
Racheal Oluwayemisi Fashogbon
Department of Biological Sciences, Ajayi Crowther University, Oyo, Nigeria
LiveDNA: 234.34604
Bukola Adebayo-Tayo
Department of Microbiology, University of Ibadan, Oyo State, Nigeria
Jadesola Sanusi
Department of Biological Sciences, Crescent University, Abeokuta, Nigeria







ABSTRACT
Background and Objective: The extracellular polysaccharide secreted by Lactic Acid Bacteria (LAB) has been used in the food industry due to their viscosity, bio-thickening properties as well as immunomodulatory potentials. Due to the increase in the demand for EPS, there is a need to study the production of extracellular polysaccharides produced by Lactic Acid Bacteria (LAB) under different production conditions for optimum EPS production. Materials and Methods: Five EPS producing LAB isolated from fermented dairy products were collected from the culture collection Centre, maintained in De Ma, Rogosa and Sharpe (MRS) broth and the EPS producing LAB were molecularly characterized. Modified Exopolysaccharide Selection Medium (mESM) was used to produce the EPS. Results: The EPS producers were molecularly characterized as Lactobacillus tucetti FASHADFF2, Lactobacillus delbrueckii FASHADYG2, Weissella sp. FASHADFF1, Weissella sp. FASHADWR2 and Leuconostoc mesenteroides FASHADWR1. The EPS produced by the five LAB strains ranged from 109.11-185.02 mg L–1. About 35°C and pH 5 supported the highest EPS production by the isolates. Glucose, sucrose, yeast extract, alanine and folic acid supported the highest EPS production. Conclusion: This study demonstrated that all the five LAB strains were EPS producers and they were 99-100% molecularly related to the identified species of LAB. Optimizing the production of EPS by the LAB strains yields a higher amount of EPS.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Racheal Oluwayemisi Fashogbon, Bukola Adebayo-Tayo and Jadesola Sanusi, 2021. Optimization of Extracellular Polysaccharide Substances from Lactic Acid Bacteria Isolated from Fermented Dairy Products. Microbiology Journal, 11: 1-11.
DOI: 10.3923/mj.2021.1.11
URL: https://scialert.net/abstract/?doi=mj.2021.1.11
DOI: 10.3923/mj.2021.1.11
URL: https://scialert.net/abstract/?doi=mj.2021.1.11
INTRODUCTION
An increase in the demand for Bacteria Exopolysaccharide (EPS) produced by Lactic Acid Bacteria (LAB) has been greatly researched1,2. Exopolysaccharides (EPS) are long-chain polysaccharides with high molecular weight secreted by microbial cells consisting of non-carbohydrate substitutes such as acetate, pyruvates and phosphate3,4. Microbial EPS is mainly in two distinct forms depending on their locations. They could be capsular EPS which are cell-bound EPSs that closely adhere to the bacterial surface (cEPSs) or Free EPS which are EPSs that are released into the surrounding medium (fEPSs). However, the production of EPSs by LAB is often distinguished as ropy or non-ropy EPS. The ropy EPS is secreted into the surrounding medium while the non-ropy EPS remains or adhered to the cell surface creating a discrete covering5. Due to the costs, fluctuating prices of plant and algal gums as well as the increased demand for polymers of natural origin used in different industries, it has encouraged manufacturers to look industrially into the making of gums which are polysaccharides derived from microbial origin6. Microbial polysaccharides may be natural or semisynthetic polymers with different structures and they are manufactured via sugar fermentation by some microorganisms like lactic acid bacteria. EPS is a water-soluble polymer, thereby majorly used as a thickening, gelling and suspending agent due to their ability to dissolve, disperse or swell in water are mainly7,8. LAB involved in food fermentation is considered as “generally recognized as safe” (GRAS) due to their widespread occurrence in food, ability to produce EPS in addition to their long-lived applications9,10. LAB enhanced shelf life, improved digestibility, pleasant sensory profile, improve the texture and flavor of food products. These properties are ensured through the production of certain useful metabolites which include organic acids, ethanol, aroma compounds bacteriocins, enzymes and exopolysaccharides11. Streptococcus, Lactococcus, Pediococcus, Lactobacillus, Leuconostoc and Weissella species are LAB strains that frequently produced EPSs12,13. Microbial EPS is known for its antioxidant potential with varied activity. The EPS produced by Weissella confusa also have immunomodulatory potential2. Optimization conditions such as the composition of the culture medium (carbon, nitrogen and cation sources), incubation time, pH, temperature, oxygen concentration, agitation made EPS released by LAB have a great effect in microbial polysaccharide production14,15 thereby enhancing taste perception16,17. Optimization of the growth environment is a critical point for the largest EPS yield18. The amount as well as the EPS composition produced by LAB are strongly influenced by culture medium and fermentation conditions19,20. An additional physiological benefit of EPS is that it enhances the colonization of probiotic bacteria because it can be retained in the gastrointestinal tract for a long period. The EPS released by LAB is being used in the food industries because of its emulsifying and bio-thickening potential. In recent times, it was discovered that there is a need to develop healthy foods and this development has posed lots of challenges in the food industry which resulted in a lot of research into the production of EPSs by LAB. The research work is therefore aimed at the production of EPS from LAB, molecularly characterize and determine the effect of physicochemical parameters on EPS produced by LAB strains.
MATERIALS AND METHODS
Study area: This study was carried out at the Department of Microbiology, Physiology Laboratory and the Research Central Laboratory, University of Ibadan, Oyo State from June, 2017-January, 2018.
Collection of cultures: Five EPS producing LAB isolated from fermented milk and yogurt were collected from the culture collection Centre of our previous work in the Microbial Physiology Unit, Department of the Microbiology University of Ibadan, Ibadan, Nigeria21. The LAB was maintained in De Man, Rogosa and Sharpe (MRS) broth (Difco)22 and stored in a refrigerator at 28°C for three days.
Molecular characterization of EPS producing LAB strains
Genomic DNA extraction: In this study, an amplified segment of the 16S rDNA gene of each LAB isolate was sequenced and the sequences were compared to the strains in the National Centre for Biotechnology Information (NCBI) BLAST library. The five LAB isolates were subcultures on MRS agar and incubated at 30°C for 18 hrs. The DNA was extracted from the LAB isolates and purified using the DNA Extraction kit (Invisorb Spin) as instructed by the manufacturer23. The 18 hrs old LAB culture of each isolate were centrifuged at 10,000 ×g for 10 min at 4°C. The twice with Dnase-free deionized water and re-suspended in 100 μL Chelex. The suspension was incubated for 20 min at 56°C, vortexed, boiled and freeze cycle for 5 min. Centrifugation was done at 14,000×g for 5 min at 4°C. Total 20 μL Proteinase ? (20 mg mL–1) (Thermo Scientific, USA) was inoculated into 80 μL of the supernatant and was incubated at 65°C for 60 min. The Cell debris was collected by centrifugation at 14,000×g for 5 min at 4°C and the supernatant was stored at -20°C. A spectrophotometer (Shimadzu UV-1800, Japan) was used to measure the concentrated and pure extracted DNA24.
Molecular identification of LAB by sequencing of 16S rRNA gene: Fragments of the 16S rRNA gene of LAB isolates were amplified distinctly using PCR and was performed in a thermal cycler (Bio-Rad Laboratories Inc., Hercules, USA). Each PCR contained 10 μM of each 27 F (5’ AGAGTTTGATCCTGGCTCAG 3’) and 1492 R (5’ GGTTACCTTGTTACGACTT 3’) universal primer (Lane, 1991), 1 μL of the extracted DNA, 2.5 mM of each dNTP, 1 μL taqTM DNA polymerase (Fermentas, St. Leon-Rot, Germany) with 5 μL PCR buffer. Cycling conditions were set at an initial denaturation at 95°C for 60 sec followed by 35 cycles of denaturation at 94°C for 60 sec, annealing at 50°C for 30 sec and primer elongation at 72°C for 60 sec and final elongation at 72°C for 5 min. Gel electrophoresis (3% Agarose gel, 1 μL loading dye with 5 μL PCR products and stained with ethidium bromide for gel documentation) was used to separate the amplicons of LABs. PCR yields were purified using Wizard® SV Gel and PCR Clean-Up System (Promega, Madison, WI, USA) and sequenced with universal primer 27 F. The nucleotide sequences were analyzed by BLAST algorithm packages (Biological sequence alignment editor for Win95/98/NT/XP/7).
Production of EPS: Modified Exopolysaccharide Selection Medium (mESM) was used to produce the EPS from the LAB22. Seed culture of the EPS producing LAB was prepared by transferring 0.5 mL of the stock frozen culture to 10 mL of MRS broth and incubated for 16 h at 30°C. The resulting culture was transferred (2% v/v) into Exopolysaccharide Selection Medium (mESM). The medium contains 5% skimmed milk (Oxoid), 0.35% yeast extract (Oxoid), 0.35% peptone (Difco) and 5% glucose (BDH) and incubated at 30°C for 16 hrs25,26. The fermentation medium was heated in a boiling water bath for 15 min. to inactivate enzymes and then cooled to room temperature. 250 μg of 80% (w/v) trichloroacetic acid was added to the medium and stored at 4°C to precipitate the protein. The cleared supernatant was obtained by centrifugation (Sigma 3-18 K centrifuge, Germany) at 15,000 g for 20 min at 4°C to remove cells and coagulated proteins. The EPS was precipitated from the supernatant with three volumes of old ethanol (4°C, 100%) followed by overnight incubation at -20°C. The resulting precipitate (EPS) was collected after centrifugation.
Extraction and quantification of EPS: The total sugar concentration was determined by using the phenol-sulfuric acid method. 0.1 mL of EPS samples were diluted in 2.0 mL of distilled water. 1.0 mL of 6% phenol and 5.0 mL of 95% (v/v) sulfuric acid was added to the solution. Absorbance was read at 490 nm. The concentration of EPS was determined in triplicate and 2.0 mL distilled water was used as blank. The EPS content of each sample was calculated by the standard curve. The glucose standard curve was prepared for the quantitative determination with some modifications27. The fermentation medium was centrifuged at 15,000×g for 15 mins at 4°C. The EPS was precipitated at 4°C by the addition of 2 volumes of ethanol (100%). The resulting precipitate was collected after centrifugation (15,000 xg for 15 mins at 4°C). Total sugar was determined using the phenol-sulphuric acid method27.
Effect of physicochemical parameters on EPS production: To increase the yield of EPS, the following media components and cultivation conditions were investigated28. The conditions include: cultivation temperature (15, 20, 25, 30, 35, 40 and 45°C), pH (3, 4, 5 and 6), carbon sources in the fermenting medium was replaced with glucose, sucrose, mannitol, lactose, maltose and fructose, nitrogen source was replaced with peptone, yeast extract, urea and sodium acetate, amino acid was substituted with alanine, lysine, tryptophan and valine while vitamins were replaced with riboflavin and folic acid. The supplemented medium was sterilized, inoculated and incubated for 24 hrs.
Statistical analysis: Results from the experimental design were statistically subjected to analysis of variance (ANOVA) using SPSS (version 11.0, Chicago, IL). Probability values (p≤0.05) was considered significant to indicate the difference.
RESULTS AND DISCUSSION
Molecular characterization by gene sequencing: The results of the sequence similarity (%) by BLASTN in the GenBank of the NCBI are shown in Table 1. The 16S rRNA gene sequences obtained in this study exhibited 99-100% of sequences in NCBI. Genotypic identification of the LAB strains includes 2 strains of Lactobacillus, 2 strains of Weissella and one strain of Leuconostoc. Lactobacillus tucetti FASHADFF2 had 100% relatedness to Lactobacillus tucceti, Lactobacillus delbrueckii FASHADYG2, Weissella sp. FASHADFF1, Weissella sp. FASHADWR2 and Leuconostoc mesenteroides FASHADWR1 had 99 and 100% relatedness to Lactobacillus delbrueckii, Weissella confusa, Weissella koreensis and Leuconotoc mesenteroides. The molecular methods are important for bacterial identification. The molecular method is more reliable in identifying LAB by giving the precise and accurate strains of microorganisms.
| Table 1: Comparative taxonomy identification of LAB isolated in the study by sequencing of 16S rRNA coding gene according to the database | |||||
| Isolates code | Phenotypic Identification | Genotypic identification | Similarities (%) | Accession number | |
| WCFF1 | Weissella confuse | Weissella confusa JCM 1093 | 99 | Weissella confusa FASHADFF1 | MH790313 |
| LTFF2 | Lactobacillus tucceti | Lactobacillus tucceti strain CECT 5920 | 100 | Lactobacillus tucceti FASHADFF2 | MH790316 |
| LMWR1 | Leuconostoc mesenteroides | Leuconostoc mesenteroides srain ATCC8293 | 100 | Leuconostoc mesenteroides FASHADWR1 | MH790314 |
| WKWR2 | Weissella koreensis | Weissella koreensis strain S-5623 | 99 | Weissella koreensis FASHADWR2 | MH790312 |
| LDYG2 | Lactobacillus delbrueckii | Lactobacillus delbrueckii subsp. bulgaricus ATCC 11842 | 99 | Lactobacillus delbrueckii FASHADYG2 | MH790315 |
Molecular methods have gained acceptance for accuracy and reliability for microbial identification to validate phenotypically determined taxa, it was however observed that they do not always correlate with phenotypic characterization29,30. Polymerase Chain Reaction (PCR) based methods is essential for specific classification and detection of lactic acid bacteria strains31. Blast search analysis using the whole 16S rRNA sequence resulted in identification above 90% level, which is close to the considered cut-off value indicating species identity32. Gene sequencing, BLAST searching at NCBI and submission to GenBank identified the five EPS producing LAB isolates as Weissella confusa FASHADFF1, Weissella koreensis FASHADFF2, Leuconostoc mesenteroides FASHADWR1, Lactobacillus tucceti FASHADFF2 and Lactobacillus delbrueckii FASHADYG2.
Phylogenetic trees: The phylogenetic trees of the isolated LAB strains based on 16S rDNA sequences (Neighbor-Joining Analysis of CLC bio software) are shown in Fig. 1(a-e).
Quantification of EPS produced by the LAB strains: The EPS production by the LAB strain is shown in Fig. 2. There was a significant difference (p<0.05) in EPS produced by all the LAB isolates. The EPS production of five (5) LAB strains (L. tucceti FASHADFF2, L. delbrueckii FASHADYG2, Weissella koreensis FASHADWR2, Weissella confusa FASHADFF1 and L. mesenteroides FASHADWR1) ranged from 109.11-185.02 mg L–1. L. delbrueckii FASHADYG2 had the highest amount of EPS (185.02 mg L–1) production while the least EPS production was observed in Leuconostoc mesenteroides (109.11 mg L–1). This result is in support of the work of Mostefaoui et al.33, who observed that Lactobacillus strain had the highest EPS yield ranging from 60-740 mg L–1. Laws and Marshall3 obtained an EPS yield of 175 mg L–1 produced by Lactobacillus delbrueckii subsp. bulgaricus LY03. However, Mozzi et al.12 reported that Lactobacillus sakei 0-1 produced an EPS yield of 1375 mg L–1.
Physicochemical parameters: The effect of temperature on EPS production by the selected LAB isolates is shown in Fig. 3. There was a significant increase in EPS production as the temperature increases to 35°C and then gradual reduction began to occur in the EPS yield. L. delbrueckki FASHADYG1 and W. confusa FASHADFF1 produced the highest quantity of EPS (1841 and 1816 mg L–1) at 35°C. W. confusa FASHADFF1, Leuconostoc mesenteroides FASHADWR1 and W. koreensis FASHADWR2 produced the least EPS (908, 807 and 741 mg L–1) at 35°C. EPS yield reduced generally at 40°C. Temperature is a critical factor in the synthesis of polysaccharides.
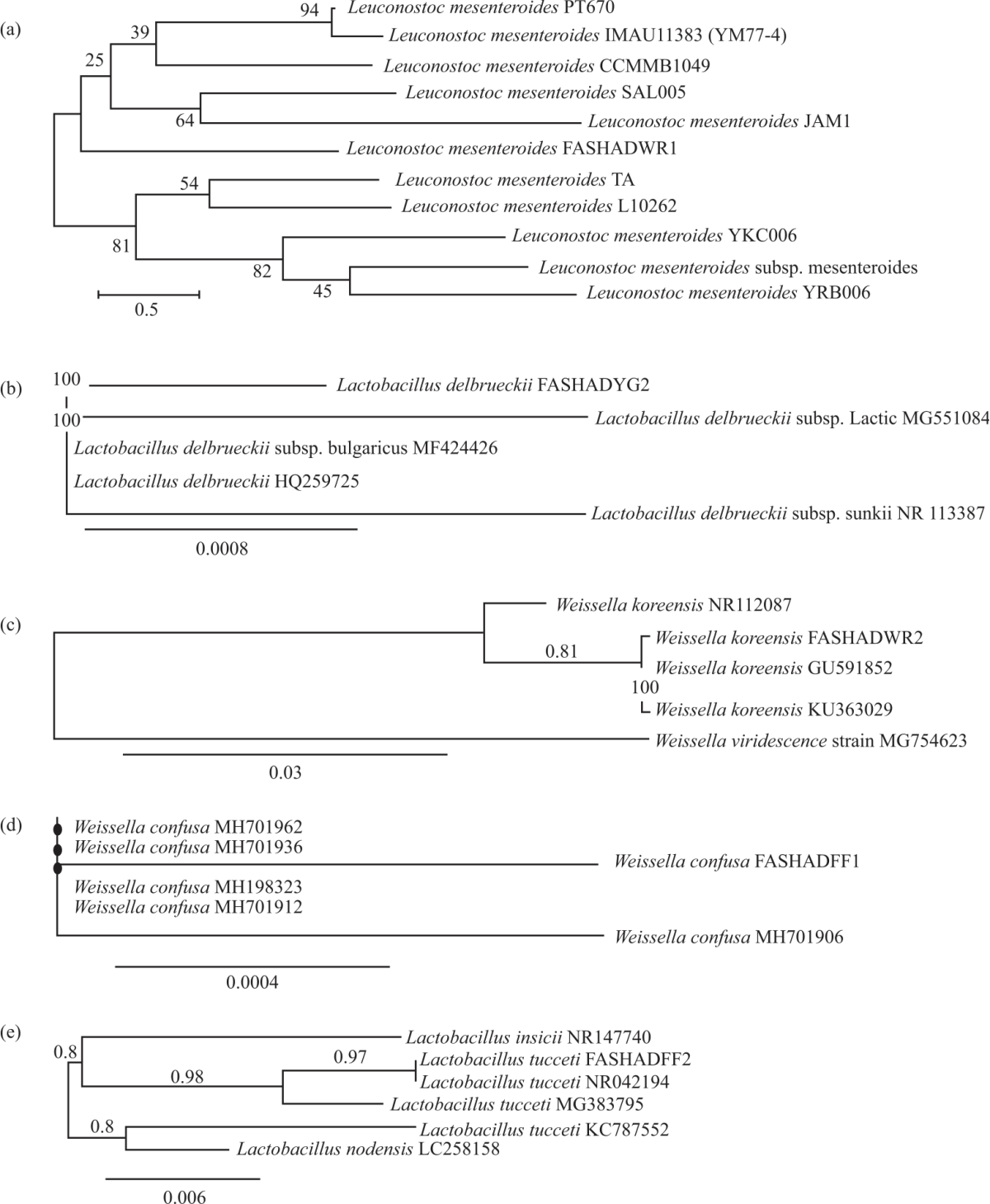 |
| Fig. 1(a-e): | Phylogenetic tree of different isolated LAB strains (a) Leuconostoc mesenteroides FASHADWR2 (LMWR1), (b) Lactobacillus delbrueckii FASHADYG2 (LDYG2), (c) Weissella koreensis FASHADWR2 (WKWR2), (d) Weissella confusa FASHADFF1 (WCFF1) and (e) Lactobacillus tucceti FASHADFF2 (LTFF2) |
The highest EPS production was recorded at 35°C by Lactobacillus delbrueckii FASHADYG2. This could be because the LAB isolates in this research work are mesophiles. Joshi and Koiji34 reported that the maximum temperature for EPS production was at 37°C. This correlates with the research of Gamar et al.35 where the optimal temperature for its growth was in the range of 30-37°C. Prasanna et al.36 also observed that the highest production of polysaccharides occurred between 25-35°C. However, studies have shown that species of thermophilic LAB such as Streptococcus thermophilus LY03 and Lactobacillus delbrueckii have a better EPS production at 42°C and above 45°C, respectively3,37.
The effect of pH on EPS production by the LAB strains is shown in Fig. 4. W. confusa FASHADFF1, W. koreensis FASHADWR2 and Lactobacillus delbrueckii FASHADYG2 supported the highest EPS yield at pH 5. pH 6 also supported the best for EPS production (1369 mg L–1) by W. confusa FASHADFF1. However, Leuconostoc mesenteroides FASHADWR1 produces the least EPS at pH 5 and pH 6. Generally, Acidic pH leads to high EPS production until pH 5 where there was a reduction.
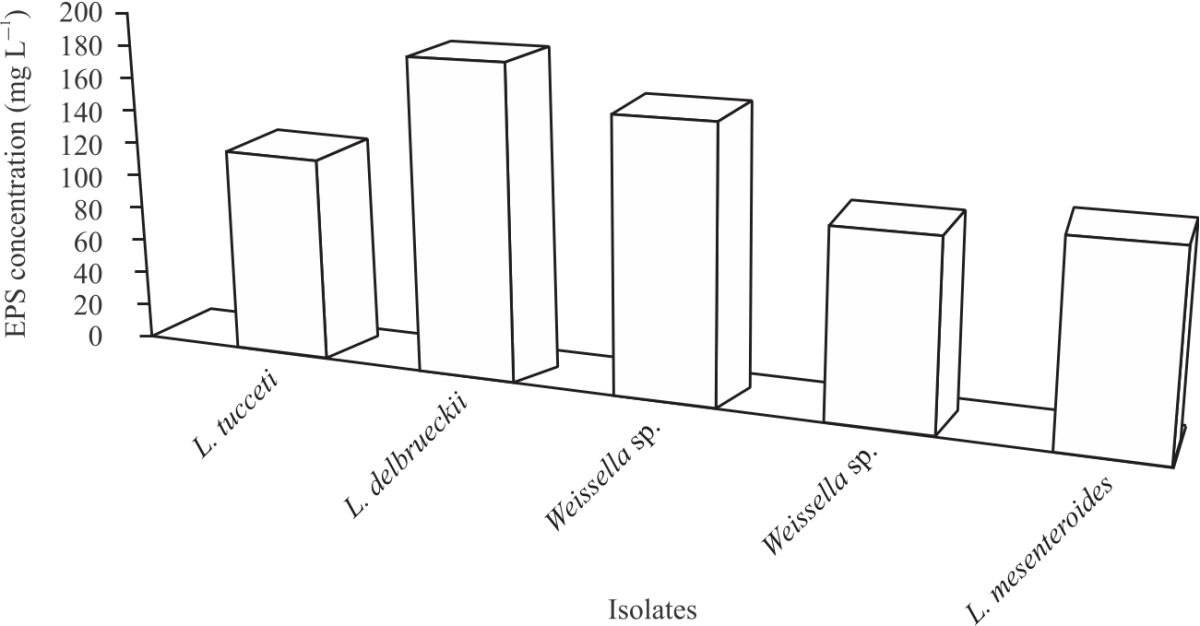 |
| Fig. 2: | Production of EPS by the LAB strains |
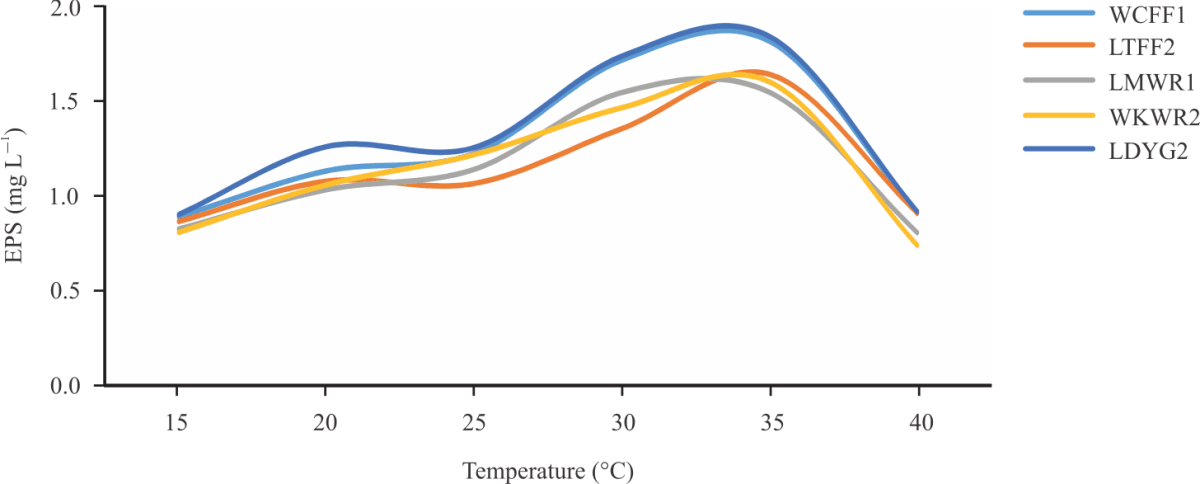 |
| Fig. 3: | Effect of temperature on EPS production by the selected LAB strains |
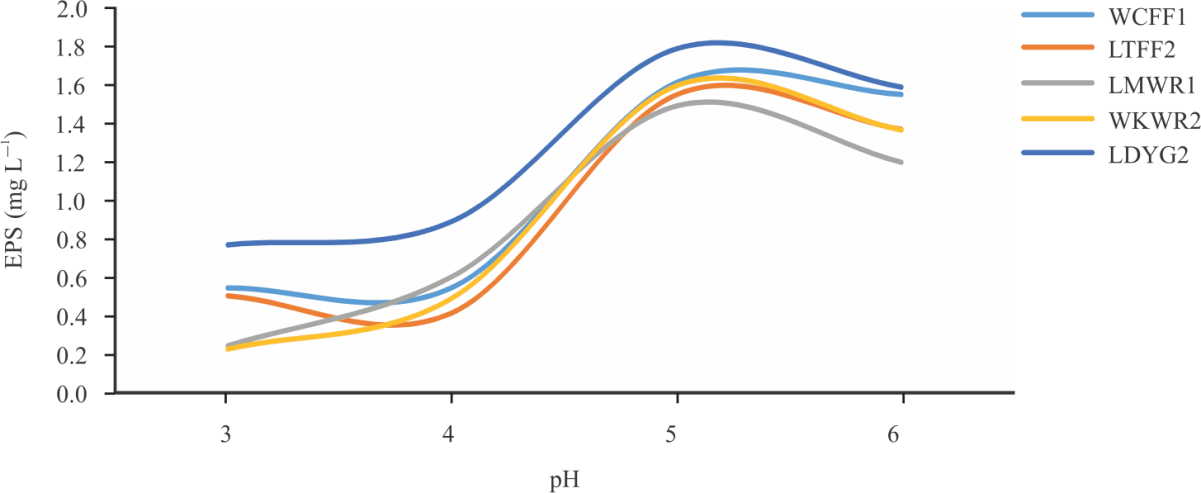 |
| Fig. 4: | Effect of pH on EPS production by the selected LAB strains |
Reduction in pH during fermentation occurs due to the fermentative transformation of carbohydrates to lactic acid and acetic acid by the isolates. The ability of LAB to lower the pH of fermented food leads to a hindrance of food spoilage which brings about an increase in its shelf life. By lowering the pH and acid production (acetic, lactic and carbonic), LAB adds to the preservation of food by releasing a vast array of antimicrobial compounds and proteins38. Production of lactic acid by lactic bacteria strains reduces the pH and increases its acidity39.
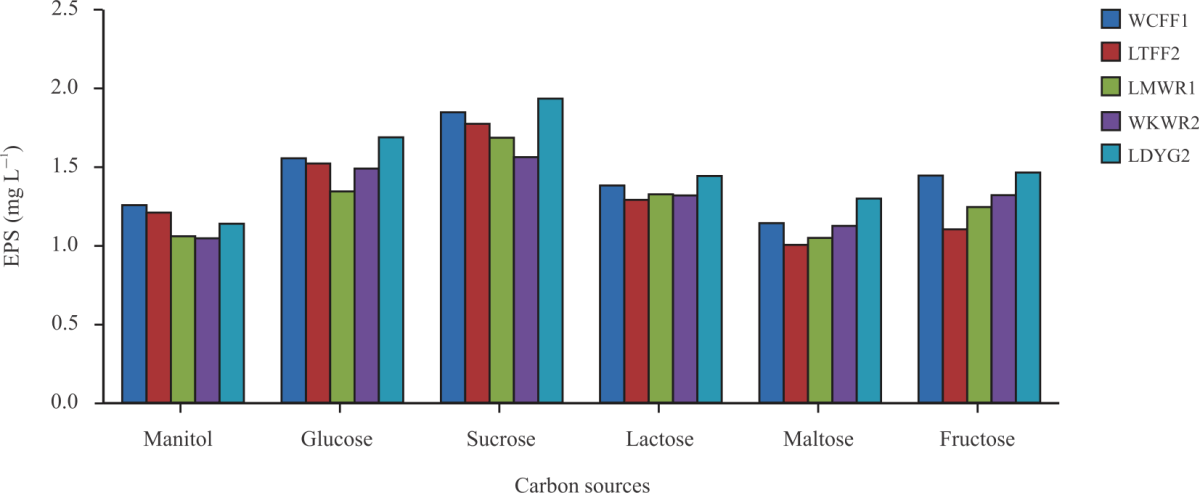 |
| Fig. 5: | Effect of Carbon sources on EPS production by the selected LAB strains |
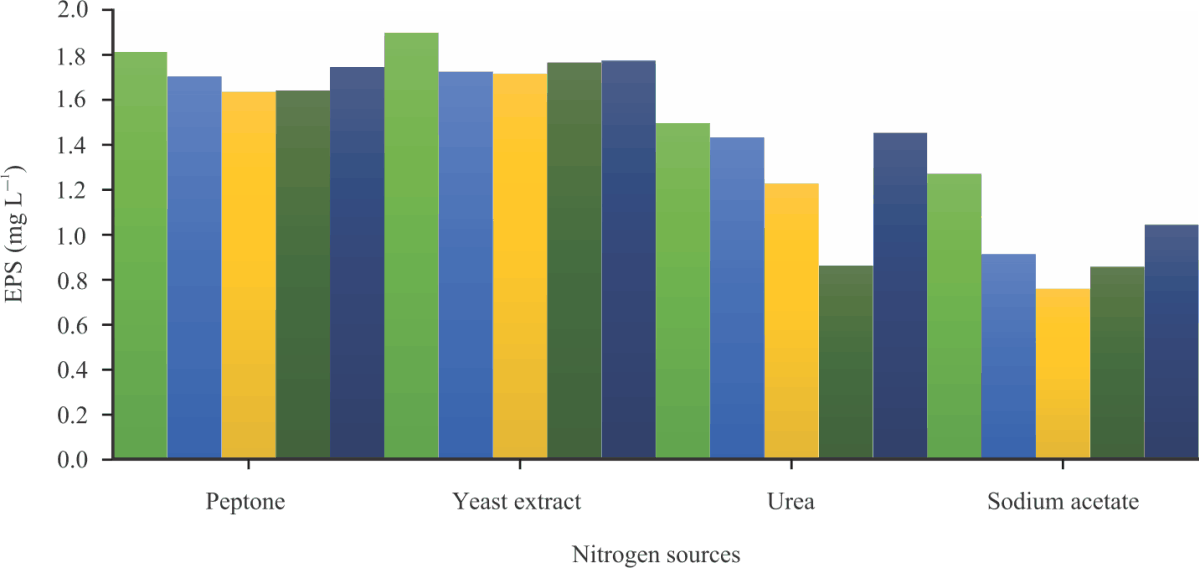 |
| Fig. 6: | Effect of Nitrogen sources on EPS production by the selected LAB strains |
The effect of different carbon sources on EPS production by the selected LAB strains is shown in Fig. 5. Generally, W. confusa FASHADFF1 and L. delbrueckii FASHADYG2 utilizes glucose and sucrose (1684 and 1774 mg L–1) followed by lactose and fructose for high EPS yield. The least EPS production was observed when mannitol and maltose were used as carbon sources. Generally, Leu. mesenteroides FASHADWR1 (1004 mg L–1) and Weissella koreensis FASHADFF2 (1047 mg L–1) produced the least quantity of EPS. The highest EPS yield utilizing sucrose as a carbon source could be because sucrose is a disaccharide and it is broken down to glucose and fructose. LAB is also able to efficiently utilize sucrose and glucose than other sugars as an energy source or precursor for the EPS synthesis of this strain40. Carbon is required for growth and EPS production by bacteria. In this study, it was found that all the isolates metabolized all tested sugars. Aslim41 and Lane42 reported that LAB strains can utilize carbohydrate sugars, such as glucose, lactose, sucrose and mannose as the carbon source for EPS production. Zotta et al.43 reported that sucrose has to be one of the most suitable carbon sources for EPS production by LAB including Weissella sp.
The effect of nitrogen sources on EPS production by the LAB is shown in Fig. 6. W. confusa FASHADFF1, L. tucetti FASHADFF2, Leu. mesenteroides FASHADWR1, W. koreensis FASHADWR2 and L. delbrueckii FASHADYG2 had the highest EPS production (1897, 1725, 1765, 1717 and 1774 mg L–1) when yeast extract was used as a nitrogen source. The least EPS (856, 912, 758, 127 and 1043 mg L–1) was produced when Sodium acetate was used. Generally, L. delbrueckii FASHADYG2 and W. confusa FASHADFF1 produced the highest amount of EPS by utilizing all the nitrogen sources used for this research work. Yeast extract, however, supported the highest production of EPS which is in line with the work of Razack et al.44 in which yeast extract gave the highest EPS production by Bacillus. Himanshu et al.45 reported that ratios of carbon and nitrogen sources play the most important role in cellular growth and exo-biopolymer production.
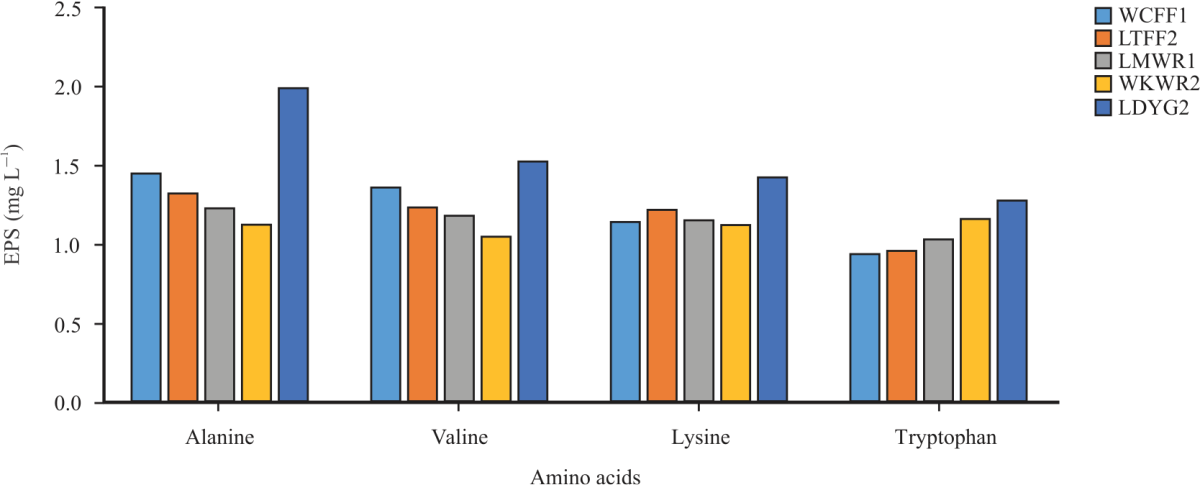 |
| Fig. 7: | Effect of amino acid on EPS production by the selected LAB strains |
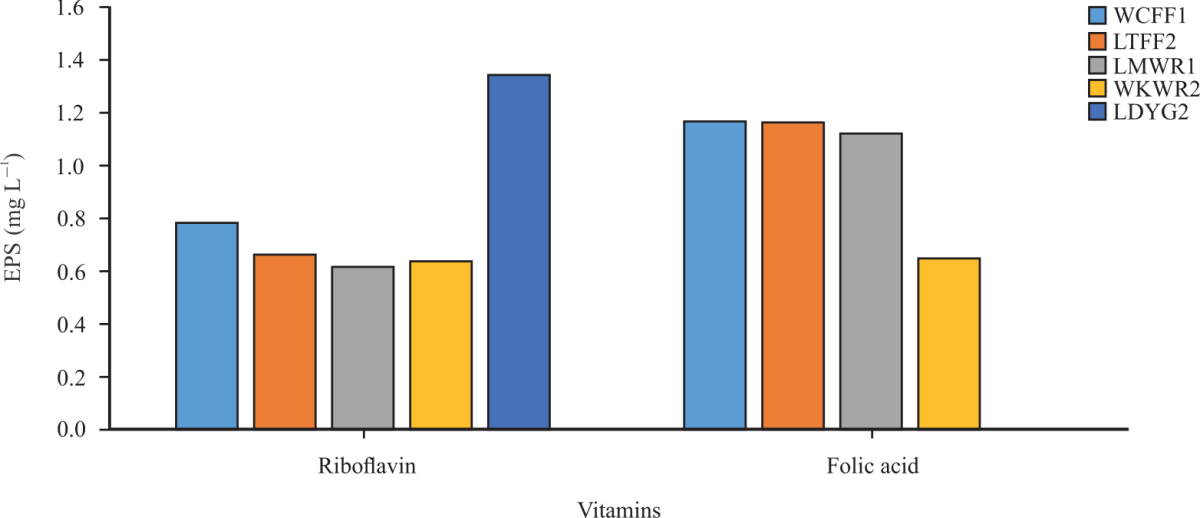 |
| Fig. 8: | Effect of vitamins on EPS production by the selected LAB strains |
The effect of different amino acids on EPS production by the LABs is shown in Fig. 7. W. confusa FASHADFF1, L. tucetti FASHADFF2 and Leu. mesenteroides FASHADWR1 had the highest EPS production (1416, 1292 and 1199 mg L–1) when alanine was used as a source of amino acid with L. delbrueckii FASHADYG2 having the highest EPS production (1946 mg L–1). W. koreensis FASHADWR2 had the highest EPS production (1133 mg L–1) when tryptophan was used and the least EPS production (1023 mg L–1) was observed when valine was utilized. The least production (1248 mg L–1) was recorded when tryptophan was utilized. However, alanine supported the highest amount of EPS production while lysine had the least EPS production. This could be a result of alanine being an essential nutrient for the growth and production of EPS. Othman et al.46 and Zhongchun et al.47 reported that Lactobacillus sp. consumes more of alanine, then lysine, valine, tryptophan, aspartic acid, etc. Studies have shown that concentrations of most of the amino acids slightly increase due to fermentation. There is evidence that these amino acids have unique roles in human metabolism. In addition to providing substrates for protein synthesis, suppressing protein catabolism and serving as substrates for gluconeogenesis, they also trigger muscle protein synthesis and promote protein synthesis48.
The effect of vitamins on EPS production by the LAB strains is shown in Fig. 8. Folic acid supported the highest EPS production by three LAB strains (W. confusa FASHADFF1, L. tucetti FASHADFF2, Leu. mesenteroides FASHADWR1). Riboflavin supported EPS production by two (W. koreensis FASHADWR2 and L. delbrueckii FASHADYG2). The ability of folic acid to support EPS production by L. delbrueckii FASHADYG2 (LDYG2) maybe because Lactobacillus spp can utilize folate. As suggested by Wouters et al.49, the amount of folate may be increased depending on the starter cultures. Among LAB, many Lactobacillus spp. and Lactococci spp. including L. plantarum, L. bulgaricus, Lc. lactis, Streptococcus thermophilus and Enterococcus spp. can utilize folate. On the other hand, some Lactobacilli (L. gasseri, L. salivarius, L. acidophilus and L. johnsonii) used as both starter cultures and probiotics, cannot utilize folate because they lack few specific genes involved in folate biosynthesis50. The ability of Riboflavin to support the production of EPS may because riboflavin is a component of flavin coenzymes which appears to be essential for the growth of lactic acid bacteria51. Although diverse LAB EPS have been studied, little information is available regarding their physicochemical parameters. Factors such as medium composition and fermentation conditions (pH and temperature) can affect EPS production. This study describes some molecularly characterized microbial EPS producers and fermentation parameters necessary to enhance EPS yield in the chemical, food and pharmaceutical industries. Exopolysaccharides produced by lactic acid bacteria are natural and nontoxic thereby highly recommended for use as a bio-thickener. However, the production of EPS from Modified Exopolysaccharide Selection Medium (mESM) is tedious, expensive and time-consuming. Although milk is relevant in most food industries, other mediums that are cheap, readily available with the ability to support the growth and EPS production in industries should be studied.
CONCLUSION
The LAB isolated from fermented foods were molecularly identified as Leuconostoc mesenteroides FASHADWR1, Weissella confusa FASHADFF1, Lactobacillus tucceti FASHADFF2, Weissella koreensis FASHADWR2 and Lactobacillus delbrueckii FASHADYG2. This study revealed that the Physico-chemical parameters and their optimization greatly influence the production of EPS when compared with the number of EPS produced before optimization. From the findings, it may be suggested that sucrose, yeast extract, alanine and riboflavin are the best carbon, nitrogen, amino acid and vitamin sources for EPS production. The best pH was 5 while 35°C was the best temperature for maximum EPS production.
SIGNIFICANCE STATEMENT
This study discovers the optimization of exopolysaccharides produced by some lactic acid bacteria that can be beneficial as a bio-thickener and viscosifier in most food, pharmaceutical and chemical industries. This study will help the researcher to uncover the critical areas of the optimum temperature, pH and the best carbon, nitrogen, amino acid and vitamin sources for higher EPS yield that many researchers were not able to explore. Thus, a new theory on exopolysaccharide production obtained from lactic acid bacteria using other sources for optimization may be arrived at.
ACKNOWLEDGMENT
Authors are grateful to the University of Ibadan, Oyo State and Ajayi Crowther University, Oyo Town, Oyo State, Nigeria.
REFERENCES
- Imran, M.Y.M., N. Reehana, K.A. Jayaraj, A.A.P. Ahamed and D. Dhanasekaran et al., 2016. Statistical optimization of exopolysaccharide production by Lactobacillus plantarum NTMI05 and NTMI20. Int. J. Biol. Macromol., 93: 731-745.
CrossRefDirect Link - Adebayo-Tayo, B., R. Ishola and T. Oyewunmi, 2018. Characterization, antioxidant and immunomodulatory potential on exopolysaccharide produced by wild type and mutant weissella confusa strains. Biotechnol. Rep.
CrossRefDirect Link - Laws, A., Y. Gu and V. Marshall, 2001. Biosynthesis, characterisation and design of bacterial exopolysaccharides from lactic acid bacteria. Biotechnol. Adv., 19: 597-625.
CrossRefDirect Link - Liu, Q., X. Huang, D. Yang, T. Si, S. Pan and F. Yang, 2016. Yield improvement of exopolysaccharides by screening of the Lactobacillus acidophilus ATCC and optimization of the fermentation and extraction conditions. EXCLI J., 15: 119-133.
CrossRefDirect Link - Broadbent, J.R., D.J. McMahon, D.L. Welker, C.J. Oberg and S. Moineau, 2003. Biochemistry, genetics and applications of exopolysaccharide production in Streptococcus thermophilus: A review. J. Dairy Sci., 86: 407-423.
CrossRefDirect Link - Mano, J.F., G.A. Silva, H.S. Azevedo, P.B. Malafaya and R.A. Sousa et al., 2007. Natural origin biodegradable systems in tissue engineering and regenerative medicine: Present status and some moving trends. J. Roy. Soc. Inter., 4: 999-1030.
CrossRefDirect Link - Kadajji, V.G. and G.V. Betageri, 2011. Water soluble polymers for pharmaceutical applications. Polymers, 3: 1972-2009.
CrossRefDirect Link - Weiss, J. and G. Muschiolik, 2006. Factors affecting the droplet size of water-in-oil emulsion (W/O) and the oil globule size in water-in-oil-in-water emulsions (W/O/W). J. Disper. Sci. Technol., 28: 703-716.
CrossRefDirect Link - Aguirre, M. and M.D. Collins, 1993. Lactic acid bacteria and human clinical infection. J. Applied Bacteriol., 75: 95-107.
Direct Link - Suryawanshi, N., S. Naik and J.S. Eswari, 2019. Extraction and optimization of exopolysaccharide from Lactobacillus sp. using response surface methodology and artificial neural networks. Prep. Biochem. Biotechnol., 49: 987-996.
CrossRefDirect Link - Rattanachaikunsopon, P. and P. Phumkhachorn, 2010. Lactic acid bacteria: Their antimicrobial compounds and their uses in food production. Ann. Boil. Res., 1: 218-228.
Direct Link - Mozzi, F., G.S. de Giori and G.F. de Valdez, 2003. UDP-galactose 4-epimerase: A key enzyme in exopolysaccharide formation by Lactobacillus casei CRL 87 in controlled pH batch cultures. J. Applied Microbiol., 94: 175-183.
CrossRefDirect Link - Patel, A. and J.B. Prajapati, 2013. Food and health applications of exopolysaccharides produced by lactic acid bacteria. Adv. Dairy Res., Vol. 1.
CrossRefDirect Link - Benattouchea, Z., D. Bouhadib and G.B. Rahoc, 2018. Antioxidant and antibacterial activities of exopolysaccharides produced by lactic acid bacteria isolated from yogurt. Int. J. Foods Stud., 7: 30-37.
CrossRefDirect Link - Mozzi, F., F. Vaningelgem, E.M. Hebert, R. van der Meulen, M.R.F. Moreno, G.F. de Valdez and L. de Vuyst, 2006. Diversity of heteropolysaccharide-producing lactic acid bacterium strains and their biopolymers. Applied Environ. Microbiol., 72: 4431-4435.
CrossRefDirect Link - Ayala-Hernández, I., A.N. Hassan, H.D. Goff and M. Corredig, 2009. Effect of protein supplementation on the rheological characteristics of milk permeates fermented with exopolysaccharide-producing Lactococcus lactis subsp. cremoris. Food Hydrocol., 23: 1299-1304.
CrossRefDirect Link - Girard, M. and C. Schaffer-Lequart, 2007. Gelation and resistance to shearing of fermented milk: Role of exopolysaccharides. Int. Dairy J., 17: 666-673.
CrossRefDirect Link - Zajšek, K., A. Goršek and M. Kolar, 2013. Cultivating conditions effects on kefiran production by the mixed culture of lactic acid bacteria imbedded within kefir grains. Food Chem., 139: 970-977.
CrossRefDirect Link - Benhadria, M.K., M.A.T. Touil and B. Meddah, 2017. Optimization of production of microbial exopolysaccharides (EPS) with essential oils from two medicinal plants. J. Appl. Biosci., 111: 10925-10933.
CrossRefDirect Link - Behare, P.V., R. Singh, R. Nagpal and K.H. Rao, 2013. Exopolysaccharides producing Lactobacillus fermentum strain for enhancing rheological and sensory attributes of low-fat dahi. J. Food Sci. Technol., 50: 1228-1232.
CrossRefDirect Link - Ishola, R. and B. Adebayo-Tayo, 2018. Mutagenesis and immobilization effect on exopolysaccharide production by weissella confusa and Lactobacillus delbrueckii. J. Adv. Microbiol., 10: 1-10.
CrossRefDirect Link - De Man, J.C., M. Rogosa and M.E. Sharpe, 1960. A medium for the cultivation of Lactobacilli. J. Applied Bacteriol., 23: 130-135.
CrossRefDirect Link - Gevers, D., G. Huys and J. Swings, 2001. Applicability of rep-PCR fingerprinting for identification of Lactobacillus spieces. FEMS Microbiol. Lett., 205: 31-36.
CrossRef - Angelov, A.I., G. Petrova, A.D. Angelov, P. Stefanova and I.Y. Bokossa et al., 2017. Molecular identification of yeasts and lactic acid bacteria involved in the production of beninese fermented food degue. The Open Biotechnol. J., 11: 94-104.
CrossRefDirect Link - Verdern Berg, D.J.C., G.W. Obijinn, A.C. Janssen, M.L. Giuseppin and R. Vreeker et al., 1995. Production of a novel extracellular polysaccharides by Lactobacillus sakei 0-1 and characterization of polysaccharides. Applied Environ. Microbiol., 61: 2840-2844.
Direct Link - Smitinont, T., C. Tansakul, S. Tanasupawat, S. Keeratipibul and L. Navarini et al., 1999. Exopolysaccharide-producing lactic acid bacteria strains from traditional Thai fermented foods: Isolation, identification and exopolysaccharide characterization. Int. J. Food. Microbiol., 51: 105-111.
CrossRef - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Jiang, J., S. Guo, W. Ping, D. Zhao and J. Ge, 2020. Optimization production of exopolysaccharide from Leuconostoc lactis L2 and its partial characterization. Int. J. Bio. Macromol., 159: 630-639.
CrossRefDirect Link - Blaiotta, G., C. Pennacchia, D. Ercolini, G. Moschetti and F. Villani, 2003. Combining denaturing gradient gel electrophoresis of 16S rDNA V3 region and 16S–23S rDNA spacer region polymorphism analyses for the identification of staphylococci from Italian fermented sausages. Syst. Appl. Microbiol., 26: 423-433.
CrossRefDirect Link - Basaran, P., N. Basaran and I. Cakir, 2001. Molecular differentiation of Lactococcus lactis subspecies lactis and cremoris strains by ribotyping and site-specific PCR. Curr. Microbiol., 42: 45-48.
CrossRef - Adiguzel, G. and M. Atasever, 2009. Phenotypic and genotypic characterization of lactic acid bacteria isolated from Turkish dry fermented sausage. Rom. Biotechnol. Let., 14: 4130-4138.
Direct Link - Caro, I., G. Bécares, L. Fuentes, M.R. Garcia-Armesto and J. Rúa et al., 2015. Evaluation of three PCR primers based on the16S rRNAgene for the identification of lactic acid bacteria from dairy origin. CyTA - J. Food, 13: 181-187.
CrossRefDirect Link - Mostefaoui, A., A. Hakem, B. Yabrir, S. Boutaiba and A. Badis, 2014. Screening for exopolysaccharide-producing strains of thermophilic lactic acid bacteria isolated from Algerian raw camel milk. Afr. J. Microbiol. Res., 8: 2208-2214.
CrossRefDirect Link - Joshi, S.R. and K. Koijam, 2014. Exopolysaccharide production by a lactic acid bacteria, Leuconostoc lactis isolated from ethnically fermented beverage. Natl. Acad. Sci. Lett., 37: 59-64.
CrossRefDirect Link - Gamar, L., K. Blondeau and J.M. Simonet, 1997. Physiological approach to extracellular polysaccharide production by Lactobacillus rhamnosus strain C83. J. Applied Microbiol., 83: 281-287.
CrossRefDirect Link - Prasanna, P.H.P., A.S. Grandison and D. Charalampopoulos, 2012. Effect of dairy-based protein sources and temperature on growth, acidification and exopolysaccharide production of Bifidobacterium strains in skim milk. Food Res. Int., 47: 6-12.
CrossRefDirect Link - De Vuyst, L. and B. Degeest, 1999. Heteropolysaccharides from lactic acid bacteria. FEMS Microbiol. Rev., 23: 153-177.
CrossRefPubMedDirect Link - Elliason, D.J. and S.R. Tatini, 1999. Enhanced inactivation of Salmonella Typhimurium and verotoxigenic Escherichia coli by Nisin at 6-5°C. Food Microbiol., 16: 257-267.
CrossRefDirect Link - Soltani, M., N. Guzeler and A. Hayaloglu, 2015. The influence of salt concentration on the chemical, ripening and sensory characteristics of Iranian white cheese manufactured by UF-treated milk. J. Dairy Res., 82: 365-374.
CrossRefDirect Link - Looijesteijn, P.J., L. Trapet, E. de Vries, T. Abee and J. Hugenholtz, 2001. Physiological function of exopolysaccharides produced by Lactococcus lactis. Int. J. Food Microbiol., 64: 71-80.
CrossRefDirect Link - Aslim, B., Z.N. Yuksekdag, E. Sarikaya and Y. Beyatli, 2005. Determination of the bacteriocin-like substances produced by some lactic acid bacteria isolated from Turkish dairy products. LWT-Food Sci. Technol., 38: 691-694.
CrossRefDirect Link - Lane, D.J., 1991. 16S/23S rRNA Sequencing. In: Nucleic Acid Techniques in Bacterial Systematic, Stackebrandt, E. and M. Goodfellow (Eds.). John Wiley and Sons, New York, USA., ISBN-13: 9780471929062, pp: 115-175.
Direct Link - Zotta, T., P. Piraino, E. Parente, G. Salzano and A. Ricciardi, 2008. Characterization of lactic acid bacteria isolated from sourdoughs for Cornetto, a traditional bread produced in Basilicata (Southern Italy). World J. Microbiol. Biotechnol., 24: 1785-1795.
CrossRefDirect Link - Razack, S.A., V. Velayutham and V. Thangavelu, 2013. Medium optimization for the production of exopolysaccharide by Bacillus subtilis using synthetic sources and agro wastes. Turk. J. Biol., 37: 280-288.
CrossRefDirect Link - Kishk, Y.F.M. and H.M.A. Al-Sayed, 2007. Free-radical scavenging and antioxidative activities of some polysaccharides in emulsions. LWT-Food Sci. Technol. , 40: 270-277.
CrossRefDirect Link - Nor, Z.O., H.A. El-Enshasy, A.M. Roslinda, M.R. Sarmidi and A.A. Ramlan, 2009. Kinetics of cell growth and functional characterization of probiotic strains Lactobacillus delbrueckii and Lactobacillus paracasei isolated from breast milk. Deutsche Lebensmittel-Rundschau, 105: 444-450.
Direct Link - Zhongchun, T., Z. Loudan, L. Junqi, J. Yutao, H. Lijia and D. Dongmei, 2014. An in vitro study on the effect of free amino acidsalone or combination withnisin on biofilms as well as on planktonic bacteria of Streptococus mutans. PLoS ONE, Vol. 9.
CrossRefDirect Link - Haug, A., A.T. Hostmark and O.M. Harstad, 2007. Bovine milk in human nutrition: A review. Lipids Health Dis., 6: 25-25.
PubMedDirect Link - Wouters, J.T.M., E.H.E. Ayad, J. Hugenholtz and G. Smith, 2002. Microbes from raw milk for fermented dairy products. Int. Dairy J., 12: 91-109.
CrossRefDirect Link - Guldfeldt, L.U., K.I. Sorensen, P. Stroman, H. Behrndt and D. Williams et al., 2001. Effect of starter cultures with a genetically modified peptidolytic or lytic system on Cheddar cheese ripening. Intl. Dairy J., 11: 373-382.
CrossRefDirect Link - Cocaign-Bousquet, M., C. Garrigues, L. Novak, N.D. Lindley and P. Loubiere, 1994. Rational development of a simple synthetic medium for the sustained growth of Lactococcus lactis. J. Applied. Bacteriol., 79: 108-116.
CrossRefDirect Link