
E.O. Ibukun
Department of Biochemistry, Federal University of Technology Akure, Ondo State, Nigeria
G.O. Oladipo
Department of Biochemistry, Federal University of Technology Akure, Ondo State, Nigeria







ABSTRACT
Objective: This study was aimed at evaluating the renaloprotective effect of alcoholic red wine against CCl4-induced toxicity. Methodology: In vitro antioxidant activity of the alcoholic red wine was typified by total antioxidant activity, ABTS, DPPH and hydroxyl radicals, reducing power and metal chelating activities. In vivo evaluation of the ameliorative effect of alcoholic red wine against CCl4-renalotoxicity was experimented on kidney function by the profiling of biochemical indices, such as alkaline phosphatase (ALP) activity, creatinine clearance, Total Protein (TP) and the antioxidant status evaluated by activities of superoxide dismustase (SOD), glutathione peroxidase (GSH-Px), catalase (CAT) and glutathione-S-transferase (GST). Results: The results revealed that extracts of red wine had total antioxidant activity value of 96.57±6.441 mg g–1 AAE. Conclusion: The study revealed that the alcoholic red wine has ameliorative effect, which did not nullify the effect of the toxicological stress in all of the results.
PDF Abstract XML References Citation
Received: October 27, 2015;
Accepted: March 31, 2016;
Published: September 15, 2016
How to cite this article
E.O. Ibukun and G.O. Oladipo, 2016. Ameliorative Potential of Alcoholic Red Wine Against CCl4-induced Renalotoxicity in Albino Rats. Kidney Research Journal, 6: 1-8.
DOI: 10.3923/krj.2016.1.8
URL: https://scialert.net/abstract/?doi=krj.2016.1.8
DOI: 10.3923/krj.2016.1.8
URL: https://scialert.net/abstract/?doi=krj.2016.1.8
INTRODUCTION
Red wine basically come from the fruit (skins and seeds) and vine stems. The more represented phenol groups in wine are flavonoids and non-flavonoids, which are again present in a larger amount in red than in white wines and include free and conjugated myricetin, quercetin, kaempferol and isorhamnetin; (+)-catechin, (-)-epicatechin, gallic acid, p-coumaric acid, caffeic acid, caftaric acid, trans-resveratrol, cis-resveratrol and trans-resveratrol glucoside1. The delphinidin-like flavonolsmyricetin, laricitrin and syringetin were missing in all white varieties, indicating that the enzyme flavonoid 3, 5-hydroxylase is not expressed in white grape varieties2. Moderate intake of alcohol from any type of alcoholic beverage has been strongly associated with several beneficial effects on the cardiovascular system3. Since then several epidemiological studies have further assessed the beneficial effects of regular wine consumption on the cardiovascular risk4,5.
Free radical induced lipid peroxidation is believed to be one of the major causes of cell membrane damage leading to a number of pathological situations6. The CCl4 was formerly used for metal degreasing and as drycleaning, fabric-spotting and fire extinguisher fluids, grain fumigant and reaction medium. Because of its harmful effects, these uses are now banned and it is only used in some industrial applications7. The primary routes of potential human exposure to CCl4 are inhalation, ingestion and dermal contact. High exposure to CCl4 can cause liver, kidney and central nervous system damage and liver is especially sensitive to CCl4 because of its role as the body’s principal site of metabolism8.
The leading mechanism underlying improved endothelial function with regular moderate wine intake is an increase of nitric oxide synthesis9, most likely due to a remarkable increase of nitric oxide synthase promotor activity10. This study aimed at evaluate the renaloprotective effect of alcoholic red wine (11%) against CCl4 intoxication as well as evaluate the in vivo effect of alcoholic red wine consumption on the kidney.
MATERIALS AND METHODS
Chemicals, reagents and samples: Reagents and chemicals used in this experiment were obtained from different sources such as British Drug House (BDH) and Sigma limited and were all of good analytical grades. All the solutions, buffers and reagents were prepared using glass distilled water. The alcoholic red wine (11%) used was bought from a registered supermarket in Akure, Ondo State, Nigeria. The wine was concentrated using rotary evaporator and rediluted with distilled water to concentration, which served as the stock of 500 mg mL–1 used for in vitro antioxidant evaluation.
In vitro antioxidant assays
Assay for total antioxidant activity: Using standard laboratory protocol to the reagent solution; sulphuric acid (0.6 M), sodium phosphate (28 mM) and ammonium molybdate (4 mM); 0.3 mL of extract was added and incubated at 95°C in a water bath for 90 min. After cooling to room temperature absorbance was recorded at 765 nm against reagent blank. The absorbance of the sample was extrapolated on the ascorbic acid standard curve to obtain concentration of the sample in mg mL–1 then the total antioxidant activity (mg g–1 ascorbic acid equivalence) was calculated.
Assay of reducing power activity: The reducing power of the extracts was determined according to the method of Oyaizu11. Extract (0.5 mL) was mixed with 1.25 mL each of phosphate buffer and potassium ferricyanide (C6N6FeK3). The mixture was incubated at 50°C for 20 min. Trichloroacetic acid (1.25 mL) was then added and the mixture centrifuged at 3000 rpm for 10 min. Thereafter, 1.25 mL of the upper layer of the solution was mixed with 1.25 mL of distilled water and 0.25 mL of FeCl3. The absorbance was read at 700 nm. Higher absorbance of the reaction mixture indicates greater reductive potential.
Assay of DPPH radical scavenging (1, 1-diphenyl-2-picrylhydrazyl) activity: The DPPH radical scavenging activity of the extracts were evaluated according to the method described by Leong and Shui12. Exactly 1 mL of 0.3 mM DPPH prepared in methanol was added to 1 mL of extract of various concentrations (100-500 μg mL–1) and allowed to react at room temperature for 30 min. The reaction mixture was vortexed thoroughly and left in the dark at room temperature for 30 min. The absorbance of the mixture was measured spectrophometrically at 517 nm and the percentage scavenging activity was calculated using the equation as follows:
where, Ac is the absorbance of control and As is the absorbance of the extract.
Assay of 2,2’-azino-bis-(3-ethylbenzothiazoline-6-sulphonic acid) (ABTS) radical scavenging activity: The ABTS radical scavenging activity of the plant extract was determined according to the method of Re et al.13. The stock solutions were of 7 mM ABTS+ and 2.4 mM potassium persulphate. The working solution was then prepared by mixing the two stock solutions in equal quantities and allowing them to react for 16 h at room temperature in the dark. The solution was diluted by mixing 5 mL ABTS+ solution with 145 mL of distilled water to obtain an absorbance of 0.076±0.001 U at 734 nm. Extracts (1 mL) at various concentrations (100-500 μg mL–1) were allowed to react with 1 mL of ABTS+ solution and the absorbance was measured at 734 nm after 30 min using a spectrophotometer and the percentage scavenging activity was calculated using the equation as follows:
where, Ac is the absorbance of control and As is the absorbance of the extract.
Assay of metal chelating activity: The metal chelating activity was determined according to the method of Haro-Vicente et al.14. Extract (1 mL) was added to 100 μL of 1 mM FeSO4.7H2O. The reaction mixture was left at room temperature for 2 min. After which 0.5 mL of 0.5 mM 1, 10-phenanthroline was added and the mixture was incubated for 10 min at room temperature. The absorbance was read at 510 nm. The Fe2+ chelating capacity was calculated thus:
Assay of hydroxyl radical scavenging activity: The hydroxyl radical scavenging activity was measured by studying the competition between deoxyribose and the extracts for hydroxyl radicals generated from the Fe2+/ascorbate/EDTA/ H2O2 system according to the method of Halliwell et al.15. The reaction mixture contained 1 mL (3.0 mM deoxyribose, 0.1 mM EDTA, 2 mM H2O2, 0.1 mM L-Ascorbic acid, 0.1 mM FeCl3.6H2O in 10 mM phosphate buffer, pH 7.4) and various concentrations of the extracts (100-500 μg mL–1). The reaction mixtures were incubated at 37°C for 1 h, followed by the addition of 1 mL of 1% (w/v) TBA (in 0.25 N HCl) and 1.0 mL 10% (w/v) TCA. The reaction mixtures were heated in boiling water bath at 100°C for 20 min and the pink chromogen (malondialdehyde-(TBA) adduct) was extracted into 1.0 mL of butan-1-ol and the absorbance was read at 532 nm against reagent blank. The hydroxyl radical scavenging activity calculated thus:
Experimental design: Twenty five male albino rats weighing 180-200 g were used according to the standard guidelines of the Care and Use of Experimental Animal Resources. The rats were housed in cages under standard laboratory conditions (12 h light/dark cycle, 23±2°C). There were 5 groups of animals distributed thus:
| Group 1 | : | Untreated, unintoxicated group-negative control group (C-) |
| Group 2 | : | Rats intoxicated with CCl4 (1 mL kg–1 b.wt., s.c.) in a 1:1 solution with olive oilintraperitoneally (i.p.)16 after 4 weeks without red wine pretreatment (test control (C+)) |
| Group 3 | : | Rats were pre-treated with alcoholic red wine (500 μL b.wt.) for 4 weeks orally and CCl4 intoxication at the end of the 4th week (RCCl4 group) |
| Group 4 | : | Rats which served as alcoholic red wine master control (500 μL b.wt. for 4 weeks) (RC group) |
| Group 5 | : | Reference group which was treated with standard drug-quercetin (100 μg mL–1, 1 mL kg–1 b.wt.) (Q) |
Urine and blood samples were collected. Rats were anaesthetized by cervical dislocation and sacrificed loss of consciousness. The kidneys were excised, washed in ice cold 1.15% potassium chloride solution, blotted with filter paper and weighed and were homogenized in 5% w/v sodium phosphate buffer (pH 7.4) using a teflon homogenizer. The homogenates were centrifuged 3000 rpm for 20 min. The supernatants were decanted carefully and the resulting supernatant were stored at 4°C before being used for biochemical evaluations.
Biochemical assays
Total protein in kidney homogenate: This was carried out using the manufacturer protocol of Randox Total Protein Kit based on Weichselbaum17. One milliliter of reagent R1; Sodium hydroxide (100 mmol L–1), sodium-potassium tartrate (16 mmol L–1), potassium iodide (15 mmol L–1) and copper II sulphate (6 mmol L–1) was added to 0.02 mL of the test sample, the mixture was incubated at 25°C and the absorbance was then measured against the reagent blank at a wavelength of 546 nm. Total protein concentration was determined as:
Alkaline phosphatase (ALP) activity: Assay of alkaline phosphatase (ALP) activity was carried out according to the procedure provided by Randox Kit Manufacturer, which is based on the method of Englehardt18. Exactly 1.0 mL of the reagent (1 mol L–1, pH 9.8 diethanolamine buffer, 0.5 mmol L–1 MgCl2; substrate: 10 mmol L–1 p-nitrophenolphosphate) was added to 0.02 mL of sample and then mixed. The absorbance was read for 3 min at intervals of 1 min at a wavelength of 405 nm.
The ALP activity was determined using the equation:
U L–1 = 2760×A405 nm min–1
Creatinine clearance rate: The creatinine concentration was determined according to Randox Kit Manufacturer protocol. Urine from rats was collected for 12 h and the volumes of the urine were noted per milliliter. The creatinine concentration in the plasma and the urine were determined using standard biochemical procedures. And the creatinine clearance rates were calculated thus:
Where:
| Ccr | = | Concentration of creatinine (mL min–1) |
| Ucr | = | Urine creatinine (mg mL–1) |
| V | = | Flow rate of urine (mL min–1) |
| Pcr | = | Plasma creatinine (mg mL–1) |
Superoxide dismutase activity (SOD): Kidney SOD activity was assayed by the method of Kakkar et al.19. Reaction mixture contained 1.2 mL of sodium pyrophosphate buffer (0.052 mM, pH 7.0), 0.1 mL of phenazinemethosulphate (PMS) (186 μM), 0.3 mL of Nitro Blue Tetrazolium (NBT) (300 μM). About 0.2 mL of the supernatant obtained after centrifugation (1500×g, 10 min followed by 10,000×g, 15 min) of 10% kidney homogenate was added to reaction mixture. Enzyme reaction was initiated by adding 0.2 mL of NADH (780 μM) and stopped precisely after 1 min by adding 1 mL of glacial acetic acid. Amount of chromogen formed was measured by recording color intensity at 560 nm. Results are expressed as units/mg protein.
Glutathione peroxidase activity (GSH-Px): Glutathione peroxidase (GSH-Px) activity was measured by NADPH oxidation, using a coupled reaction system consisting of glutathione, glutathione reductase and cumenehydroperoxide20. Hundred microliters of enzyme sample were incubated for 5 min with 1.55 mL stock solution (prepared in 50 mM tris buffer, pH 7.6 with 0.1 mM EDTA) containing 0.25 mM GSH, 0.12 mM NADPH and 1 U glutathione reductase. The reaction was initiated by adding 50 μL of cumenehydroperoxide (1 mg mL–1) and the rate of disappearance of NADPH with time was determined by monitoring absorbance at 340 nm. One unit of enzyme activity is defined as the amount of enzyme that transforms one micromole of NADPH-NADP per minute. Results are expressed as units/mg protein.
Catalase activity (CAT): The activity of CAT was measured using its perioxidatic function according to the method of Johansson and Borg21. Fifty microliters potassium phosphate buffer (250 mM, pH 7.0) was incubated with 50 μL methanol and 10 μL hydrogen peroxide (0.27%). The reaction was initiated by addition of 100 μL of enzyme sample with continuous shaking at room temperature (20°C). After 20 min, reaction was terminated by addition of 50 μL of 7.8 M potassium hydroxide. Hundred microliters of purpald (4-amino-3-hydrazino-5-mercapto-1,2,4-triazole, 34.2 mM in 480 mMHCl) was immediately added and the mixture was again incubated for 10 min at 20°C with continuous shaking. Potassium peroxidate (50 μL, 65.2 mM) was added to obtain a colored compound. The absorbance was read at 550 nm in a spectrophotometer. Results are expressed as micromoles of formaldehyde produced/mg protein.
Reduced glutathione (GSH): Reduced glutathione (GSH) level in the kidney was assayed following the method of Ellman22, modified by Hissin and Hilf23. The homogenate (720 μL) was double diluted and 5% TCA was added to it to precipitate the protein content of the homogenate. After centrifugation (10,000×g for 5 min) at 4°C the supernatant was taken, 5,5'-dithiolbis-2-nitrobenzoic acid (DTNB) solution (Ellmam’s reagent) was added to it and the absorbance was measured at 412 nm on a spectrophotometer. A standard graph was drawn using different concentrations of standard GSH solution (1 mg mL–1). With the help of the standard graph, GSH contents in the homogenates of the experimental animals were calculated.
Glutathione-S-transferase (GST): The GST catalyzes the conjugation reaction with glutathione in the first step of mercapturic acid synthesis. The GST activity was measured by the method of Habig et al.24. The reaction mixture contained suitable amount of the enzyme (25 μg of protein in homogenates), 1 mL of KH2PO4 buffer, 0.2 mL of EDTA, 0.1 mL of 1-chloro-2,4-dinitrobenzene (CDNB) and GSH. The reaction was carried out at 37°C and monitored spectrophotometrically by the increase in absorbance of the conjugate of GSH and CDNB at 340 nm. A blank was run in absence of the enzyme. One unit of GST activity is 1 μmol product formation per minute.
The results of replicate readings were pooled and expressed as Mean±Standard Deviation. One way analysis of variance was used to analyze the results and Duncan multiple test was used for the post hoc (DMRT). Statistical Package for Social Science (SPSS) 17.0 for windows was used for the analysis. The significance level was set at p<0.05.
RESULTS
CCl4-induced stress: The profile of biochemical indices such as alkaline phosphatase (ALP) activity (Fig. 1a), creatinine clearance (Fig. 1b) and Total Protein (TP) (Fig. 1c) were used in assessing the anti-toxic properties of the red wine extracts on the kidney.
Figure 1d-h revealed the effects of red wine on biochemical variables in the kidney homogenates of all experimental animal groups. The CCl4 intoxication markedly decreased antioxidants enzymes as well as glutathione levels and this could be as a result of oxidative stress in CCl4-intoxicated rats. There was clear evidence that CCl4-induced kidney injury was associated with free radical injury and oxidative stress as revealed by the antioxidants activities. Effects of CCl4, CCl4+red wine and alcoholic red wine treatments on renal tissue’s GSH-Px, SOD, CAT, GSH and GST are presented in Fig. 1d-h. The renal antioxidant activity of GSH-Px, SOD, CAT, GSH and GST significantly decreased (p<0.05) in the CCl4-intoxicated group of rats. The normal control group (C-) maintained optimal activity of the antioxidants studied. Administration of alcoholic red wine significantly (p<0.05) decreased the activities of the endogenous antioxidant enzymes. Alcoholic red wine proved significantly in restoring the altered activity of antioxidant enzymes like SOD, GSH-Px, CAT, GST and GSH towards their normal values in the kidney homogenates due to the intoxication.
DISCUSSION
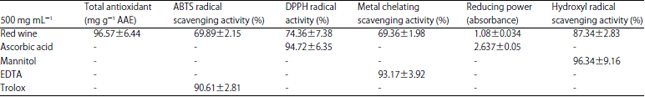
The evaluations of the antioxidant activities of the red wine extract was imperative to evaluating its possible in vivo modulation of the antioxidant status in animal model. Table 1 revealed the activities of red wine extract competitively with standard antioxidant compounds. It was obvious from the result that red wine extract had metal chelating activity as well as radicals scavenging abilities. The result also established red wine as a proton-donator, a reductive activity peculiar with good antioxidant compounds.
In general, a number of chemicals including various environmental toxicants and even clinically useful drugs can cause severe cellular damages in different organs of the body through metabolic activation to highly reactive substances such as free radicals25.
Carbon tetrachloride (CCl4) is a potent hepatotoxin and is rapidly bio-transformed by cytochrome P-450 to trichloro methyl (CCl3) radical and Cl16. In the liver and kidneys after the initiation of lipid peroxidation, CCl3 react with oxygen of cellular proteins and lipids producing a trichloro methyl peroxyl radical, which attacks rapidly lipid membrane of endoplasmic reticulum. Living cells continuously react with oxygen leading to the production of free radicals16. The reactive oxygen species are predisposing factors for many diseases including cancer, arthritis, aging and immunodeficiency diseases26. The damage caused by free radicals is partly ameliorated by a large group of protective agents called antioxidants that react with free radicals stopping the damage they cause. These radicals are capable of initiating a chain of lipid peroxidation reactions by abstracting hydrogen from polyunsaturated fatty acids (PUFA).
| Table 1: | In vitro antioxidant abilities of the 500 mg mL–1 extract of red wine with standard antioxidant compounds for each assay |
 | |
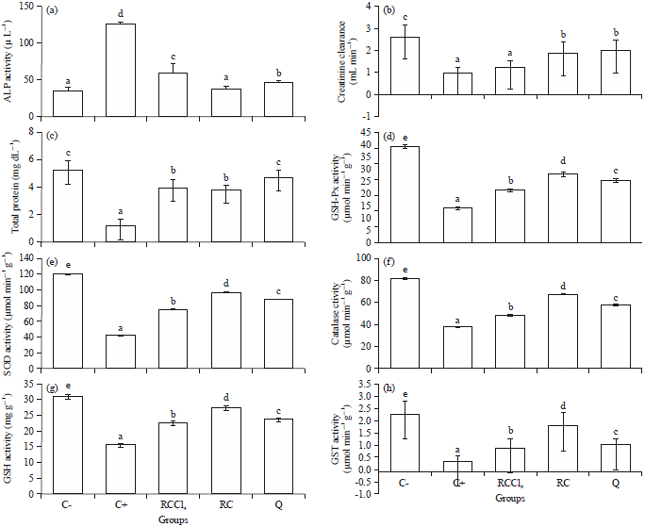 | |
| Fig. 1(a-h): | Effects of extracts of red wine on kidney (a) ALP activity, (b) Creatinine clearance, (c) Total protein, (d) GSH-Px activity, (e) Superoxide dismutase activity, (f) Catalase activity, (g) GSH activity and (h) GST activity. Values are expressed as mean (n = 5). Values with different superscript are significantly different (p<0.05) |
Peroxidation of lipids, particularly those containing PUFA can dramatically change the properties of biological membranes, resulting in severe cell damage and play a significant role in pathogenesis of diseases27.
The ALP values in the kidney revealed that the alcoholized red wine had ameliorating effect on the CCl4-induced stress. Alkaline phosphatases are a group of enzymes found primarily in the liver (isoenzyme ALP-1) and bone (isoenzyme ALP-2). Alkaline phosphatases act by splitting off phosphorus (an acidic mineral) creating an alkaline pH. The result (Fig. 1a) also revealed that the red wine consumption at this concentration does not show toxicological stress as an implication for the presence of alcohol (11%).
The creatinine clearance rate (Fig. 1b) of the red wine extract used for pretreating CCl4 induction ameliorated the effect of the assault on the kidney, although significantly lower than treatment with quercetin. There was no significant difference between the red wine master and the quercetin treated group. The alcoholic red wine facilitated the removal of creatinine from the circulatory system compared to the urinary flow. The creatinine clearance rate, which defines the activities in the glomerular filtrate, ascribed the absence of tumor in the glomerulus to the administered alcoholic red wine. This is in agreement with the work of Al-Sowayan and Mousa28 on ameliorative effect of olive leaf extract on carbon tetrachloride-induced nephrotoxicity in rats.
The red wine showed ability to prevent oxidative damage to the kidney. Figure 1c revealed that there was significant difference between red wine treated group and the test control group (C+). Similar result was indicated for the wine master group, which when compared to the negative control had stressors responsible for endogenous degradation of these nephron proteins. The protective activity was primarily due to the ability of the polyphenolic contents of the red wine to scavenge free radical; protonating them, chelating the metallic ions or synergizing the endogenous antioxidants against free radicals. In all of these, the objectives remain the protection of biomolecules, such as nucleic acid, proteins and lipid from oxidative damage from free radical (CCl3 in this case). Another mechanism responsible for this paradox may be related to an increase of marine omega-3 fatty acids concentration in blood (an effect they termed "Fish-like effect" of wine), since long chain omega-3 fatty acids are known to protect against cardiovascular complications to reduce inflammation and to limit severe cardiac arrhythmias29,30.
In vivo studies indicated that CCl4 enhanced lipid peroxidation, reduced renal microsomal NADPH cytochrome P450 and renal reduced/oxidized glutathione ratio (GSH/GSSG) in kidney cortex as well as renal microsomes and mitochondria31. Evidence suggested that various enzymatic and non-enzymatic systems have been developed by mammalian cells to cope with ROS and other free radicals32. However, when a condition of oxidative stress establishes, the defense capacities against ROS becomes insufficient33. The ROS also affects the antioxidant defense mechanisms, by reducing the intracellular concentration of GSH and decreases the activity of SOD, CAT and GSH-Px. It has also been observed to decrease the detoxification system produced by GST34. In the kidney GST is a cystolic protein, highly specific for the cells of the proximal tubules.
From the present study, it was observed that CCl4 intoxicated group significantly decreased in SOD, CAT, GSH-Px and GST activities, depleted the GSH content in kidney. It has been reported that SOD, CAT, GSH-Px and GST constitute a mutually supportive team of defense against ROS35. The decreased activity of SOD in kidney in CCl4‾intoxicated rats may be due to the inactivation of the antioxidative enzymes. This would cause an increased accumulation of superoxide radicals, which could further cause deleterious effects. The GST bind to liophilic compounds and acts as an enzyme for GSH conjugation reactions36. Decreased GST activity during CCl4 toxicity might be due to the decreased availability of GSH resulted during the enhanced cellular oxidation.
Administration of alcoholic red wine prior to CCl4 intoxication activated the antioxidant machineries of the kidney as revealed from enhanced levels of SOD, CAT, GSH-Px and GST activities, increased GSH content. However, the reduction in the activities of the endogenous antioxidant systems in alcoholic red wine master group (RC) compared with the normal control, could be as a result of radical generating effect of alcohol which was capable of causing depletion in the antioxidant status, thus resulting in oxidative stress.
CONCLUSION
The antioxidant activities of the extract red wine showed scavenging activities of free radicals as well as ability to chelate metallic ions, thus could be a dietary source of bioactive compounds. This study described the renaloprotective activities of the alcoholic red wine against CCl4 intoxication. Despite the adaptogenic activity of the wine, the effect of the alcoholic content cannot be neglected, limiting its dietary recommendation as a result of the alcoholic content inexcessive consumption. However, further study should demonstrate comparative study on dealcoholize red wine as well as histopathological studies on kidney functions.
REFERENCES
- Burns, J., P.T. Gardner, J. O'Neil, S. Crawford and I. Morecroft et al., 2000. Relationship among antioxidant activity, vasodilation capacity and phenolic content of red wines. J. Agric. Food Chem., 48: 220-230.
CrossRefDirect Link - Mattivi, F., R. Guzzon, U. Vrhovsek, M. Stefanini and R. Velasco, 2006. Metabolite profiling of grape: Flavonols and anthocyanins. J. Agric. Food Chem., 54: 7692-7702.
CrossRefDirect Link - St. Leger, A.S., A.L. Cochrane and F. Moore, 1979. Factors associated with cardiac mortality in developed countries with particular reference to the consumption of wine. Lancet, 313: 1017-1020.
CrossRefDirect Link - Renaud, S. and M. de Lorgeril, 1992. Wine, alcohol, platelets and the French paradox for coronary heart disease. Lancet, 339: 1523-1526.
CrossRefPubMedDirect Link - Di Castelnuovo, A., L. Iacoviello and G. De Gaetano, 2003. Alcohol and coronary heart disease. N. Engl. J.Med., 348: 1719-1722.
CrossRefDirect Link - Slater, T.F. and B.C. Sawyer, 1971. The stimulatory effects of carbon tetrachloride and other halogenoalkanes on peroxidative reactions in rat liver fractions in vitro. General features of the systems used. Biochem. J., 123: 805-814.
CrossRefDirect Link - Sakata, T., N. Watanabe, N. Hobara and H. Nagashima, 1987. Chronic liver injury in rats by carbon tetrachloride inhalation. Bull. Environ. Contam. Toxicol., 38: 959-961.
CrossRefDirect Link - Venkov, C.D., P.R. Myers, M.A. Tanner, M. Su and D.E. Vaughan, 1999. Ethanol increases endothelial nitric oxide production through modulation of nitric oxide synthase expression. Thromb. Haemost., 81: 638-642.
PubMedDirect Link - Leikert, J.F., T.R. Rathel, P. Wohlfart, V. Cheynier, A.M. Vollmar and V.M. Dirsch, 2002. Red wine polyphenols enhance endothelial nitric oxide synthase expression and subsequent nitric oxide release from endothelial cells. Circulation, 106: 1614-1617.
CrossRefPubMedDirect Link - Oyaizu, M., 1986. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Dietetics, 44: 307-315.
CrossRefDirect Link - Leong, L.P. and G. Shui, 2002. An investigation of antioxidant capacity of fruits in Singapore markets. Food Chem., 76: 69-75.
CrossRefDirect Link - Re, R., N. Pellegrini, A. Proteggente, A. Pannala, M. Yang and C. Rice-Evans, 1999. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biol. Med., 26: 1231-1237.
CrossRefDirect Link - Haro-Vicente, J.F., C. Martinez-Gracia and G. Ros, 2006. Optimisation of in vitro measurement of available iron from different fortificants in citric fruit juices. Food Chem., 98: 639-648.
CrossRefDirect Link - Halliwell, B., J.M.C. Gutteridge and O.I. Aruoma, 1987. The deoxyribose method: A simple test-tube assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem., 165: 215-219.
CrossRefDirect Link - Adewole, S.O., A.A. Salako, O.W. Doherty and T. Naicker, 2007. Effect of melatonin on carbon tetrachloride-induced kidney injury in Wistar rats. Afr. J. Biomed. Res., 10: 153-164.
CrossRefDirect Link - Weichselbaum, T.E., 1946. An accurate and rapid method for the determination of proteins in small amounts of blood serum and plasma. Am. J. Clin. Pathol., 10: 40-49.
PubMedDirect Link - Kakkar, P., B. Das and P.N. Viswanathan, 1984. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys., 21: 130-132.
PubMedDirect Link - Tappel, A.L., 1978. Glutathione peroxidase and hydroperoxides. Methods Enzymol., 52: 506-513.
CrossRefPubMedDirect Link - Johansson, L.H. and L.A. Borg, 1988. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem., 174: 331-336.
CrossRefPubMedDirect Link - Ellman, G.L., 1959. Tissue sulfhydryl groups. Arch. Biochem. Biophys., 82: 70-77.
CrossRefPubMedDirect Link - Hissin, P.J. and R. Hilf, 1976. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem., 74: 214-226.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Noguchi, T., K.L. Fong, E.K. Lai, S.S. Alexander and M.M. King et al., 1982. Specificity of a phenobarbital-induced cytochrome P-450 for metabolism of carbon tetrachloride to the trichloromethyl radical. Biochem. Pharmacol., 31: 615-624.
CrossRefPubMedDirect Link - Al-Fartosi, K.G., A. Majid, M.A. Auda and M.H. Hussein, 2012. The role of camel's milk against some oxidant-antioxidant markers of male rats treated with CCl4. Int. J. Res. Pharmaceut. Biomed. Sci., 3: 385-389.
Direct Link - Aleynik, S.I., M.A. Leo, X. Ma, M.K. Aleynik and C.S. Lieber, 1997. Polyenylphosphatidylcholine prevents carbon tetrachloride-induced lipid peroxidation while it attenuates liver fibrosis. J. Hepatol., 27: 554-561.
CrossRefDirect Link - Al-Sowayan, N.S. and H.M. Mousa, 2014. Ameliorative effect of olive leaf extract on carbon tetrachloride-induced nephrotoxicity in rats. Life Sci. J., 11: 238-242.
Direct Link - Guiraud, A., M. De Lorgeril, S. Zeghichi, F. Laporte and P. Salen et al., 2008. Interactions of ethanol drinking with n-3 fatty acids in rats: Potential consequences for the cardiovascular system. Br. J. Nutr., 100: 1237-1244.
CrossRefDirect Link - Gu, X., L. Creasy, A. Kester and M. Zeece, 1999. Capillary electrophoretic determination of resveratrol in wines. J. Agric. Food Chem., 47: 3223-3227.
CrossRefDirect Link - Walker, G., D. Kunz, W. Pignat, I. Wiesenberg, H. Van den Bosch and J. Pfeilschifter, 1996. Tetranactin inhibits interleukin 1β and cAMP induction of group II phospholipase A2 in rat renal mesangial cells. Eur. J. Pharmacol., 306: 265-270.
CrossRefDirect Link - Recknagel, R.O., E.A. Glende, Jr., J.A. Dolak and R.L. Waller, 1989. Mechanisms of carbon tetrachloride toxicity. Pharmacol. Ther., 43: 139-154.
CrossRefPubMedDirect Link - Yamamoto, Y. and S. Yamashita, 1999. Plasma Ubiquinone to ubiquinol ratio in patients with hepatitis, cirrhosis and hepatoma and in patients treated with percutaneous transluminal coronary reperfusion. BioFactors, 9: 241-246.
CrossRefPubMedDirect Link - Bandyopadhyay, U., D. Das and R.K. Banerjee, 1999. Reactive oxygen species: Oxidative damage and pathogenesis. Curr. Sci., 77: 658-666.
Direct Link - Anandan, R., R.D. Rekha and T. Devaki, 1999. Protective effect of Picrorhiza kurroa on mitochondrial glutathione antioxidant system in D-galactosamine-induced hepatitis in rats. Curr. Sci., 76: 1543-1545.
Direct Link