
Tersagh Ichor
Department of Biological Sciences, University of Agriculture, P.M.B 2373, Makurdi, Benue State, Nigeria
G.M. Gberikon
Department of Biological Sciences, University of Agriculture, P.M.B 2373, Makurdi, Benue State, Nigeria
Dooshima Nevkaa
Department of Biological Sciences, University of Agriculture, P.M.B 2373, Makurdi, Benue State, Nigeria







ABSTRACT
Biodegradation of phenanthrene by a consortium of aerobic heterotrophic bacteria and cyanobacteria isolated from hydrocarbon polluted brackish water of Bodo creek was monitored using GC-MS for 56 days. The initial quantity of phenanthrene in the treatment options monitored on day 0 was 2.8, 1.95, 3.17 and 2.76 mg L–1 for (A) Aerobic heterotrophic bacteria, (B) Cyanobacteria, (AB) aerobic heterotrophic bacteria+cyanobacteria and (C) Control, respectively on day 56. Mineralization of phenanthrene progressed to 0.1, 0.0, 0.01 and 0.0 mg L–1 for the treatments of A, B, A+B and C, respectively. The quantity of phenanthrene in all the treatments fluctuated throughout the period monitored hence its biodegradation did not vary significantly with time, t(4) = 1.76 (p>0.05).
PDF Abstract XML References Citation
Received: October 14, 2015;
Accepted: November 08, 2015;
Published: March 15, 2016
How to cite this article
Tersagh Ichor, G.M. Gberikon and Dooshima Nevkaa, 2016. Biodegradation of Phenanthrene by a Consortium of Aerobic Heterotrophic Bacteria and Cyanobacteria in Petroleum Hydrocarbon Polluted Brackish Water of Bodo Creek. Microbiology Journal, 6: 1-8.
DOI: 10.3923/mj.2016.1.8
URL: https://scialert.net/abstract/?doi=mj.2016.1.8
DOI: 10.3923/mj.2016.1.8
URL: https://scialert.net/abstract/?doi=mj.2016.1.8
INTRODUCTION
Polycyclic Aromatic Hydrocarbons (PAHs) are compounds containing carbon and hydrogen with the carbon arranged in a series of adjoining six member benzene rings and is present in water, crude oil and tars. They are known to be recalcitrant and are not easily degradable. They are considered hazardous because of their potential trophic biomagnifications and acute toxicity (Darville and Wilhm, 1984). The fate of PAHs in the environment in nature is of great environmental concern due to their toxic, mutagenic and carcinogenic properties and is associated with both abiotic and biotic processes, including volatilization, photooxidation, chemical oxidation, bioaccumulation and microbial transformation. Microbial activity has been deemed the most influential and significant cause of PAH removal (Cerniglia, 1992; Nwuche and Ugoji, 2008; Haghighat et al., 2008; Agbozu and Opuene, 2009; Atlas and Bragg, 2009).
Phenanthrene is a tricyclic aromatic hydrocarbon that is present in high concentrations in Polycyclic Aromatic Hydrocarbon (PAH) contaminated sediments, surface soils and waste sites. It is low molecular weight PAH that is hydrophobic and widely distributed in the environment, occurring as natural constituents of fossil fuels and their anthropogenic pyrolysis products (Kanaly and Harayama, 2000; Wilson and Jones, 1993) and readily adsorbed onto particulate matter and thus, coastal and marine sediments become the ultimate sinks for such compounds (Hughes et al., 1997; Yu et al., 2005; Osuji and Ezebuiro, 2006). It does not pose a risk to human health since it is not known to exhibit any genotoxic or carcinogenic effects though it is reported to be toxic to fish and algae (Moody et al., 2001). It is considered a prototypic PAHs and serve as one of the signature compounds to detect PAH contamination since its structure is found in carcinogenic PAHs such as; benzo (a) pyrene and benz (a) anthracene. It has also been used as model PAHs to determine factors that affect the bioavailability biodegradation potential and rate of microbial degradation of PAHs in the environment (Kanaly and Harayama, 2000; Sutherland et al., 1995; Bouchez et al., 1995). However, other authorities report that phenanthrene is known to be a human skin photosensitizer and mild allergen (Fawell and Hunt, 1988) and has also been found to be an inducer of sister chromatid exchanges and a potent inhibitor of gap junction intercellular communications (Weis et al., 1998).
The role of indigenous microbial communities in the removal of hydrocarbon from the environment has been reported by other researchers in previous studies. Investigations shows that a small fraction of all natural microbial populations irrespective of location and environmental factors can grow on both aromatic and aliphatic hydrocarbons (Ruberto et al., 2003; Solano-Serena et al., 1999). A variety of bacterial species that possess the capability to utilize phenanthrene as the sole source of carbon and energy have been isolated in previous studies (Cerniglia, 1992; Sutherland et al., 1995). Menn et al. (1993) reported degradation of phenanthrene by Pseudomonas species, Streptomyces flavovirens (Sutherland et al., 1993); Mycobacterium, Nocardia and Rhodococcus species (Churchill et al., 1999; Dean-Ross and Cerniglia, 1996; Rehmann et al., 1998; Schneider et al., 1996; Saito et al., 2000); Bacillus species (Doddamani and Ninnekar, 2000). Janbandhu and Fulekar (2011) identified Sphingobacterium sp., Bacillus cereus and Achromobacter insolitus MHF ENV IV with effective phenanthrene-degrading ability. The biodegradation data indicated that about 100, 56.9 and 25.8% degradation at a concentration of 100, 250 and 500 mg L–1 was achieved within 14 days by the isolates, respectively. Shiaris (1989) reported that rates of phenanthrene mineralization along a natural salinity gradient in an Urban Estuary, Boston Harbor, Massachusetts related significantly to increasing salinity along the transect as determined by linear regression analysis and were unaffected by higher salinity waters. The result suggests that phenanthrene degradation in brackish waters is mainly a function of obligate marine microorganisms. Tang and Krieger-Brockett (2007) measured the mineralization rates of 14C-phenanthrene in contaminated, undisturbed marine sediments using whole core injection method to assess the natural attenuation of microbial activity as a function of sediment depth. The experiment showed significant biodegradation activities (0.0012-0.0036 day–1) in the sediment depth from 0-10 cm.
Effective microbial consortium has attracted the attention of researchers worldwide due to synergistic interactions among members of the consortium (Tao et al., 2007). A microbial consortium W4 isolated from Henan oil field was reported to have effectively degraded phenanthrene (Jacques et al., 2008). Wang et al. (2008) investigated biodegradation of solid phenanthrene at 200 mg L–1 in liquid medium by a microbial consortium isolated from Henan oil field after 6 days of incubation and reported that the degradation was higher than 95% at 37°C and 120 rpm. Arulazhagan et al. (2010) monitored the mineralization of phenanthrene from marine environment by bacterial consortium and reported 99% phenanthrene degradation in 4 days. There is a dearth of information from available literatures on degradation of phenanthrene by cyanobacteria and associated aerobic heterotrophic bacteria which is the focus of this study.
The study sought to monitor biodegradation of phenanthrene by aerobic heterotrophic bacteria, cyanobacteria and the consortium of aerobic heterotrophic bacteria and cyanobacteria and also to isolate and identity the microorganisms involved using conventional culturing and molecular approaches.
MATERIALS AND METHODS
The study area is oil contaminated water body, Bodo creek located in Ogoni land in Gokana, LGA of Rivers State, in Niger Delta region.
Media and incubation conditions: BG-11 medium consisting in (g L–1) of NaNO3 (1.5), K2HPO4 (0.004), MgSO4. 7H2O (0.0075), CaCl2.2H2O (0.036), Na2CO3 (0.02), citric acid (0.006), ferric ammonium citrate (0.006), disodium magnesium EDTA (0.001) and trace metal solution 1.0 mL L–1 comprising of H3BO3, MnCl2.4H2O, Co(NO3)2.6H2O, ZnSO4.7H2O, Na2Mo4.2H2O, CuSO4.5H2O was prepared for the growth of cyanobacteria and adjusted to a pH of 7.4. The medium was prepared according to the methods of Salleh et al. (2003).
Enumeration of cyanobacteria: Ten brackish water and sediment samples were collected from crude oil contaminated Bodo creek in Rivers State. The water sample was shaken to suspend sediment and then 1 mL of the raw water sample was removed and diluted into 99 mL of sterile distilled water which was serially diluted and 1 mL removed from the third dilution and plated on BG-11 medium solidified with agar agar and treated with ciprofloxacin and nystatin in duplicate plates. Precisely, 1 g of the soil sediment sample was dissolved in 9 mL of distilled water and filtered using, Whatman No. 1 filter paper.
The filterate was serially diluted and same procedure followed as in water. An aliquot of cyanobacteria culture was prepared using different medium- water volume ratio of 90:10, 80:20, 70:30, 60:40 and 50:50 mL and vice-versa and incubated in a cotton wool corked Erlenmeyer flask for 14 days and exposed to natural sunlight for 12 h and darkness for 12 h under ambient temperature. This was monitored and shaken twice daily to avoid sedimentation of the nutrients from the BG-11 medium.
Enumeration of total hydrocarbon utlizing bacteria: The vapour phase transfer method was adopted for enumeration of hydrocarbon utilizing bacteria, following serial dilutions of water and sediment samples in the respective treatments, aliquots of 0.1 mL of the dilutions was plated out in duplicates on a sterile mineral salt agar as described by Mills et al. (1978).
The salt agar contained the following in g L–1; NaCl 10.0 MgSO4. 7H2O, 0.42, KCl 0.29, KH2PO4 0.83, K2HPO4 1.25, NH4NO3 0.42, agar 15.0 and distilled water.
Sterile Whatman No. 1 filter paper saturated with crude oil was placed on the inside cover of each of the Petri dishes and incubated with the agar side up. The filter supplied the microorganisms with hydrocarbon by vapour phase transfer to the inocula. The plates were placed in the incubator (Gallenkamp Duostat incubator size 2 England) at 37°C for 7days after which colonies were counted to determine titre (viable counts per unit) of the water and sediment samples (Ichor et al., 2014).
Preparation of inoculum and biodegradation experiment: Aerobic heterotrophic bacteria aliquot was prepared by transferring a loopful of 24 h culture of each isolate into 400 mL of sterile nutrient broth in 500 mL Erlenmeyer flask and incubated for 24 h.
A bloom culture of cyanobacterial aliquot using BG II medium formulations was also prepared by transterring 1 mL of each viable culture in to 400 mL of the medium and incubated for 48 h under natural sunlight for 12 h and darkness for 12 h.
Treatment options of brackish water samples labeled AHB and CB were prepared by asceptically transferring 200 mL each of aerobic heterotrophic bacteria and cyanobacteria aliquot into 500 mL of sterile distilled water in two separate 1000 mL flask, respectively. The AHB+CB was prepared by transferring 200 mL each of aerobic heterotrophic bacteria and cyanobacteria aliquot into another 500 mL of sterile distilled water in 1000 mL flask. Each option was standardized using 0.5 M Macfarland solution (Ichor et al., 2014). The inoculum size used for each treatment option was 100 mL of standardized aliquot. The water containers for the experimental set up were each filled with 11 L of water and labeled AHB, CB, AHB+CB and C for the control and contaminated with 32300 ppm of sterile Bony light crude oil sample obtained from shell petroleum development company. Water samples AHB, CB, AHB+CB were inoculated with aerobic heterotrophic bacteria, cyanobacteria and aerobic heterotrophic bacteria+cyanobacteria, respectively while the control was left uninoculated. Sample CB was treated with 0.25 mg mL–1 of ciprofloxacin and nystatin, AHB was treated with CuSO4 and nystatin to prevent cyanobacteria and fungal activities in the set up, AHB+CB was treated with only 0.25 mg mL–1 of nystatin while the control was left untreated. The containers for the treatments were washed with detergents and rinsed severally with distilled water prior to the experimental set up (Ichor et al., 2014).
Molecular analysis of bacterial and cyanobacterial isolates DNA extraction and polymerase chain reaction: Gene sequencing and analysis for bacterial isolates used was done as reported by Ichor et al. (2014). Universal primers CYA 106F (CGC ACG GGT GAG TAA CGC GTG A and CYA 359F(GGG GAA TYT TCC GCA ATG GG) with a 40 nucleotide GC clamp (5CGC CCG CCG CGC CCC GCG CCG GTC CCG CCG CCC CCG CCC G 3) on the 5 end forward primer and CYA 781R (equimolar mixture of CYA781Ra (GAC TAC TGG GGT ATC TAA TCC CAT T) and CYA 781Rb (GAC TAC AGG GGT ATC TAA TCC CTT T) reverse primers for amplification of a segment of cyanobacterial 16SrRNA gene (70) were synthesized. A semi nested PCR reaction was carried out with the first reaction using primers CYA 106F and CYA 781R and followed by a reaction with primers CYA 359F and CYA781R. The PCR was carried out in a 25 μL final volume of reaction mixture containing 100 ng of DNA 2.5 μL of 10x PCR buffer with 200 μg bovine serum albumin (nuclease free) and 0.2 μ Taq DNA polymerase ( Banngalore Genei, India) in a 1 cycler (BioRad, USA). The thermal cycling profile was thus; initial denaturation for 3 min at 94°C followed by 35 amplification cycles each consisting of 1.5 min denaturation at 94°C, 1 min annealing at 59°C and a 2 min elongation at 72°C with a final 5 min elongation at 72°C.
Sequences obtained were compared with known sequences in the GenBank using the Basic Local Alignment Search Tool (BLAST) of the National Centre for Biotechnology Information (NCBI). Species were identified based on the percentage similarity with known sequences in the data base (Ichor et al., 2014).
RESULTS
The hydrocarbon degrading aerobic heterotrophic bacteria and their accession numbers isolated from water and sediment from Bodo creek implicated in phenanthrene degradation are in were as reported by Ichor et al. (2014); cyanobacteria isolates used in mineralization of phenanthrene. Figure 1 depicts phenanthrene degradation by (A) Aerobic heterotrophic bacteria, (B) Cyanobacteria, (A+B) Consortium of aerobic heterotrophic bacteria and (C) Cyanobacteria and control as monitored for 56 days using GC-MS. The initial quantity of phenanthrene in A- 2.8, B-1.95, A+B-3.17 and C-2.76 which reduced to 0.68, 0.05, 1.28 and 1.27 after seven days of inoculation as monitored. The quantity of phenanthrene in all the treatment fluctuated throughout the period monitored but completely mineralized to 0.0 for treatment B and C, where, as treatments A and A+B had 0.1 and 0.01 respectively.
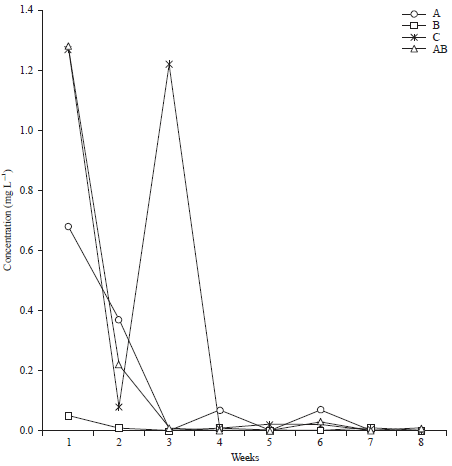 | |
| Fig. 1: | Biodegradation of phenanthrene |
Biodegradation of phenanthrene did not significantly vary with time, t(4) = 1.76 (p<0.05).
DISCUSSION
A study was carried out a study to monitor biodegradation of phenanthrene by isolates from petroleum hydrocarbon contaminated brackish water of Bodo creek. The bacteria isolated from water and identified with their accession numbers include; Alcaligenes faecalis (KF056900.1), Bacillus pumilus (KF717600.1), Bacillus pumilus (JX680132.1), Bacillus pumilus (HQ334985.1), Myroides odoratimimus (GU186112.2) Bacillus cohnii (KC813164.1), Citrobacter murliniae (KF254752.1), Bacillus pumilus (HM744710.1), Bordetella sp. (KF601910.1), Lysinibacillus fusiformis (KC329831.1), Bacillus pumilus KF387715.1), Myroides pelagicus (NR041042), Bacillus pumilus (JQ833612.1) and Enterobacter cloacae (JQ832514.1), where, as Bacillus safensis (KF 641818.1), Bacillus pumilus (FJ906741.1), Bacillus pumilus (KC492105.1), Bacillus pumilus (HQ334984.1), Bacillus licheniformis (JQ835466.1), Cedecea davisae (KC201361.1), Enterobacter hormaechei (JQ829491.1), Bacillus pumilus (KF717600.1), Bacillus pumilus (JQ833773.1), Bacillus subtilis (JQ832514.1), Enterobacter hormaechei (JQ8299302.1), Bacillus aeophilus (KC414716.1), Citrobacter freundii (RF535142.1), Pseudomonas aeruginosa (JQ659950.1), Bacillus megaterum (JQ881645.1) and Escherichia coli (HE583396.1) were isolated from sediments. The isolates belonged to the families of Enterobacteriaceae, Bacilliceae, Alcaligenaceae, Pseudomonadaceae, Flavobactericeae and Planococcaceae (Ichor et al., 2014). The cyanobacteria isolated from surface water of Bodo creek with their accession numbers include Microleus sp. (KF157400.1), Oscillatoriales cyanobacterium (KC163194.1), Phormidium sp. (EU282429.1), Synechococcus sp. (GQ130146.1), Microcystis elabens (U40335.1), Anabaena cicadae (KF15740.1) Pseudanabaena minima (HQ132935.1), Leptolyngbya sp. (JX401929.1), Microcystis holsatica (U40336.1), Phormidium faveolarum and Filamentous cyanobacterium (KF793929.1) (Ichor et al., 2014). The bacterial, cyanobacterial isolates and the consortium of aerobic heterotrophic bacteria and cyanobacteria studied degraded phenanthrene effectively on the 56th day as monitored using GC-MS though fluctuations were observed. The fluctuations could be due to novel synthesis where complex molecules break down to form simpler ones, in this could some complex components of hydrocarbons could have been metabolized in to phenanthrene or its metabolites during the biodegradation process thus increasing its quantity in the medium. Phenanthrene mineralization rates was previously reported at varying degrees of salinity and the mineralization related significantly to increasing salinity along the transect as determined by linear regression analysis. Rates ranged from 1 ng/h/g dry sediment at the freshwater site to>16 ng/h/g dry sediment at the 30% salinity site (Shiaris, 1989). The results suggested that phenanthrene degraders in low salinity estuarine sediments subject to salt water intrusion are tolerant to a wide range of salinities but phenanthrene degradation in brackish waters is mainly a function of obligate marine microorganisms. Our study isolated cyanobacteria and aerobic heterotrophic bacteria which are resident flora of Bodo creek and tested for biodegradation of phenanthrene which result showed efficiency of phenanthrene removal.
Evidence abounds from previous researchers on biodegradation of phenanthrene by indigenous microorganisms in saline water. Narro et al. (1992) reported on biodegradation of phenanthrene by the marine cyanobacterium Agmenellum quadruplicatum PR-6. The marine cyanobacterium A. quadruplicatum PR-6 metabolized phenanthrene to form phenanthrene trans-9,10-dihydrodiol and 1-methoxyphenanthrene as the major metabolites. However, the beneficial application of cyanobacteria in remediation of contaminated waters, either natural aquatic environments or industrial effluents, is still not optimally manipulated. For example, cyanobacterial species such as; Oscillatoria salina, Plectonema terebrans, Aphanocapsa sp. and Synechococcus sp., developed as mats in aquatic environments, have been successfully used in bioremediation of oil spills in different parts of the world (Raghukumar et al., 2001; Radwan and Al-Hasan, 2001; Cohen, 2002). Ji et al. (2013) investigated biodegradation of phenanthrene by Aulosira fertilissima Ghose using GC MS and reported 80% efficiency for its removal and biodegradation after 7 days of treatment. These reports corroborate our findings on the consortium of cyanobacteria studied though no amendment was applied to enhance its removal.
There have been reports from previous studies on the removal of phenanthrene by bacteria. Zhao et al. (2009) reported on the degradation of phenanthrene in the presence of 5-15% NaCl by a halophilic bacterial consortium developed from soil samples collected from the Shengli oil field in China and phenanthrene was completely degraded by the enrichment in 8 days. Molecular analysis of the enrichment culture indicated the presence of alpha and gamma-proteobacteria including members of the genus Halomonas, Chromohalobacter, Alcanivorax, Marinobacter, Idiomarina and Thalassospira. Dastgheib et al. (2012) obtained a mixed culture (Qphe-SubIV) consisting of Halomonas sp. and Marinobacter sp. from hydrocarbon-contaminated saline soil collected from five different regions in Iran. The organisms degraded several PAHs including naphthalene, phenanthrene, anthracene, fluoranthene, fluorine, pyrene, benz[a]anthracene and benzo[a]pyrene as the sole carbon sources in the presence of 1-15% NaCl. Al-Mailem et al. (2013) investigated on the ability of Marinobacter sedimentalis and Marinobacter falvimaris isolated from a hypersaline in sabkhas and reported on their capability to degrade biphenyl, phenanthrene, anthracene and naphthalene as the sole sources of carbon and energy at 6% NaCl. Gao et al. (2013) isolated Marinobacter nanhaiticus strain D15-8W from a phenanthrene-degrading enrichment obtained from a sediment from the South China Sea. The strain D15-8W degraded naphthalene, phenanthrene or anthracene as the sole source of carbon and energy in the presence of 0.5-15% with optimum degradation in the presence of 1-5% NaCl. Erdogmus et al. (2013) showed the degradation of naphthalene, phenanthrene and pyrene as the sole carbon sources in 20% NaCl by several archaeal strains including; Halobacterium piscisalsi, Halorubrum ezzemoulense, Halobacterium salinarium, Haloarcula hispanica, Haloferax sp. Halorubrum sp. and Haloarcula sp., isolated from brine samples of Camalt Saltern in Turkey.
Present study showed degradation of phenanthrene by aerobic heterotrophic bacteria isolated from brackish water and sediment samples with salinity of 19.67 and 17.87 ppt respectively in 56 days monitored. The result of this study showed that the genus Bacillus dominated the water and sediment samples among other genera like Alcaligenes, Enterobacter, Pseudomonas and Escherichia which are known to be very frequent in sea water and were also isolated from petroleum hydrocarbon polluted aquatic environments. The isolated aerobic heterotrophic bacteria belong to the families of Enterobacteriaceae, Bacilliceae, Alcaligenaceae, Pseudomonadaceae, Flavobactericeae and Planococcaceae. (Ichor et al., 2014).
CONCLUSION
The consortium of the isolated aerobic heterotrophic bacteria and cyanobacteria degraded phenanthrene throughout the 56 days period monitored though fluctuations were observed just as it was obtainable in other treatment options. It is proven by previous study reports that degradation of PAHs could not be oxidized effectively by one single species. It was apparently more effectively done by a consortium since mixed cultures were observed to outperform single species even when isolated from the consortium. Degradation of phenanthrene also occurred in the control implying that Bodo creek characterized by brackish water harbours microorganisms with inherent capabilities for petroleum hydrocarbon. Further study was suggested on the metabolic pathway/products of phenanthrene biodegradation by resident microorganisms in Bodo creek.
REFERENCES
- Agbozu, I.E. and K. Opuene, 2009. Occurrence and diagenetic evolution of perylene in the sediments of Oginigba creek, Southern Nigeria. Int. J. Environ. Res., 3: 117-124.
Direct Link - Al-Mailem, D.M., M. Eliyas and S.S. Radwan, 2013. Oil-bioremediation potential of two hydrocarbonoclastic, diazotrophic Marinobacter strains from hypersaline areas along the Arabian Gulf coasts. Extremophiles, 17: 463-470.
CrossRefDirect Link - Janbandhu, A. and M.H. Fulekar, 2011. Biodegradation of Phenanthrene using adapted microbial consortium isolated from petrochemical contaminated environment. J. Hazard. Mater. 187: 333-340.
CrossRefPubMedDirect Link - Arulazhagan, P., N. Vasudevan and I.T. Yeom, 2010. Biodegradation of polycyclic aromatic hydrocarbon by a halotolerant bacterial consortium isolated from marine environment. Int. J. Environ. Sci. Technol., 7: 639-652.
CrossRefDirect Link - Atlas, R. and J. Bragg, 2009. Bioremediation of marine oil spills: When and when not-the Exxon Valdez experience. Microbial Biotechnol., 2: 213-221.
CrossRefDirect Link - Bouchez, M., D. Blanchet and J.P. Vandecasteele, 1995. Substrate availability in phenanthrene biodegradation: Transfer mechanism and influence on metabolism. Applied Microbiol. Biotechnol., 43: 952-960.
CrossRefDirect Link - Cerniglia, C.E., 1992. Biodegradation of polycyclic aromatic hydrocarbons. Biodegradation, 3: 351-368.
CrossRefDirect Link - Churchill, S.A., J.P. Harper and P.F. Churchill, 1999. Isolation and characterization of a Mycobacterium species capable of degrading three- and four-ring aromatic and aliphatic hydrocarbons. Applied Environ. Microbiol., 63: 549-552.
Direct Link - Cohen, Y., 2002. Bioremediation of oil by marine microbial mats. Int. Microbiol., 5: 189-193.
CrossRefDirect Link - Dastgheib, S.M.M., M.A. Amoozegar, K. Khajeh, M. Shavandi and A. Ventosa, 2012. Biodegradation of polycyclic aromatic hydrocarbons by a halophilic microbial consortium. Applied Microbiol. Biotechnol., 95: 789-798.
CrossRefDirect Link - Dean-Ross, D. and C.E. Cerniglia, 1996. Degradation of pyrene by Mycobacterium flavescens. Applied Microbiol. Biotechnol., 46: 307-312.
CrossRefDirect Link - Doddamani, H.P. and H.Z. Ninnekar, 2000. Biodegradation of phenanthrene by a Bacillus species. Curr. Microbiol., 41: 11-14.
CrossRefDirect Link - Erdogmus, S.F., B. Mutlu, S.E. Korcan, K. Guven and M. Konuk, 2013. Aromatic hydrocarbon degradation by halophilic archaea isolated from Camalti Saltern, Turkey. Water Air Soil Pollut., Vol. 224.
CrossRefDirect Link - Gao, W., Z. Cui, Q. Li, G. Xu, X. Jia and L. Zheng, 2013. Marinobacter nanhaiticus sp. nov., polycyclic aromatic hydrocarbon-degrading bacterium isolated from the sediment of the South China Sea. Antonie Van Leeuwenhoek, 103: 485-491.
CrossRefDirect Link - Haghighat, S., A.A. Sepahy, M.M. Assadi and H. Pasdar, 2008. Ability of indigenous Bacillus licheniformis and Bacillus subtilis in microbial enhanced oil recovery. Int. J. Environ. Sci. Technol., 5: 385-390.
CrossRefDirect Link - Hughes, J.B., D.M. Beckles, S.D. Chandra and C.H. Ward, 1997. Utilization of bioremediation processes for the treatment of PAH-contaminated sediments. J. Ind. Microbiol. Biotechnol., 18: 152-160.
CrossRefDirect Link - Ichor, T., P.O. Okerentugba and G.C. Okpokwasili, 2014. Biodegradation of total petroleum hydrocarbon by aerobic heterotrophic bacteria isolated from crude oil contaminated brackish waters of bodo creek. J. Bioremed. Biodeg., Vol. 5.
Direct Link - Moody, J.D., J.P. Freeman, D.R. Doerge and C.E. Cerniglia, 2001. Degradation of phenanthrene and anthracene by cell suspensions of Mycobacterium sp. Strain PYR-1. Applied Environ. Microbiol., 67: 1476-1483.
CrossRefPubMedDirect Link - Kanaly, R.A and S. Harayama, 2000. Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria. J. Bacteriol., 182: 2059-2067.
CrossRefDirect Link - Ji, N.K., M. Barot and S.R. Khan, 2013. Some intermediate bio-transformants during biodegradation of high molecular weight phenanthrene and fluoranthene by cyanobacterial species-Aulosira fertilissima ghose. Int. J. Applied Sci. Biotechnol., 1: 97-105.
CrossRefDirect Link - Menn, F.M., B.M. Applegate and G.S. Sayler, 1993. NAH plasmid-mediated catabolism of anthracene and phenanthrene to naphthoic acids. Applied Environ. Microbiol., 59: 1938-1942.
Direct Link - Narro, M.L., C.E. Cerniglia, C. van Baalen and D.T. Gibson, 1992. Metabolism of phenanthrene by the marine cyanobacterium Agmenellum quadruplicatum PR-6. Applied Environ. Microbiol., 58: 1351-1359.
Direct Link - Nwuche, C.O. and E.O. Ugoji, 2008. Effects of heavy metal pollution on the soil microbial activity. Int. J. Environ. Sci. Tech , 5: 409-414.
Direct Link - Osuji, L.C. and P.E. Ezebuiro, 2006. Hydrocarbon contamination of a typical mangrove floor in Niger Delta, Nigeria. Int. J. Environ. Sci. Technol., 3: 313-320.
Direct Link - Radwan, S.S. and R.H. Al-Hasan, 2001. Potential application of coastal biofilm-coated gravel particles for treating oily waste. Aquat. Microb. Ecol., 23: 113-117.
Direct Link - Raghukumar, C., V. Vipparty, J. David and D. Chandramohan, 2001. Degradation of crude oil by marine cyanobacteria. Applied Microbiol. Biotechnol., 57: 433-436.
CrossRefDirect Link - Rehmann, K., H.P. Noll, C.E.W. Steinberg and A.A. Kettrup, 1998. Pyrene degradation by Mycobacterium sp. Strain KR2. Chemosphere, 36: 2977-2992.
CrossRefDirect Link - Jacques, R.J.S., B.C. Okeke, F.M. Bento, A.S. Teixeira, M.C.R. Peralba and F.A.O. Camargo, 2008. Microbial consortium bioaugmentation of a polycyclic aromatic hydrocarbons contaminated soil. Bioresour. Technol., 99: 2637-2643.
CrossRefDirect Link - Ruberto, L., S.C. Vazquez and W.P.M. Cormack, 2003. Effectiveness of the natural bacterial flora, biostimulation and bioaugmentation on the bioremediation of a hydrocarbon contaminated Antarctic soil. Int. Biodeterior. Biodegrad., 52: 115-125.
CrossRefDirect Link - Saito, A., T. Iwabuchi and S. Harayama, 2000. A novel phenanthrene dioxygenase from Nocardioides sp. strain KP7: Expression in Escherichia coli. J. Bacteriol., 182: 2134-2141.
CrossRefDirect Link - Salleh, A.B., F.M. Ghazali, R.N.Z.A. Rahman and M. Basri, 2003. Bioremediation of petroleum hydrocarbon pollution. Indian J. Biotechnol., 2: 411-425.
Direct Link - Schneider, J., R. Grosser, K. Jayasimhulu, W. Xue and D. Warshawsky, 1996. Degradation of pyrene, benz[a]anthracene and benzo[a]pyrene by Mycobacterium sp. strain RJGII-135, isolated from a former coal gasification site. Applied Environ. Microbiol., 62: 13-19.
Direct Link - Shiaris, M.P., 1989. Phenanthrene mineralization along a natural salinity gradient in an urban estuary, Boston harbor, massachusetts. Microbial Ecol., 18: 135-146.
CrossRefDirect Link - Solano-Serena, F., R. Marchal, M. Ropars, J.M. Lebeault and J.P. Vandecasteele, 1999. Biodegradation of gasoline: Kinetics, mass balance and fate of individual hydrocarbons. J. Applied Microbiol., 86: 1008-1016.
CrossRefDirect Link - Sutherland, J.B., P.P. Fu, S.K. Yang, L.S. von Tungeln, R.P. Casillas, S.A. Crow and C.E. Cerniglia, 1993. Enantiomeric composition of the trans-dihydrodiols produced from phenanthrene by fungi. Applied Environ. Microbiol., 59: 2145-2149.
Direct Link - Tang, Y.J. and B. Krieger-Brockett, 2007. Mathematical analysis of the whole core injection method accuracy for measuring phenanthrene biodegradation rates in undisturbed marine sediments. Chemosphere, 68: 804-813.
CrossRefDirect Link - Tao, X.Q., G.N. Lu, Z. Dang, C. Yang and X.Y. Yi, 2007. A phenanthrene-degrading strain Sphingomonas sp. GY2B isolated from contaminated soils. Process Biochem., 42: 401-408.
CrossRefDirect Link - Wang, J., H. Xu, M. An and G. Yan, 2008. Kinetics and characteristics of phenanthrene degradation by a microbial consortium. Petroleum Sci., 5: 73-78.
CrossRefDirect Link - Weis, L.M., A.M. Rummel, S.J. Masten, J.E. Trosko and B.L. Upham, 1998. Bay or baylike regions of polycyclic aromatic hydrocarbons were potent inhibitors of Gap junctional intercellular communication. Environ. Health Perspect., 106: 17-22.
Direct Link - Wilson, S.C. and K.C. Jones, 1993. Bioremediation of soil contaminated with polynuclear aromatic hydrocarbons (PAHs): A review. Environ. Pollut., 81: 229-249.
Direct Link - Yu, S.H., L. Ke, Y.S. Wong and N.F.Y. Tam, 2005. Degradation of polycyclic aromatic hydrocarbons by a bacterial consortium enriched from mangrove sediments. Environ. Int., 31: 149-154.
CrossRefDirect Link - Zhao, B., H. Wang, X. Mao and R. Li, 2009. Biodegradation of phenanthrene by a halophilic bacterial consortium under aerobic conditions. Curr. Microbiol., Vol. 58.
CrossRefDirect Link - Darville, R.G. and J.L. Wilhm, 1984. The effect of naphthalene on oxygen consumption and hemoglobin concentration in Chironomus attenuatus and on oxygen consumption and life cycle of Tanytarsus dissimilis. Environ. Toxicol. Chem., 3: 135-141.
CrossRefDirect Link - Mills, A.L., C. Breuil and R.R. Colwell, 1978. Enumeration of petroleum-degrading marine and estuarine microorganisms by the most probable number method. Can. J. Microbial., 24: 552-557.
PubMedDirect Link
Johnson onyeke Reply
Wow,this is interesting to know.