing to oxidative damage mediated by arsenic (Morakinyo et al., 2010; Sikka, 2004). Glutathion is a pivotal components of cell growth differentiation and protection comprising cellular thiol redox buffer which mediates and maintains thiol/disulfide redox balance (Sies, 1999). Severe oxidative stress corresponds to decrease in glutathione levels. Glutathione has major role in suppression and scavenging of ROS (Willcox et al., 2004). Arsenic has been a proven agent disturbing the pro-oxidant -antioxidant balance owing to its intrinsic affinity for sulfahydril groups of GSH (Yamanaka et al., 1991). GSH exerts protective activity against toxic infiltrators viz., arsenic metabolism by donating electrons to convert arsenic5+ to arsenic3+ thus GSH depletion due to arsenic is a hallmark of arsenic induced cellular damage (Ramanathan et al., 2002). Naringin elucidate a discernable up-regulation of GSH and down-regulation of MDA conforming its ability to reverse arsenic induced oxidative damage.
SOD accentuate the dismutation of superoxide to H2O2 which is regulated by catalase (Usoh et al., 2005). SOD acts as primary line of defence to free radical induced aberrations. The reduction in levels of SOD corresponds to reduction of cellular defensive mechanism (Kandhare et al., 2012a). In present investigation, naringin elevated the levels of SOD in a dose dependent manner demonstrating its ability to restore cellular oxido-reductive balance.
It has been demonstrated that administration of sodium arsenite leads to elevate nitric oxide levels which produce cytotoxicity and oxidative damage (Bera et al., 2010; Flora et al., 2009). Superfluous of nitric oxide interact with superoxide anion to produce peroxynitrite radical that causes additional cell damage by nitrating and oxidizing cellular macromolecules. NO has dual role in cellular stress management mechanism (Gosavi et al., 2012b; Patil et al., 2012a). There are three isoforms of nitric oxide synthase (NOS) viz., eNOS (endothelial NOS), nNOS(neuronal NOS) and iNOS (inducible NOS). iNOS play a major pathological role in mediating inflammation host defence mechanism and maintenance of nitrosative balance in the cellular environments (Patil et al., 2012d). NO is produced by iNOS which in turn blind with O2¯ leading to formation of ONOO¯ (Beckman and Crow, 1993). This radical is termed as peroxinitrites which damages intra-mitochondrial and cellular oxidative balance (Radi, 2004). In the present investigation, naringin reduced the elevated nitric oxide levels thus evidencing its potential to restore disregulated nitrosative balance.
It has been reported that ROS are produced by cytokines. The production of hydroxyl radicals and lipid peroxidation is increased in tumorigenic fibroblast cells and mesangal cells by TNF-α (Bohler et al., 2000). Inorganic form of arsenic is considered to trigger inflammatory response. Exposure to arsenite in rats have been found to increase serum levels of TNF-α (Mukherjee et al., 2006). Over-expression of TNF-α was reported in humans chronically exposed to inorganic arsenite. TNF-α production is remarkably increased by cytotoxic effects of arsenite in murine micophages (Sakurai et al., 1998). Due to arsenic induced pro-inflammatory cytokines there may be increased level of oxidative stress which leads to produce toxic effect of arsenite in present study. However, treatment of naringin prominently reduced the TNF-α level which is in line with the study of previous findings (Kandhare et al., 2012b). IL-1β is a member of pleiotropic family, utterly formed by macrophages (Bergh and Soder, 1990; Cannon et al., 1990) and influences Leydig cells in a paracrine manner. Additionally, testosterone secretion was noteworthy increased by the administration of IL-1β into rat testis (Gerendai et al., 2005). Moreover, inhibition of Leydig cell steroidogenesis during inflammation is reported by IL-1β (Calkins et al., 1988). Aforementioned mechanism of IL-1β may be responsible in present investigation to produce testicular toxicity in sodium arsenite treated rats which was significantly restored by the naringin treatment.
Co-enzyme Q10 (ubiquinone or 2, 3 dimethoxy-5 methyl-6-decaprenyl benzoquinone) is a liquid-soluble potent antioxidant and anti-inflammatory substance. It has been previously reported that treatment with Co-enzyme Q10 significantly decreased the arsenic-induced expression of Fas ligand and caspase-3 via its free radical scavenging activity, anti-inflammatory action with reduced tumor necrosis factor-α production and inhibition of nuclear factor-κB expression thus prevent testicular injury and dysfunction induced by arsenic exposure (Fouad et al., 2011). Treatment with naringin also showed similar effect in amelioration of arsenic-induced testicular toxicity via down-regulation of oxido-inflammatory mediators and pro-inflammatory cytokines.
CONCLUSION
In conclusion, results of present study suggested that sodium arsenite altered the activities of oxido-nitrosative stress, pro-inflammatory cytokines in testis along with biochemical parameters. On the basis of this study, the use of naringin may be useful in the protection from sodium arsenite induced toxicity. Hence, naringin exerted its protective effects against sodium arsenite induced testicular toxicity via modulation of endogenous antioxidant enzyme, inhibition of release of oxido-inflammatory mediators such as MDA and NO as well as pro-inflammatory cytokines.
ACKNOWLEDGMENT
The authors would like acknowledge Dr. S.S. Kadam, Vice-Chancellor and Dr. K.R. Mahadik, Principal, Poona College of Pharmacy, Bharati Vidyapeeth Deemed University, Pune, India, for providing the necessary facilities to carry out the study.
REFERENCES
- Akondi, R.B., P. Kumar, A. Annapurna and M. Pujari, 2011. Protective effect of rutin and naringin on sperm quality in streptozotocin (STZ) induced Type 1 diabetic rats. Iran. J. Pharm. Res., 10: 585-596.
PubMed - Ali, M.M., A. Monira and A. El-Kader, 2004. The influence of naringin on the oxidative state of rats with streptozotocin-induced acute hyperglycaemia. Z. Naturforsch. C, 59: 726-733.
Direct Link - Bates, M.N., A.H. Smith and C. Hopenhayn-Rich, 1992. Arsenic ingestion and internal cancers: A review. Am. J. Epidemiol., 135: 462-476.
Direct Link - Beckman, J.S. and J.P. Crow, 1993. The chemistry of peroxynitrite a product from the reaction of nitric oxide and pathological implications of nitric oxide superoxide and peroxynitrite formation. Biochem. Soc. Trans., 21: 330-334.
PubMed - Benavente-Garcia, O. and J. Castillo, 2008. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular and anti-inflammatory activity. J. Agric. Food Chem., 56: 6185-6205.
CrossRefPubMedDirect Link - Bergh, A. and O. Soder, 1990. Interleukin-1β, but not interleukin-1α, induces acute inflammation-like changes in the testicular microcirculation of adult rats. J. Reprod. Immunol., 17: 155-165.
CrossRefDirect Link - Bertolero, F., G. Pozzi, E. Sabbioni and U. Saffiotti, 1987. Cellular uptake and metabolic reduction of pentavalent to trivalent arsenic as determinants of cytotoxicity and morphological transformation. Carcinogenesis, 8: 803-808.
CrossRefDirect Link - Bohler, T., J. Waiser, H. Hepburn, J. Gaedeke and C. Lehmann et al., 2000. TNF-α and IL-1α induce apoptosis in subconfluent rat mesangial cells. Evidence for the involvement of hydrogen peroxide and lipid peroxidation as second messengers. Cytokine, 12: 986-991.
CrossRefDirect Link - Cannon, J.G., R.G. Tompkins, J.A. Gelfand, H.R. Michie and G.G. Stanford et al., 1990. Circulating interleukin-1 and tumor necrosis factor in septic shock and experimental endotoxin fever. J. Infect. Dis., 161: 79-84.
CrossRefDirect Link - Chang, S.I., B. Jin, P. Youn, C. Park, J.D. Park and D.Y. Ryu, 2007. Arsenic-induced toxicity and the protective role of ascorbic acid in mouse testis. Toxicol. Applied Pharmacol., 218: 196-203.
CrossRefPubMedDirect Link - Choe, S.C., H.S. Kim, T.S. Jeong, S.H. Bok and Y.B. Park, 2001. Naringin has an antiatherogenic effect with the inhibition of intercellular adhesion molecule-1 in hypercholesterolemic rabbits. J. Cardiovasc. Pharmacol., 38: 947-955.
PubMed - Das, J., J. Ghosh, P. Manna, M. Sinha and P.C. Sil, 2009. Taurine protects rat testes against NaAsO2-induced oxidative stress and apoptosis via mitochondrial dependent and independent pathways. Toxicol. Lett., 187: 201-210.
CrossRefDirect Link - Flora, S.J., K. Bhatt and A. Mehta, 2009. Arsenic moiety in gallium arsenide is responsible for neuronal apoptosis and behavioral alterations in rats. Toxicol. Applied Pharmacol., 240: 236-244.
CrossRefDirect Link - Fouad, A.A., A.I. Al-Sultan and M.T. Yacoubi, 2011. Coenzyme Q10 counteracts testicular injury induced by sodium arsenite in rats. Eur. J. Pharmacol., 655: 91-98.
CrossRefPubMedDirect Link - Gerendai, I., P. Banczerowski and V. Csernus, 2005. Interleukin 1-β injected into the testis acutely stimulates and later attenuates testicular steroidogenesis of the immature rat. Endocrine, 28: 165-170.
CrossRefDirect Link - Ghosh, P., A.D. Kandhare, D. Gauba, K.S. Raygude and S.L. Bodhankar, 2012. Determination of efficacy, adverse drug reactions and cost effectiveness of three triple drug regimens for the treatment of Helicobacter pylori infected acid peptic disease patients. Asian Pac. J. Trop. Dis., 2: S783-S789.
CrossRefDirect Link - Ghosh, P., A.D. Kandhare, K.S. Raygude, D. Gauba, T.P. Gosavi and S.L. Bodhankar, 2012. Cigarette smoking and H. pylori infection: A meta-analysis of literature. Der Pharmacia Lettre, 4: 128-134.
Direct Link - Ghosh, P., A.D. Kandhare, K.S. Raygude, V.S. Kumar, A.R. Rajmane, M. Adil and S.L. Bodhankar, 2012. Determination of the long term diabetes related complications and cardiovascular events using UKPDS risk engine and UKPDS outcomes model in a representative western Indian population. Asian Pac. J. Trop. Dis., 2012: S642-S650.
CrossRefDirect Link - Gonzalez-Gallego, J., S. Sanchez-Campos and M.J. Tunon, 2007. Anti-inflammatory properties of dietary flavonoids. Nutr. Hosp., 22: 287-293.
PubMedDirect Link - Gosavi, T.P., P. Ghosh, A.D. Kandhare, V.S. Kumar, M. Adil, A.R. Rajmane and S.L. Bodhankar, 2012. Therapeutic effect of H. pylori nosode, a homeopathic preparation in healing of chronic H. pylori infected ulcers in laboratory animals. Asian Pac. J. Trop. Dis., 2: S603-S611.
CrossRefDirect Link - Gosavi, T.P., A.D. Kandhare, P. Ghosh and S.L. Bodhankar, 2012. Anticonvulsant activity of Argentum metallicum, a homeopathic preparation. Der Pharmacia Lettre, 4: 626-637.
Direct Link - Hayden, M.R., 2002. Islet amyloid, metabolic syndrome and the natural progressive history of type 2 diabetes mellitus. Jop, 3: 126-138.
Direct Link - Jagetia, G.C. and T.K. Reddy, 2002. The grapefruit flavanone naringin protects against the radiation-induced genomic instability in the mice bone marrow: A micronucleus study. Mutat. Res./Fundam. Mol. Mech. Mutagen., 519: 37-48.
CrossRefDirect Link - Jagetia, G.C., V.A. Venkatesha and T.K. Reddy, 2003. Naringin, a citrus flavonone, protects against radiation-induced chromosome damage in mouse bone marrow. Mutagenesis, 18: 337-343.
CrossRefDirect Link - Jana, K., S. Jana and P.K. Samanta, 2006. Effects of chronic exposure to sodium arsenite on hypothalamo-pituitary-testicular activities in adult rats: Possible an estrogenic mode of action. Reprod. Biol. Endocrinol., Vol. 4.
CrossRef - Javed, A., A.D. Kandhare, M.V.K. Patil, A. Sinha, P. Singh, P. Ghosh and S.L. Bodhankar, 2013. Naringin alleviates paracetamol induced hepatotoxicity via modulation of endogenous enzyme and oxidative stress in wistar rats. Proceedings of the 46th Annual Conference of Indian Pharmacological Society, December 16-18, 2013, Nimhans Convention Centre, Bangalore, India.
- Kaltreider, R.C., A.M. Davis, J.P. Lariviere and J.W. Hamilton, 2001. Arsenic alters the function of the glucocorticoid receptor as a transcription factor. Environ. Health Persp., 109: 245-251.
PubMedDirect Link - Kamble, H., A.D. Kandhare, S. Bodhankar, V. Mohan and P. Thakurdesai, 2013. Effect of low molecular weight galactomannans from fenugreek seeds on animal models of diabetes mellitus. Biomed. Aging Pathol., 3: 145-151.
CrossRefDirect Link - Kandhare, A.D., K.S. Raygude, P. Ghosh and S.L. Bodhankar, 2011. The ameliorative effect of fisetin, a bioflavonoid, on ethanol-induced and pylorus ligation-induced gastric ulcer in rats. Int. J. Green Pharm., 5: 236-243.
Direct Link - Kandhare, A.D., S.L. Bodhankar, V. Singh, V. Mohan and P.A. Thakurdesai, 2013. Anti-asthmatic effects of type-A procyanidine polyphenols from cinnamon bark in ovalbumin-induced airway hyperresponsiveness in laboratory animals. Biomed. Aging Pathol., 3: 23-30.
CrossRefDirect Link - Kandhare, A.D., P. Ghosh, A.E. Ghule and S.L. Bodhankar, 2013. Elucidation of molecular mechanism involved in neuroprotective effect of Coenzyme Q10 in alcohol-induced neuropathic pain. Fundam. Clin. Pharmacol., 27: 603-622.
CrossRefPubMedDirect Link - Kandhare, A.D., P. Ghosh, A.E. Ghule, G.N. Zambare and S.L. Bodhankar, 2013. Protective effect of Phyllanthus amarus by modulation of endogenous biomarkers and DNA damage in acetic acid induced ulcerative colitis: Role of phyllanthin and hypophyllanthin. Apollo Med., 10: 87-97.
CrossRefDirect Link - Kandhare, A.D., V.S. Kumar, M. Adil, A.R. Rajmane, P. Ghosh and S.L. Bodhankar, 2012. Investigation of gastro protective activity of Xanthium strumarium L. by modulation of cellular and biochemical marker. Orient. Pharmacy Exp. Med., 12: 287-299.
CrossRefDirect Link - Kandhare, A.D., K.S. Raygude, P. Ghosh, A.E. Ghule and S.L. Bodhankar, 2012. Neuroprotective effect of naringin by modulation of endogenous biomarkers in streptozotocin induced painful diabetic neuropathy. Fitoterapia, 83: 650-659.
CrossRefPubMedDirect Link - Kandhare, A.D., K.S. Raygude, P. Ghosh, A.E. Ghule and S.L. Bodhankar, 2012. Therapeutic role of curcumin in prevention of biochemical and behavioral aberration induced by alcoholic neuropathy in laboratory animals. Neurosci. Lett., 511: 18-22.
CrossRefPubMedDirect Link - Kandhare, A.D., K.S. Raygude, P. Ghosh, A.E. Ghule, T.P. Gosavi, S.L. Badole and S.L. Bodhankar, 2012. Effect of hydroalcoholic extract of Hibiscus rosa sinensis Linn. leaves in experimental colitis in rats. Asian Pac. J. Trop. Biomed., 2: 337-344.
CrossRefPubMedDirect Link - Kandhare, A.D., K.S. Raygude, P. Ghosh, T.P. Gosavi and S.L. Bodhankar, 2011. Patentability of animal models: India and the globe. Int. J. Pharm. Biol. Arch., 2: 1024-1032.
Direct Link - Kandhare, A.D., K.S. Raygude, V.S. Kumar, A.R. Rajmane and A. Visnagri et al., 2012. Ameliorative effects quercetin against impaired motor nerve function, inflammatory mediators and apoptosis in neonatal streptozotocin-induced diabetic neuropathy in rats. Biomed. Aging Pathol., 2: 173-186.
Direct Link - Kawaguchi, K., S. Kikuchi, H. Hasegawa, H. Maruyama, H. Morita and Y. Kumazawa, 1999. Suppression of Lipopolysaccharide-induced tumor necrosis Factor-release and liver injury in mice by naringin. Eur. J. Pharmacol., 368: 245-250.
CrossRefDirect Link - Kobayashi, Y., X. Cui and S. Hirano, 2005. Stability of arsenic metabolites, arsenic triglutathione [As(GS)3] and methylarsenic diglutathione [CH3As(GS)2], in rat bile. Toxicology, 211: 115-123.
CrossRefDirect Link - Liu, S.X., M. Athar, I. Lippai, C. Waldren and T.K. Hei, 2001. Induction of oxyradicals by arsenic: Implication for mechanism of genotoxicity. Proc. Natl. Acad. Sci. USA., 98: 1643-1648.
CrossRefDirect Link - Mann, K.K., A.M. Padovani, Q. Guo, A.L. Colosimo, H.Y. Lee, J.M. Kurie and W.H. Miller Jr., 2005. Arsenic trioxide inhibits nuclear receptor function via SEK1/JNK-mediated RXRα phosphorylation. J. Clin. Invest., 115: 2924-2933.
CrossRef - Martin, M.J., E. Marhuenda, C. Perez-Guerrero and J.M. Franco, 1994. Antiulcer effect of naringin on gastric lesions induced by ethanol in rats. Pharmacology, 49: 144-150.
CrossRefDirect Link - Morakinyo, A.O., P.U. Achema and O.A. Adegoke, 2010. Effect of Zingiber officinale (ginger) on sodium Arsenite-induced reproductive toxicity in male rats. Afr. J. Biomed. Res., 13: 39-45.
Direct Link - Mukherjee, S., D. Das, M. Mukherjee, A.S. Das and C. Mitra, 2006. Synergistic effect of folic acid and vitamin B12 in ameliorating Arsenic-induced oxidative damage in pancreatic tissue of rat. J. Nutr. Biochem., 17: 319-327.
CrossRefDirect Link - Nandi, D., R.C. Patra and D. Swarup, 2005. Effect of cysteine, methionine, ascorbic acid and thiamine on Arsenic-induced oxidative stress and biochemical alterations in rats. Toxicology, 211: 26-35.
CrossRefDirect Link - Pant, N., R.C. Murthy and S.P. Srivastava, 2004. Male reproductive toxicity of sodium arsenite in mice. Human Exp. Toxicol., 23: 399-403.
CrossRef - Patil, M.V.K., A.D. Kandhare and S.D. Bhise, 2012. Anti-inflammatory effect of Daucus carota root on experimental colitis in rats. Int. J. Pharm. Pharm. Sci., 4: 337-343.
Direct Link - Patil, M.V.K., A.D. Kandhare and S.D. Bhise, 2012. Anti-arthritic and Anti-inflammatory activity of Xanthium srtumarium L. ethanolic extract in freund's complete adjuvant induced arthritis. Biomed. Aging Pathol., 2: 6-15.
CrossRefDirect Link - Patil, M.V.K., A.D. Kandhare and S.D. Bhise, 2012. Effect of aqueous extract of Cucumis sativus Linn. fruit in ulcerative colitis in laboratory animals. Asian Pac. J. Trop. Biomed., 2: S962-S969.
CrossRefDirect Link - Patil, M.V.K., A.D. Kandhare, P. Ghosh and S.D. Bhise, 2012. Determination of role of GABA and nitric oxide in anticonvulsant activity of Fragaria vesca L. ethanolic extract in chemically induced epilepsy in laboratory animals. Oriental Pharmacy Exp. Med., 12: 255-264.
CrossRefDirect Link - Punithavathi, V.R., R. Anuthama and P.S.M. Prince, 2008. Combined treatment with naringin and vitamin C ameliorates Streptozotocin-induced diabetes in male Wistar rats. J. Applied Toxicol., 28: 806-813.
CrossRefPubMedDirect Link - Radi, R., 2004. Nitric oxide, oxidants and protein tyrosine nitration. Proc. Natl. Acad. Sci. USA., 101: 4003-4008.
CrossRef - Rajadurai, M. and P.S.M. Prince, 2007. Preventive effect of naringin on isoproterenol-induced cardiotoxicity in Wistar rats: An in vivo and in vitro study. Toxicology, 232: 216-225.
CrossRefDirect Link - Ramanathan, K., B.S. Balakumar and C. Panneerselvam, 2002. Effects of ascorbic acid and a-tocopherol on Arsenic-induced oxidative stress. Hum. Exp. Toxicol., 21: 675-680.
CrossRefDirect Link - Ramanathan, K., M.A. Contreras, C.L. Perkins, S. Asher and F.S. Hasoon et al., 2003. Properties of 19.2% efficiency ZnO/CdS/CuInGaSe2 Thin-film solar cells. Prog. Photovolt. Res. Appl., 11: 225-230.
CrossRef - Raygude, K.S., A.D. Kandhare, P. Ghosh and S.L. Bodhankar, 2012. Anticonvulsant effect of fisetin by modulation of endogenous biomarkers. Biomed. Preventive Nutr., 2: 215-222.
CrossRefDirect Link - Raygude, K.S., A.D. Kandhare, P. Ghosh, A.E. Ghule and S.L. Bodhankar, 2012. Evaluation of ameliorative effect of quercetin in experimental model of alcoholic neuropathy in rats. Inflammopharmacology, 20: 331-341.
CrossRefPubMedDirect Link - Sakurai, T., T. Kaise and C. Matsubara, 1998. Inorganic and methylated arsenic compounds induce cell death in murine macrophages via different mechanisms. Chem. Res. Toxicol., 11: 273-283.
CrossRef - Sarkar, M., G.R. Chaudhuri, A. Chattopadhyay and N.M. Biswas, 2003. Effect of sodium arsenite on spermatogenesis, plasma gonadotrophins and testosterone in rats. Asian J. Androl., 5: 27-31.
Direct Link - Sharpe, R.M., K. Donachie and I. Cooper, 1988. Re-evaluation of the intratesticular level of testosterone required for quantitative maintenance of spermatogenesis in the rat. J. Endocrinol., 117: 19-26.
CrossRefDirect Link - Sharpe, R.M., S. Maddocks, M. Millar, J.B. Kerr, P.T. Saunders and C. McKinnell, 1992. Testosterone and spermatogenesis. Identification of stage-specific, androgen-regulated proteins secreted by adult rat seminiferous tubules. J. Androl., 13: 172-184.
PubMed - Sies, H., 1999. Glutathione and its role in cellular functions. Free Radic. Biol. Med., 27: 916-921.
CrossRefPubMedDirect Link - Sikka, S.C., 2004. Role of oxidative stress and antioxidants in andrology and assisted reproductive technology. J. Androl., 25: 5-18.
CrossRefPubMedDirect Link - Singh, D., V. Chander and K. Chopra, 2004. Protective effect of naringin, a bioflavonoid on Glycerol-induced acute renal failure in rat kidney. Toxicology, 201: 143-151.
CrossRefDirect Link - Singh, N., D. Kumar, K. Lal, S. Raisuddin and A.P. Sahu, 2010. Adverse health effects due to arsenic exposure: Modification by dietary supplementation of jaggery in mice. Toxicol. Applied Pharmacol., 242: 247-255.
CrossRefDirect Link - Smith, A.H., C. Hopenhayn-Rich, M.N. Bates, H.M. Goeden and I. Hertz-Picciotto et al 1992. Cancer risks from arsenic in drinking water. Environ. Health Perspect., 97: 259-267.
Direct Link - Steffensen, K.R. and J.A. Gustafsson, 2004. Putative metabolic effects of the Liver X Receptor (LXR). Diabetes, 53: S36-S42.
CrossRef - Taepongsorat, L., P. Tangpraprutgul, N. Kitana and S. Malaivijitnond, 2008. Stimulating effects of quercetin on sperm quality and reproductive organs in adult male rats. Asian J. Androl., 10: 249-258.
PubMed - Tchounwou, P.B., B.A. Wilson, A.A. Abdelghani, A.B. Ishaque and A.K. Patlolla, 2002. Differential cytotoxicity and gene expression in human liver carcinoma (HepG2) cells exposed to arsenic trioxide and monosodium acid methanearsonate (MSMA). Int. J. Mol. Sci., 3: 1117-1132.
CrossRef - Tseng, C.H., 2004. The potential biological mechanisms of Arsenic-induced diabetes mellitus. Toxicol. Appl. Pharmacol., 197: 67-83.
PubMedDirect Link - Usoh, I.F., E.J. Akpan, E.O. Etim and E.O. Farombi, 2005. Antioxidant actions of dried flower extracts of Hibiscus sabdariffa L. on sodium arsenite-induced oxidative stress in rats. Pak. J. Nutr., 4: 135-141.
CrossRefDirect Link - Visnagri, A., A.D. Kandhare, S. Chakravarty, P. Ghosh and S.L. Bodhankar, 2014. Hesperidin, a flavanoglycone attenuates experimental diabetic neuropathy via modulation of cellular and biochemical marker to improve nerve functions. Pharm. Biol.
CrossRef - Visnagri, A., A.D. Kandhare, P. Ghosh and S.L. Bodhankar, 2013. Endothelin receptor blocker bosentan inhibits hypertensive cardiac fibrosis in pressure overload-induced cardiac hypertrophy in rats. Cardiovasc. Endocrinol., 2: 85-97.
CrossRefDirect Link - Visnagri, A., A.D. Kandhare, V.S. Kumar, A.R. Rajmane and A. Mohammad et al., 2012. Elucidation of ameliorative effect of co-enzyme Q10 in streptozotocin-induced diabetic neuropathic perturbation by modulation of electrophysiological, biochemical and behavioral markers. Biomed. Aging Pathol., 2: 157-172.
CrossRefDirect Link - Willcox, J.K., S.L. Ash and G.L. Catignani, 2004. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr., 44: 275-295.
CrossRefPubMedDirect Link - Yamanaka, K., A. Hasegawa, R. Sawamura and S. Okada, 1991. Cellular respons to oxidative damage in lung induced by the administration of dimethylarsinic acid, a major metabolite of inorganic arsenics in mice. Toxicol. Applied Pharmacol., 108: 205-215.
Direct Link - Yousef, M.I., F.M. El-Demerdash and F.M.E. Radwan, 2008. Sodium arsenite induced biochemical perturbations in rats: Ameliorating effect of curcumin. Food Chem. Toxicol., 46: 3506-3511.
CrossRefDirect Link