
Ekundayo Temitope Cyrus
Department of Microbiology, Federal University of Technology, P.M.B 704, Akure, Nigeria
Arotupin Daniel Juwon
Department of Microbiology, Federal University of Technology, P.M.B 704, Akure, Nigeria
Microbiology Journal
Year: 2016 | Volume: 6 | Issue: 1-2 | Page No.: 25-33







ABSTRACT
Background and Objective: Aflatoxins production has limited the use of many strains of Aspergillus flavus in food fermentation and enzyme technology, despite their excellent abilities for copious enzyme production. In a bid to overcome this problem, the effects of some selected mutagenic agents on expression of aflatoxigenic traits in A. flavus was investigated in order to enhance its potential biotechnological uses. Materials and Methods: Spores of the strain were treated with chemical mutagens which include nitrous acid, sodium azide, ethyl methane sulphonate and hydroxylamine and ultraviolet radiation for 5400 sec. Mutants obtained at every 900 sec successively were stored under protection away from light for 24 h in order to stabilize DNA dimmers and the mutants were grown in aflatoxin production medium at 30°C for 10 days. Aflatoxins produced were quantified using enzyme-linked immunosorbent assay. Data obtained were subjected to analysis of variance using Statistical Package for Social Science (SPSS) version 17 at p≤0.05. Results: Ultraviolet treatment achieved 93.11% reduction of aflatoxins expression in the mutant CTUV–75 (103.31±0.44 ppt) compared to wild strain. However, hydroxylamine showed 89.40% expression of aflatoxigenic traits in the CTNH2OH–60 compared to the parent. Conclusion: Ultraviolet radiation caused considerable repression (93.11%) of aflatoxin production in the A. flavus strain in contrast to other mutagens. Nevertheless, the equivalent concentration of aflatoxin produced still exceeded 50 ppt limits set by FAO/WHO/EU. Thus, the use of such mutant in fermentation could pose public health risk. Therefore, it is suggested that further trials of UV treatment or other molecular methods may be employed whether aflatoxins production could be reduced below the standard limit or entirely get rid off in the mutant.
PDF Abstract XML References Citation
Received: November 17, 2015;
Accepted: January 28, 2016;
Published: March 15, 2016
How to cite this article
Ekundayo Temitope Cyrus and Arotupin Daniel Juwon, 2016. Repression of Aflatoxigenic Traits in Aspergillus flavus SWtS01 Through Random Mutagenesis. Microbiology Journal, 6: 25-33.
DOI: 10.3923/mj.2016.25.33
URL: https://scialert.net/abstract/?doi=mj.2016.25.33
DOI: 10.3923/mj.2016.25.33
URL: https://scialert.net/abstract/?doi=mj.2016.25.33
INTRODUCTION
The genus Aspergillus has revolutionized biotechnological and industrial practices in term of food production, fermentation, bio-fuel and bio-solvent production in recent years. The most widely used bio-tools in industries are fungi and several enzymes, in which Aspergillus is among the most economically important fungal genera, owning to its natural ability to produce diverse arrays of metabolites, enzymes, surfactants, antimicrobials and organic acids for instance, Aspergillus species (A. oryzae, A. sojae, A. awamori and A. saitoi) are of great practical importance in fermentation industries, enzyme technologies, food industries and civilization in Japan (Ichishima, 2011). Aspergillus species have been documented in fermentation industry for the production of various enzymes, organic acids and pharmaceutically important compounds such as lovastatin (a potent cholesterol-lowering drug), antibiotics, antifungals and anti-tumor drugs (terrequinone A) and in oriental food fermentations (Hoffmeister and Keller, 2007; Varga and Samson, 2008).
However, with this invaluable industrial and biotechnological significance of Aspergillus, there are associated potential hazards with certain strains. These strains produce a range of secondary metabolites known as mycotoxins, which are harmful to humans and animals (Varga and Samson, 2008) under suitable conditions. Frisvad et al. (2009) reported that atleast 226 potentially bioactive extrolites and metabolites, including acids and proteins have been produced from A. fumigatus. The number of biosynthetic families of secondary metabolites reported to be produced by A. fumigatus is impressive and so is the number of individual extrolites.
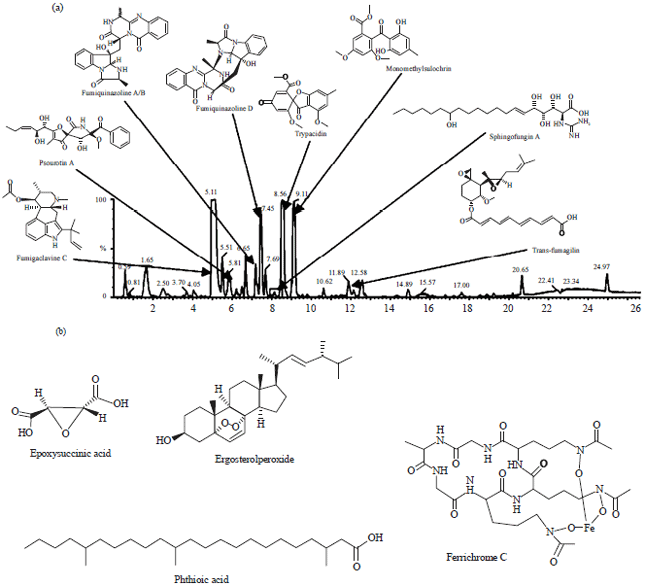  | |
| Fig. 1(a-b): | Representatives of single extrolites produced by Aspergillus fumigatus (Frisvad et al., 2009) |
Figure 1 shows that the representatives of the most toxic or otherwise bioactive major metabolites from each biosynthetic family, namely fumigatins, fumigaclavines, fumiquinazolines, trypacidin, monomethylsulochrin, fumagillins, gliotoxins, pseurotins, chloroanthraquinones, fumitremorgins, verruculogen, helvolic acids and pyripyropenes (Frisvad et al., 2009). These have limited the uses of many strains of Aspergillus in industrial practices.
On the other hand, Aspergillus flavus are capable of producing aflatoxin. Aflatoxins (AFs) B1, B2, G1, G2 (AFB1, AFB2, AFG1 and AFG2) are known group of toxic, carcinogenic and teratogenic polypeptide noted as health hazards in humans and livestock and as threat to agricultural productivity (Pitt and Hocking, 1999). The metabolic products of these aflatoxins, AFM1 and AFM2 are also relevant as direct contaminants in foods and feeds especially milks. Chemical structures of aflatoxin B1, B2, G1, G2, M1 an M2 are shown in Fig. 2 (Frisvad et al., 2009). Certain strains of A. flavus are sometimes prodigal producers of value-added products and serve as better bio-tools. This necessitates the need to suppress undesirable traits and simultaneously enhance the anticipated ones. One of the various approaches in achieving such goal is the mutagenesis directed toward exerting changes in the genetic regulation of aflatoxin biosynthesis. Therefore, the present investigation was a quest into the ability of some mutagens to impact control of aflatoxin biosynthesis through random mutation of aflatoxigenic A. flavus SWtS01.
MATERIALS AND METHODS
Aflatoxigenic fungus: The test fungus, Aspergillus flavus SWtS01 was previously isolated from sawdust and identified according to Barnett and Hunter (1972).
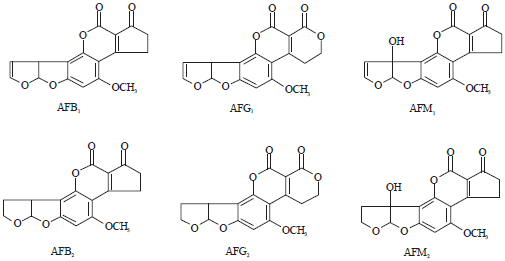 | |
| Fig. 2: | Chemical structures of aflatoxin B1, B2, G1, G2, M1and M2 (Frisvad et al., 2009) |
It was thereafter confirmed to be active producer of cellulase and aflatoxin in the Microbiology Research Laboratory of the Federal University of Technology Akure, Nigeria.
Preparation of conidial suspension: A spore suspension of the isolate was prepared by serial dilution method from 5 days old culture according to Elliaiah et al. (2002). This was prepared by the addition of sterilized phosphate buffer (pH 7.0) into the 5 days old slant culture. The spores were dislodged from the mycelia with the aid of a sterile inoculating loop and thoroughly vortexed. The inoculum size was then measured with a haemocytometer as described by Sharma (1989).
Ethyl Methane Sulphonate (EMS) mutagenesis of Aspergillus flavus SWtS01: Ethyl methane sulphonate mutant strains of Aspergillus flavus SWtS01 were generated according to Nadeem et al. (2010). About 6.0 mg stock solution of EMS was prepared. Volumes of 2.0 mL of spore suspension (108) was taken and incubated with equivalent amount of EMS stock solution for 15, 30, 45, 60, 75 and 90 min at 28±2°C. After the period of treatment, spores were centrifuged and washed with sterilized phosphate buffer (pH 7.0) twice at 6000 rpm for 15 min at 4°C. A volume of 0.1 mL of EMS-treated fungal suspension was plated onto potato dextrose agar at 28±2°C for 5 days.
Nitrous acid mutagenesis of Aspergillus flavus SWtS01: Nitrous acid-mutant strains were generated according to the modified method of Iftikhar et al. (2010). Spore culture of Aspergillus flavus SWtS01 prepared in Vogel’s medium was subjected to nitrous acid (0.1 M sodium nitrite in phosphate buffer, pH 5.0) treatment at interval of 15, 30, 45, 60, 75 and 90 min by incubating the mixture at 28±2°C. Treated spores were washed thrice at 6000 rpm for 15 min to remove traces of mutagen. After reconstituting the spores in saline, 0.1 mL was plated onto potato dextrose agar at 28±2°C for 5 days.
Sodium azide mutagenesis of Aspergillus flavus SWtS01: The sodium azide mutagenesis of Aspergillus flavus SWtS01 was conducted according to Rajeshkumar and Ilyas (2011). To 9.0 mL of the spore suspension, 1.0 mL sterile solution of sodium azide (400 μg mL–1 in phosphate buffer) was added. The reaction was allowed to proceed at room temperature. Control tube was also kept without sodium azide. About 1.0 mL of the spore solution was withdrawn from the reaction mixture at 15, 30, 45, 60, 75 and 90 min. This was centrifuged at 6000 rpm for 15 min to remove any trace of the mutagen. The spores were washed three times with sterile distilled water and again re-suspended in 10.0 mL sterile buffer. About 0.1 mL of the treated spore suspension was plated onto a potato dextrose agar at 28±2°C for 5 days.
Hydroxylamine mutagenesis of Aspergillus flavus SWtS01: Hydroxylamine-mutant strains of Aspergillus flavus SWtS01 were obtained according to the modified procedure of Damisa et al. (2013). One molar stock solution of hydroxylamine was prepared by reacting 1 M hydroxylammonium chloride with 1 M NaOH solution. About 100 mM solution of the hydroxylamine was there from prepared using phosphate buffer (pH 7.5).
To 9.0 mL of spore suspension in phosphate buffer, a 1.0 mL sterile solution of the hydroxylamine (100 mM) was added. The reaction was allowed to proceed at 28±2°C. A control was set up without the mutagen. A sample of 1.0 mL of the spore solution was withdrawn from the reaction mixture at 15, 30, 45, 60, 75 and 90 min. After the period of treatment, spores were centrifuged at 6000 rpm and washed with sterilized phosphate buffer (pH 7.0) twice for 15 min (Rajeshkumar and Ilyas, 2011). About 0.1 mL of the treated spore suspension was thereafter plated onto a potato dextrose agar at 28±2°C for 5 days.
Ultraviolet (UV) radiation mutagenesis of Aspergillus flavus SWtS01: Ultraviolet-mutant strains were created according to the method of Irfan et al. (2011). About 2.0 mL quantity of the spore suspension each was aseptically pipetted into 8 sterile petri dishes. The plates were exposed to UV radiation (366 nm) at a distance of 13.0 cm away from the centre of a germicidal lamp (Model UVL-21, San Gabriel, Calif, USA) for 15, 25, 35, 45, 55, 65, 75 and 85 min, respectively. Constant agitation of the suspension was achieved by intermittent rotation during the treatment (Elliaiah et al., 2002). The UV exposed spore suspension-containing plates were stored overnight in dark to avoid photo-reactivation and for thymine-thymine (T-T) dimmers stabilization (Irfan et al., 2011). After overnight incubation, the irradiated spore suspensions were serially diluted in phosphate buffer (pH 7.0). Every diluted spore suspension (100 μL) was plated onto potato dextrose agar at 28±2°C for 5 days to recovered mutants.
Screening of Aspergillus flavus SWtS01 mutant strains for aflatoxins production: Aflatoxin production by the mutants was assessed according to the method of Zaini et al. (2009). Ten mycelial plugs of 10 mm diameter of a 5 days old culture of the mutants were harvested. This was inoculated into a 250 mL Erlenmeyer flasks containing 50 mL yeast extract-sucrose (YES) broth medium (2% yeast extract, 20% sucrose and pH 5.5). The flask was incubated at 30°C for 10 days in the dark. Afterwards, the culture was filtered and centrifuged at 10,000 rpm for 5 min.
ELISA-based analysis of total aflatoxins produced by Aspergillus flavus SWtS01 mutants: Total aflatoxin production was evaluated by using Ridascreen Aflatoxin Total Kit (R-Biopharm AG, Darmstadt, Germany). The reagents and materials provided with the kit include microtiter plate (12×8 strips) coated with captured antibodies, calibrators/standards (6): 0, 50, 150, 450, 13500 and 4050 pg mL–1 (ppt) of aflatoxin B (1 mL) each, methanol-water and aflatoxin conjugate (6 mL) (peroxidase conjugated aflatoxin B). Others include anti-aflatoxin antibodies (6 mL) (monoclonal), substrate-chromogen solution (10 mL) (stained red because of tetramethyl benzidin), stop solution (14 mL) that contained 1 M sulphuric acid and washing buffer salt (0.05% tween 20). Reagents and materials were stored at 4-8°C.
The assay was done according to the procedure provided with the kit (R-Biopharm AG, Darmstadt, Germany). About 1 mL of the sample supernatant prepared from the culture filtrate was pipetted into a centrifuge tube. This was additionally spun at 14,000 rpm for 5 min with MiniSpin centrifuge (Eppendorf AG, model 2231, Hamburg Germany). About 100 μL of the standard solution or sample (as the case may be) was pipetted into pre-labelled wells of the test strips using a multi-channel pipette. The standard solution and sample positions were recorded accordingly. About 100 μL of the conjugate enzyme and 100 μL of the antibody solution was thereafter added into the entire wells successively with the multi-channel pipette. The plate was gently rocked by manual shaking and incubated in dark at 23±2°C for 30 min. Afterward, contents of the wells were poured out. The microwell holder was tapped upside down vigorously (three times in a row) against a stack of absorbent paper towels to ensured complete removal of liquid from the wells. Each well was subsequently washed three times with 250 μL diluted washing buffer solution in every washing step. The remaining buffer in the wells was removed by patting the plate dry on a stack of paper towels. About 100 μL of substrate solution was added to all the wells and gently mixed by manual shaking. The strip was finally incubated in dark at 23±2°C for 15 min. The reaction was afterwards terminated by the addition of 100 μL stop solution to each well in the same sequence the substrate was added. The strip was gently rocked as soon as the substrate was added. The absorbance was read at 450 nm with a microplate ELISA photometer (Model ELx800, BioTek, USA) within 30 min after the addition of stopping solution (R-Biopharm AG, Darmstadt, Germany). Aflatoxin production was compared between the wild strain and the mutants as Relative Aflatoxin Production (RAP) and as Difference in Aflatoxin Production (DAP) as described by Cyrus and Juwon (2015):
Where:
| AFm | = | Aflatoxin produced by the mutant |
| AFw | = | Aflatoxin produced by the wild strain |
Where:
| RAPm | = | Relative aflatoxin production of the mutant |
| RAPw | = | Relative aflatoxin production of the wild strain |
Statistical analysis: Data obtained were subjected to analysis of variance (ANOVA) of SPSS software version 17 (Microsoft Corporation, USA) at 95% confidence interval. Data presented on the average of three replicates (Mean±SE) were obtained from duplicate determinations. Means were separated using Duncan’s new multiple range test and differences were considered significant at p≤0.05.
RESULTS
Aflatoxin biosynthesis potential of chemical mutant strains of A. flavus SWtS01: Table 1 shows aflatoxin biosynthesis by EMS-mutant strains of A. flavus SWtS01. Aflatoxin production declined significantly among the mutant strains (83.33%) while high yield was obtained in 16.67% of the mutants. The highest synthesis was recorded in CTEMS15 (1836.00±1.00 ppt), a 17.92% upturn in production while the least was 353.50±1.50 ppt in CTEMS75, 77.30% cutback in aflatoxin production. There is no affiliation between reaction time and percentage difference in aflatoxin production (r = 0.141, p = 0.355).
Result of the effect of nitrous oxide treatment on aflatoxin production by A. flavus SWtS01 mutants is portrayed in Table 2. Aflatoxin production received considerable upsurge in CTHNO–30 (2060.50±0.50 ppt) and CTHNO–45 (2903.20±3.00 ppt) which is 32.34% and 86.45% production rise, respectively. Out of the six mutants selected 66.67% attained consequential reduction in aflatoxin yield, ranging from 22.80% CTHNO–60 to 89.69% CTHNO–75 when matched up to production realized by the parent strain. Treatment time and aflatoxin biosynthesis showed moderate pattern or association with aflatoxin production which however was not statistically significant (r = 0.414, p = 0.355).
Effect of sodium azide induced mutation on aflatoxin biosynthesis of A. flavus SWtS01 mutants is showed in Table 3. All the tested sodium azide mutant strains were able to produce aflatoxin in YES medium although with differences in the rate of production (p<0.05). Aflatoxin production ranged from 727.00±2.00 ppt (CTNA45) to 1611.50±1.50 ppt (CTNA15). An 83.33% of the mutants exhibited reduce aflatoxin yield when compared with parent strain (1557.00±2.00 ppt). A 53.31% reduction in aflatoxin biosynthesis was achieved by mutant CTNA45 obtained at 45 min exposure time to mutagenic treatment whereas 3.50% increase was obtained at 15 min. Also, there was no correlation between aflatoxin synthesis and the treatment time r = 0.532, p = 0.05).
| Table 1: | Aflatoxins production by EMS-mutants of A. flavus SWtS01 in YES medium |
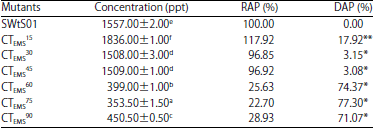 | |
Values are presented as Mean±SE (n = 2), Means with the same superscript letter(s) along the same column are not significantly different (p<0.05), SWtS01: Wild type, 15, 30,...,75 and 90: Treatment duration (min), RAP: Relative aflatoxin production, DAP: Difference in aflatoxin production of mutant and the wild, **Increase and *Decrease | |
| Table 2: | Aflatoxins production by nitrous acid mutants of A. flavus SWtS01 in YES medium |
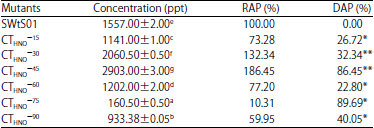 | |
Values are presented as Mean±SE (n = 2), Means with the same superscript letter(s) along the same column are not significantly different (p<0.05) SWtS01: Wild type, 15, 30,...,75 and 90: Treatment duration (min), RAP: Relative aflatoxin production, DAP: Difference in aflatoxin production of mutant and the wild, **Increase and *Decrease | |
| Table 3: | Aflatoxins production by sodium azide mutants of A. flavus and parent strain in YES medium |
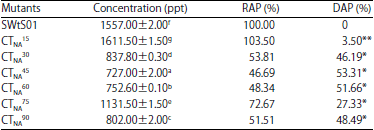 | |
Values are presented as Mean±SE (n = 2), Means with the same superscript letter(s) along the same column are not significantly different (p<0.05), SWtS01: Wild type, 15, 30,...,75 and 90: Treatment duration (min), RAP: Relative aflatoxin production, DAP: Difference in aflatoxin production of mutant and the wild, **Increase and *Decrease | |
Data presented in Table 4 shows the suppression of aflatoxin production by hydroxylamine induced mutation in the isolate. Aflatoxin was significantly different among mutants (p = 0.05). About 16.67% of the hydroxylamine mutants gave higher aflatoxin production than the parent strain (1557.00±2.00 ppt) which was 89.40% production increase in CTNH2H–60. But 75.75% reduction in aflatoxin biosynthesis was attained with CTNH2OH–90 (377.60±0.100 ppt). There was no association between treatment time and corresponding percentage reduction or increase in aflatoxin production (r = 0.181, p = 0.697). TMEMS90 (α subset1 = 0.05).
| Table 4: | Aflatoxins production by hydroxylamine mutants of A. flavus SWtS01 in YES medium |
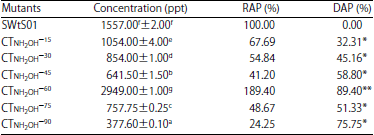 | |
Values are presented as Mean±SE (n = 2), Means with the same superscript letter(s) along the same column are not significantly different (p<0.05), SWtS01: Wild type, 15, 30,...,75 and 90: Treatment duration (min), RAP: Relative aflatoxin production, DAP: Difference in aflatoxin production of mutant and the wild, **Increase and *Decrease | |
| Table 5: | Aflatoxins production by UV-mutants of A. flavus SWtS01 in YES medium |
 | |
Values are presented as Mean±SE (n = 2), Means with the same superscript letter(s), along the same column are not significantly different (p<0.05), SWtS01: Wild type, 15, 30,...,75 and 90: Treatment duration (min), RAP: Relative aflatoxin production, DAP: Difference in aflatoxin production of mutant and the wild, **Increase and *Decrease | |
For hydroxylamine mutants, 50% showed appreciable reduction in aflatoxin biosynthesis but others gave considerable increase (Table 5). The TMNH2OH–90 (266.72±0.027 ppt) and TMNH2OH–15 (1189.50±0.300 ppt) yielded 73.55% best reduction and 17.95% highest increase in aflatoxin level, respectively pivoting on wild strain’s aflatoxin level.
Aflatoxin biosynthesis potential of physical mutant strains of aflatoxigenic A. flavus SWtS01: After effect of UV radiation treatment and subsequent aflatoxin biosynthesis by the resultant mutant strains is depicted in Table 5. Generally, there was substantial reduction in aflatoxin production by 87.50% of the UV-mutant strains of A. flavus SWtS01 contrasted with the control (parent strain). Reduction in aflatoxin production ranged from 0.05% (CTUV–25, 1498.45±3.45 ppt) to 93.11% (CTUV–75, 103.31±0.44 ppt) in comparison with wild strain (SWtS01, 1499.13±0.37 ppt). However, there is a wide dissociation between radiation time and percentage difference in aflatoxin production (r = 0.178, p = 0.647).
DISCUSSION
Aspergillus flavus has being one of the major microorganisms of great industrial importance, but aflatoxin production by some of its strains has limited this potential. Aflatoxigenic Aspergillus flavus was subjected to chemical and physical mutagenic treatments. Different mutagens showed varying effects on aflatoxin production potential of the strain. This could be attributed to differences in mutagenic action of the agents on the fungal strain. For instance, hydroxylamine exercises its mutagenic activities through hydroxylation of DNA cytosine amino group and nitrous acid modifies bases and causes base pair substitution (Hashida and Bisgaard-Frantzen, 2000).
The observed highest reduction in aflatoxin production showed by CTEMS75 EMS mutants of Aspergillus flavus is similar to what was observed in A. parasiticus (Cyrus and Juwon, 2015). Sodium azide mutagenesis achieved 53.31% aflatoxin biosynthesis repression in A. flavus contrary to 46.58% documented in A. parasiticus (Cyrus and Juwon, 2015). Strains differences and mutagenic condition could be attributed to the observations. The level of reduction in aflatoxin synthesis attained with hydroxylamine mutagenesis is very close to the earlier report on A. parasiticus. Ultraviolet rays achieved the highest reduction in aflatoxin yields. This could be attributed to structural modifications in the strain’s DNA that might not fit into the biosynthetic pathway. This result is contrary to the effects of UV in A. parasiticus which led to high level of aflatoxin production (Cyrus and Juwon, 2015). The main effect of ultra violet light is structural modification of pyrimidine (cytosine and thyamine) and dimmerization of thyamine, which distort the structure of DNA heliex and block further replication process (Sambrook and Russell, 2001).
The declined production of aflatoxin obtained in some mutants in this study, could be due to mutation that led to recession of genes that are involved in aflatoxin pathway-specific regulatory gene that either resulted in lessened accumulation of major aflatoxin precursors and vice versa. Chang et al. (2002) as earlier reported some aflatoxin pathway-specific regulatory genes. Also, Flaherty and Payne (1997) reported that mutation could led to overexpression of AFLR in A. flavus, up-regulated aflatoxin pathway gene transcription and aflatoxin accumulation. Alterations in genes that encode enzymes that are involved in aflatoxin biosynthesis and its precursors could signify the observed difference in aflatoxin production by the mutants.
Partial repair of damaged genes could substantiate the reduction of aflatoxin output seen in some mutants in this study. Mutagenic disruption of genes that encode important enzymes that are involved in bioconversion of intermediate precursors to another forms, could in addition, led to differences in the level of aflatoxin production. For instance, enzymes such as polyketide synthase (Crawford et al., 2008) β-ketoacyl synthase, acyltransferase, acyl carrier protein and thioesterase (Chang et al., 1993) are some intermediate precursor-converting enzymes reported in aflatoxin synthesis. Incomplete expression of any gene involved in aflatoxin biosynthesis as result of altered genomes in mutants could be adduced to reduced level of aflatoxin produced. Chang et al. (1993) and Yu (2012) reported that incomplete blockage of aflatoxin biosynthesis via gene mutation in A. flavus resulted to the generation of norsolorinic-acid-accumulating mutants. Any impairment in DNA that affects gene products require in catalysis of any important intermediate precursors would have impact on aflatoxin production. This is in accordance to the report of Ichishima (2007) in oxidation of norsolorinic acid anthrone to averantin. Any of the aforementioned could give rise to the results obtained in this study.
Differences in the degree and capability of mutants to reconstitute damaged genes that are involved in aflatoxin synthesis could validate the significant differences in toxin production among the mutants. Damage and subsequent rebuilding of genes involved in genetic control of factor, AflR in aflatoxin synthesis could give raise to mutations responsible for differences in biosynthesis (Yu, 2012). The differential potentials of mutants to reconstruct impaired genes (such as AFLS (AFLJ ) gene, which encode a putative transcriptional co-activator (AFLS) and AFLR gene that encodes pathway-specific transcription (Yu, 2012) relevant in genetic control in aflatoxin biosynthetic pathway, could also prefigure the observed dissimilarities in aflatoxin biosynthesis by mutants. Any form of alteration in the genes, for instance laeA, which encodes a global regulator, LaeA and VeA gene, that encodes a gene regulator, VeA that are associated with genetic control of secondary metabolism, fungal development and mycotoxin formation (Yu, 2012), would exerted significant influence on aflatoxin production as noticed in the mutants developed in this study.
CONCLUSION
Ultraviolet radiation caused considerable repression (93.11%) of aflatoxin traits in A. flavus strain in contrast to other mutagens. Nevertheless, the equivalent concentration of aflatoxins produced still exceeded 50 parts per trillion (ppt) standard limits set by FAO/WHO/EU in human food products. Thus, the use of the mutant in fermentation could pose public health risk. Although, this study gave insights into possible control of aflatoxin production through random mutagenesis, further cycles of UV-treatment or molecular methods is suggested probably would get rid of aflatoxin yield or below the threshold limit for application in human food or food grade enzyme production. This will enhance potential industrial utilization of the aflatoxigenic fungi for fermentation purposes.
SIGNIFICANT STATEMENTS
Investigation of a quest into the ability of some mutagens to impact control of aflatoxin biosynthesis through random mutation of aflatoxigenic A. flavus SWtS01.
ACKNOWLEDGMENT
The effort of Mr. A.S. Kazeem (Nigeria Agricultural Quarantine Service, Moor Plantation, Nigeria) for supply of materials and access to equipment is highly thanked.
REFERENCES
- Hoffmeister, D. and N.P. Keller, 2007. Natural products of filamentous fungi: Enzymes, genes and their regulation. Nat. Prod. Rep., 24: 393-416.
CrossRefDirect Link - Frisvad, J.C., C. Rank, K.F. Nielsen and T.O. Larsen, 2009. Metabolomics of Aspergillus fumigatus. Med. Mycol., 47: 53-71.
CrossRefDirect Link - Elliaiah, P., T. Prabhakar, B. Ramakrishna, A.T. Taleb and K. Adinarayana, 2002. Strain improvement of Aspergillus niger for the production of lipase. Indian J. Microbiol., 42: 151-153.
Direct Link - Nadeem, M., J.I. Qazi and S. Baig, 2010. Enhanced production of alkaline protease by a mutant of Bacillus licheniformis N-2 for dehairing. Braz. Arch. Biol. Technol., 53: 1015-1025.
CrossRefDirect Link - Iftikhar, T., M. Niaz, S.Q. Abbas, M.A. Zia, I. Ashraf, K.J. Lee and Ikram-ul-Haq, 2010. Mutation induced enhanced biosynthesis of lipases by Rhizopus oligosporus var. microsporus. Pak. J. Bot., 42: 1235-1249.
Direct Link - Rajeshkumar, J. and M.H.M. Ilyas, 2011. Production of phophatase by mutated fungal strains. Int. Multidiscipl. Res. J., 1: 23-29.
Direct Link - Damisa, D., F.A. Kuta and N.U. Adabara, 2013. Mutagenic treatment of Aspergillus niger with hydroxylamine for improved cellulase synthesis from cellulosic wastes. J. Biotechnol. Sci., 1: 65-72.
Direct Link - Irfan, M., J. Javed and Q. Syed, 2011. UV mutagenesis of Aspergillus niger for enzyme production in submerged fermentation. Pak. J. Biochem. Mol. Biol., 44: 137-140.
Direct Link - Zaini, F., S. Yousefi, S. Dadgar and M. Safara, 2009. Aflatoxin production by Aspergillus flavus isolates from green-tiger shrimps (Penaeus semisulcatus). Iran. J. Microbiol., 1: 18-22.
Direct Link - Cyrus, E.T. and A.D. Juwon, 2015. Effects of radiation and chemical mutagenesis on expression of aflatoxigenic traits in aspergillus parasiticus SMS08-C. Res. J. Microbiol., 10: 205-213.
Direct Link - Hashida, M. and H. Bisgaard-Frantzen, 2000. Protein engineering of new industrial amylases. Trends Glycoscci. Glycotechnol., 12: 389-401.
CrossRefDirect Link - Chang, P.K., J.W. Bennett and P.J. Cotty, 2002. Association of aflatoxin biosynthesis and sclerotialdevelopment in Aspergillus parasiticus. Mycopathologia, 153: 41-48.
CrossRefDirect Link - Flaherty, J.E. and G.A. Payne, 1997. Overexpression of aflR leads to upregulation of pathway gene transcription and increased aflatoxin production in Aspergillus flavus. Applied Environ. Microbiol., 63: 3995-4000.
Direct Link - Crawford, J.M., P.M. Thomas, J.R. Scheerer, A.L. Vagstad, N.L. Kelleher and C.A. Townsend, 2008. Deconstruction of iterative multidomain polyketide synthase function. Science, 320: 243-246.
CrossRefDirect Link - Chang, P.K., J.W. Cary, D. Bhatnagar, T.E. Cleveland and J.W. Bennett et al., 1993. Cloning of the Aspergillus parasiticus apa-2 gene associated with the regulation of aflatoxin biosynthesis. Applied Environ. Microbiol., 59: 3273-3279.
Direct Link - Yu, J., 2012. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins, 4: 1024-1057.
CrossRefDirect Link