
E.A. Al-Omireeni
Department of Biochemistry, College of Science, King Saud University, P.O. Box 2455, Riyadh-11451, Saudi Arabia
N.J. Siddiqi
Department of Biochemistry, College of Science, King Saud University, P.O. Box 2455, Riyadh-11451, Saudi Arabia
A.S. Alhomida
Department of Biochemistry, College of Science, King Saud University, P.O. Box 2455, Riyadh-11451, Saudi Arabia
Kidney Research Journal
Year: 2011 | Volume: 1 | Issue: 1 | Page No.: 33-40







ABSTRACT
The present study was carried out to study the acute toxicity of various doses of sodium fluoride (NaF) on different hydroxyproline (HyP) fractions in rat kidneys. Five groups of rats were studied: (1) control rats, (2) rats divided into four subgroups according to the dose of NaF viz., rats were injected with a single intraperitoneal dose of 5, 10, 20 and 30 mg of Naf kg-1 b.wt./24 h. Results showed that protein-bound HyP fraction was most resistant to the effect of NaF. Doses of NaF caused significant changes (p<0.05) in other HyP fractions viz., free and peptide-bound HyP. Among the soluble and insoluble collagen HyP, it was soluble collagen Hyp which was significantly altered by NaF treatment (p<0.05). NaF treatment also caused significant (p<0.05) decrease in the protein content of rat kidneys. It is concluded that NaF treatment to rats caused a disturbance in connective tissue matrix of the kidneys in rats.
PDF Abstract XML References Citation
How to cite this article
E.A. Al-Omireeni, N.J. Siddiqi and A.S. Alhomida, 2011. Effect of Different Doses of Sodium Fluoride on Various Hydroxyproline Fractions in Rat Kidneys. Kidney Research Journal, 1: 33-40.
DOI: 10.3923/krj.2011.33.40
URL: https://scialert.net/abstract/?doi=krj.2011.33.40
DOI: 10.3923/krj.2011.33.40
URL: https://scialert.net/abstract/?doi=krj.2011.33.40
INTRODUCTION
Fluorine compounds are used in various areas of medicine particularly dentistry, agriculture and industry. Fluoride has the potential to increase skeletal mass to a great extent, yet it has proven difficult to translate this into therapeutic benefit for patients with low bone mass in diseases like osteoporosis (Kleerekoper, 1996). This is in part due to toxic actions of fluoride ion on skeletal mineralization, impairment of the normal processes of bone resorption and fluoride-induced decreases in strength per unit of bone (mass or volume) (Kleerekoper, 1996). Other toxic effects of excessive exposure to fluoride include hyperemia, cerebral edema and degeneration of liver and kidneys (Machoy-Mokrzynska, 2004). Fluoride levels in surface waters vary according to nature of the rocks in the area. Surface water concentrations range from 0.01 to 0.3 mg L-1. The concentration in seawater is higher ranging from 1.2 to 1.5 mg L-1. Inspite of its toxic effects, fluoride continues to be used in different products and acute intoxications are relatively rare. The more common finding is chronic intoxication which is due to the universal presence of fluorine compounds in the environment. Fluorine compounds have been shown to act on organic part of supporting tissues including collagen and cells of the connective tissue (Machoy-Mokrzynska, 2004). One of the most commonly used marker of collagen breakdown/ degradation is hydroxyproline. The hydroxyproline (HyP) is a post translactional product of proline hydroxylation catalyzed by the enzyme prolylhydroxylase (EC 1.14.11.2) (Pihlajaneimi et al., 1991). The occurrence of this amino acid is thought to be confined exclusively to collagen, where it is present in the Y position of the Gly-X-Y repeating tripeptide (Nemethy and Scheraga, 1986). Consequently, the presence of hydroxyproline (HyP) in tissues or serum can be used as a measure of collagen or collagen degradation products (Reddy and Enwemeka, 1996).
The kidneys excrete waste products of metabolism and play an important role in maintaining the homeostasis by regulating the body water and solute balance. In addition to the excretory function the kidneys also have an endocrine function producing hormones like renin, erythropoietin etc. In our earlier studies (Siddiqi and Alhomida, 2005, 2006), we have shown that mercuric chloride treatment to rats damages the collagen which is reflected by increased levels of Hyp in serum and an increased excretion of Hyp in urine. Sodium fluoride has been shown to cause several biochemical changes in rat kidneys, the most recent being inhibition of the enzyme arginase (Tormanen, 2003). The most commonly used medium for studying fluoride toxicity is urine. Acute exposure to high doses of fluoride damages renal tissue and causes renal dysfunction (Zabulyte et al., 2007). The present study was carried out to study the effect of sodium fluoride treatment on various Hyp fractions in rat kidneys.
MATERIALS AND METHODS
Chemicals: Chloramine-T, p-dimethylaminobenzaldeyde (Ehrlich’s reagent), L-hydroxyproline, sodium acetate, citric acid, perchloric acid, n-propanol, sodium hydroxide and acetic acid were purchased from Sigma Chemical Company, St Louis, MO, USA. Glass distilled water was used throughout the study.
Study: This study was carried out at the Biochemistry Department Research Laboratory, College of Science, Sciences and Medical Studies Department for Women Students, King Saud University, Riyadh between February 2008 and May 2009.
Animal care: Healthy adult male Wistar rats weighting 150-200 g (four to six weeks old) were obtained from Breeding Laboratory, King Saud University, Riyadh, Saudi Arabia. The animals were labeled by identifying ear notches, housed in clean cages and placed in the animal care room. Ethical guidelines for animal care were followed.
Dose-response of sodium fluoride on hyp fractions in rat kidneys: Rats were allowed free access to food (Purina rodent chow) and tap water for one day. After one day, rats were divided into different groups. The following groups were studied: (1) normal rats (Control group, n = 4-6 rats); rats were divided into four subgroups according to the dose of NaF viz., rats were injected with a single dose ip of 5, 10, 20 and 30 mg of Naf kg-1 b.wt./24 h (sodium fluoride treated group, n = 5-6 rats).
Preparation of the sample: The animals were killed 24 h after the NaF injection. The animals were killed by carbondioxide asphyxiation. The kidneys were dissected out, cleared of adhering tissues and weighed. The kidneys were then homogenized in normal saline (10% w/v) and the homogenate was used for HyP determination as described below.
Extraction of free, peptide- and protein-bound hydroxyproline: Free and protein-bound HyP was extracted by the method of Varghese et al. (1981) with slight modification. Briefly, 0.5 mL of the homogenate was treated with 3x2 mL portion of re-rectified absolute alcohol and centrifuged at 600 g for 10 min. The supernatants were pooled and evaporated to dryness. The residue was dissolved in suitable amount of distilled water and an aliquot of the extract was used for estimation of free HyP. The peptide-bound HyP was determined after alkaline hydrolysis of the ethanol extractable fraction. The pellets were dissolved in distilled water and an aliquot of the extract was used for determination of protein-bound HyP. The free HyP fraction was measured on an aliquot of the ethanol extracted residue without alkali hydrolysis, whereas the peptide-bound HyP was measured after alkaline hydrolysis. The precipitate obtained on ethanol treatment of the homogenate was subjected to alkali hydrolysis to determine protein-bound HyP. Further details about the extraction of HyP fractions were described previously (Siddiqi et al., 2000). HyP was determined in different fractions.
Extraction of soluble- and insoluble-collagen HyP: Soluble- and insoluble-collagen HyP was extracted by the method of Kivirikko et al. (1965). Briefly, the tissue samples were homogenized (4 mL g-1 tissue) in a cold 0.45 M sodium chloride. The homogenate was extracted at 4°C for 24 h with occasional stirring, followed by centrifugation at 13000 rpm for 1 h. The supernatants obtained were precipitated with 4 volumes of a cold ethanol and after centrifugation twice with 80% ethanol, twice with absolute alcohol, twice with ether and twice with warm ethanol-ether (1:2). The residues were gelatinized with distilled water at 120°C for 3 h and after filtration a sample of gelatine solution was used for soluble-collagen HyP estimation as described below.
The precipitates obtained after the above centrifugation were washed 3 times with 0.45 M NaCl and twice with distilled water, after which they were extracted absolute ethanol, ether and ethanol-ether and gelatinized as above. A sample of gelatine solution was used for insoluble-collagen HyP estimation.
Determination of hydroxyproline concentration: HyP was measured by the modified alkaline hydrolysis method of Reddy and Enwemeka (1996). Briefly, to an aliquot of the sample was added into NaOH (2 N final concentration) and the aliquot was hydrolyzed by heating in a boiling water bath for about 3-4 h. An aliquot of 56 mM chloramine-T reagent was added to the hydrolyzed sample and oxidation was allowed to proceed at room temperature for 25 min. Then an aliquot of 1 M Ehrlich s reagent (p-dimethylaminobenzaldehyde) was added to the oxidized sample and the chromophore was developed by incubating the samples at 65°C for 20 min. The absorbance was read at 550 nm using an Ultrospec 2000 UV/visible spectrophotometer (Pharmcia Biotech Ltd., Science Park, Cambridge, England). The HyP concentration in the samples was calculated from the standard curve of HyP. Further details about the optimization, linearity, specificity, precision and reproducibility of the method were described previously (Siddiqi et al., 2000).
Statistical analysis: Each sample was run in duplicate. The HyP content was expressed as Mean±SD μg g-1 wet tissue, for n = 5 rats. Serum HyP levels between groups were compared using one way ANOVA analysis followed by Tukey’s for multiple comparison test. Values were considered significant if p<0.05. Statistical analysis was performed by means of InStat® package for personal computers (GraphPad TM Software, Inc., San Diego, USA).
RESULTS AND DISCUSSION
Table 1 shows the effect of various doses of sodium fluoride on different hydroxyproline fractions in rat kidneys. NaF doses of 10 and 20 mg kg-1 b.wt. caused a significant decrease of 62 and 25%, respectively (p<0.05) in free HyP when compared to control group.
| Table 1: | Effect of sodium fluoride treatment on different hydroxyproline fractions in rat kidneys |
 | |
| Values are expressed as mean±SD gram kidney weight-1 ( 4-6 rats/group), ns: Non significant as compared to control group (Tukey’s multiple comparision test); *p<0.05 compared to control group (Tukey’s multiple comparision test); **p<0.01 compared to control group (Tukey’s multiple comparision test); ***p<0.001 compared to control group (Tukey’s multiple comparision test). Animals were injected with sodium fluoride through intraperitoneal route. Each group consisted of 4-6 animals. Rats were sacrificed 24 h after the treatment | |
| Table 2: | Effect of sodium fluoride treatment on protein content in rat kidneys |
 | |
| Values are expressed as mean±SD gram kidney weight-1 (6 rats/group), ns: Non significant as compared to control group (Tukey’s multiple comparision test); *p<0.05 compared to control group (Tukey’s multiple comparision test); **p<0.01 compared to control group (Tukey’s multiple comparision test); ***p<0.001 compared to control group (Tukey’s multiple comparision test). Animals were injected with sodium fluoride through intraperitoneal route. Rats were sacrificed 24 h after the treatment | |
Similarly NaF doses of 10, 20 and 30 mg kg-1 b.wt. caused a significant decrease of peptide-bound HyP by 86, 83 and 88% respectively when compared to control rats (p<0.001). Protein bound HyP was not affected by doses of NaF except an initial dose of 5 mg kg-1 b.wt. Naf dose of 5 mg kg-1 b.wt. caused an initial increase of 163% (p<0.001) in protein bound HyP when compared to control group of rats. NaF doses of 10, 20 and 30 mg kg-1 b.wt. caused a significant decrease of 78, 76 and 79% respectively in total HyP when compared to control group (p<0.001).
Table 2 shows the effect of sodium fluoride treatment on protein levels in rat kidneys. Results show that NaF treatment caused significant (p<0.05) decrease in the protein content in rat kidneys. Doses of 5, 10, 20 and 30 mg kg-1 b.wt. of NaF caused a decrease in protein content by 163% (p<0.001), 99% (p<0.05), 4 (p<0.05) and 65% (p<0.001) respectively when compared to the control group of rats.
Figure 1 shows the effect of different doses of NaF on total collagen content in rat kidneys. NaF doses of 10, 20 and 30 mg kg-1 b.wt. caused a significant decrease of 78, 76 and 79%, respectively in total collagen when compared to control group (p<0.001).
Figure 2 shows the effect of sodium fluoride on insoluble collagen hydroxyproline in rat kidneys. All the doses of NaF used caused no significant change in insoluble collagen hydroxyproline in rat kidneys when compared to control group (p<0.05).
Figure 3 shows the effect of sodium fluoride on soluble collagen hydroxyproline in rat kidneys. NaF at doses 10 and 20 mg kg-1 b.wt. caused a decrease in kidney soluble collagen Hyp by 54% (p<0.01) and 44% (p<0.05), respectively when compared to control rats.
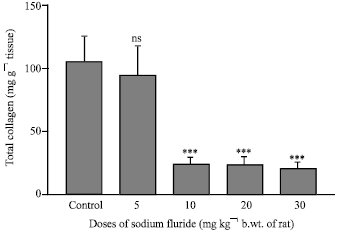 | |
| Fig. 1: | Effect of sodium fluoride on total collagen in rat kidneys. Rats were given sodium fluoride through intraperitoneal route. The animals were sacrificed 24 h after the treatment. Values are expressed as mean±SD gram kidney weight-1 (4-6 rats/group) nsnon significant as compared to control group (Tukey’s multiple comparision test); ***p<0.001 compared to control group (Tukey’s multiple comparision test) |
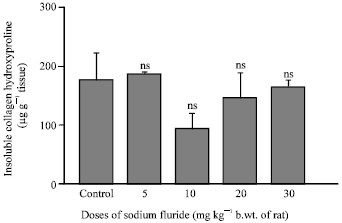 | |
| Fig. 2: | Effect of sodium fluoride on insoluble collagen hydroxyproline in rat kidneys. Rats were given sodium fluoride through intraperitoneal route. The animals were sacrificed 24 h after the treatment. Values are expressed as mean±SD gram kidney weight-1 (4-6 rats/group) nsnon significant as compared to control group (Tukey’s multiple comparision test) |
Fluoride is useful in preventing dental caries and in the treatment of osteoporosis. But excessive intake of fluoride can be toxic. The first noticeable signs of excessive exposure to fluoride is discolouration of the enamel. Abnormalities in mineralization processes affect by and large the osteoarticular system and are associated with changes in the density and structure of the bone presenting as irregular mineralization of the osteoid. Fluorine compounds also act on the organic part of supporting tissues, including collagen and other proteins and on cells of the connective tissue. These interactions reduce the content of collagen proteins, modify the structure and regularity of collagen fibers and induce mineralization of collagen (Machoy-Mokrzynska, 2004).
The kidneys are paired bean-shaped organs located on either side of the spinal column. The kidneys perform a variety of functions for the body, the most important being removal of unwanted substances (waste and surplus) from the plasma, homeostasis of the body’s water, electrolyte and acid/base status and participation in endocrine regulation.
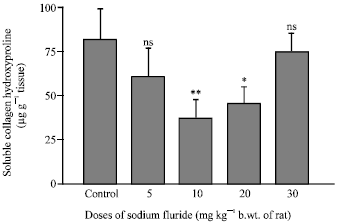 | |
| Fig. 3: | Effect of sodium fluoride on soluble collagen hydroxyproline in rat kidneys. Rats were given sodium fluoride through intraperitoneal route. The animals were sacrificed 24 h after the treatment. Values are expressed as mean±SD gram kidney weight-1 (4-6 rats/group) nsnon significant as compared to control group (Tukey’s multiple comparision test); *p<0.05 compared to control group (Tukey’s multiple comparision test) |
The amount of collagen in the kidney depends on factors like the species of the animal, its age and the presence of disease. In general collagen forms only a small proportion of the renal mass about 2% of the dry weight of renal cortex of adult rats (Weiss and Jayson, 1982) and this may be due to the presence of an active collagenolytic mechanism in the kidney (Weiss and Jayson, 1982). Nevertheless, the collagen is of great physiological importance as a support for the renal parenchyma and as a component of the basement membrane.
In the present study various doses of NaF were used to study its acute toxic effects on various Hyp fractions and total collagen in rat kidneys. NaF at a dose of 5 mg kg-1 b.wt. caused a significant change only in the protein-bound HyP while other HyP fractions remained significantly unchanged. On the other hand the protein-bound HyP seemed to be the only HyP fraction which remained resistant to significant change by other doses of HyP. This may be due to an attempt by the body to overcome the initial NaF assault in the processing of adjusting to the acute exposure to NaF. Studies of Siddiqi and Pandey (1999) have shown that during P.yoelii infection in mice, initially there is an increase in the activities of superoxide dismutase and catalase followed by there decrease at higher levels of parasitemia. Other doses of NaF used viz., 10, 20 and 30 mg kg-1 b.wt. caused a significant decrease in peptide- bound Hyp. Hyp is excreted by the kidneys as small peptides (Adams and Frank, 1980). In the present study NaF treatment caused a decrease in total collagen. The degraded collagen may be excreted in the form of peptide-bound Hyp causing a decrease in peptide-bound HyP fraction. These results are in agreement with the studies of Sharma (1982a) who have demonstrated that fluoride interferes with the collagen biosynthesis resulting in decreased collagen content (in terms of hydroxyproline). Therefore the decrease in collagen content of kidneys of NaF treated animals may be either due to decreased synthesis or increased degradation by collagenase (Machoy-Mokrzynska, 2004). 10 and 20 mg kg-1 b.wt. doses of NaF caused a decrease in free HyP fractions in the kidneys. The pool of free HyP has complex origin. It can arise from mature collagen, newly synthesized collagen, from dietary collagen or from the propeptides of collagen (Adams and Frank, 1980).
Earlier studies of Prockop (1964) have shown that there exists at least 3 pools of body collagen with half lives of about 1 day, 5 days and 50-100 days. The first two of these represent the soluble collagen fractions, i.e. fractions containing collagen not yet aggregated to insoluble-collagen fiber and the third pool, the insoluble collagen. In the present study the soluble collagen Hyp appeared to be susceptible to degradation by NaF. This may be due to the fact that it has still not aggregated to form insoluble collagen fibers. Studies of Sharma (1982b) have shown that fluoride interferes with collagen crosslinking. Studies by the same author have also demonstrated that NaF interferes with maturation and normal metabolism of tissue collagen.
NaF treatment also caused a significant decrease in the total protein content of the kidneys. Proteins are kept in their three dimensional structure by weak hydrogen bonds between adjacent proteins. Emsley et al. (1981), found that fluoride disrupts this hydrogen bonding within proteins by virtue of an unusually strong bond between fluoride ion and the NH group of amides.
In conclusion NaF treatment to rats caused a disturbance in connective tissue matrix of the kidneys.
ACKNOWLEDGMENTS
The authors would like to thank the Research Center, Sciences and Medical Studies Department for Women Students, King Saud University, Riyadh and King Abdul Aziz City for Science and Technology, Riyadh for financial support to EAA.
REFERENCES
- Adams, E. and L. Frank, 1980. Metabolism of proline and hydroxyprolines. Annu. Rev. Biochem., 49: 1005-1061.
CrossRefPubMedDirect Link - Emsley, J., J.J. Deborah, M.M. Jack, E.O. Richard and A.W. Roland, 1981. An unexpectedly strong bond: Ab initio calculations and spectroscopic studies of amide fluoride systems. J. Am. Chem. Soc., 103: 24-28.
CrossRefDirect Link - Kleerekoper, M., 1996. Fluoride and the skeleton. Crit. Rev. Clin. Lab. Sci., 33: 139-161.
CrossRefPubMedDirect Link - Machoy-Mokrzynska, A., 2004. Fluorine as a factor in premature aging. Ann. Acad. Med. Stetin., 50: 9-13.
PubMedDirect Link - Nemethy, G. and H.A. Scheraga, 1986. Stabilization of collagen fibrils by hydroxyproline. Biochemistry, 25: 3184-3188.
CrossRefPubMedDirect Link - Prockop, D.J., 1964. Isotopic studies on collagen degradation and the urinary excretion of hydroxyproline. J. Clin. Invest., 43: 453-460.
CrossRefPubMedDirect Link - Reddy, K. and C.S. Enwemeka, 1996. A simplified method for the analysis of hydroxyproline in biological tissues. Clin. Biochem., 29: 225-229.
CrossRefDirect Link - Sharma, Y.D., 1982. Effect of sodium fluoride on collagen cross-link precursors. Toxicol. Lett., 10: 97-100.
PubMedDirect Link - Sharma, Y.D., 1982. Variations in the metabolism and maturation of collagen after fluoride ingestion. Biochim. Biophys. Acta., 715: 137-141.
PubMedDirect Link - Siddiqi, N.J., A.A. Al-Jafari and A.S. Alhomida, 2000. Investigation of total, free, peptide-bound, protein-bound, soluble and insoluble-collagen hydroxyproline content in tissues from the Arabian camel (Camelus dromadarius). Cell. Biochem. Funct., 18: 243-248.
Direct Link - Tormanen, C.D., 2003. Substrate inhibition of rat liver and kidney arginase with fluoride. J. Inorg. Biochem., 93: 243-246.
CrossRefPubMedDirect Link - Zabulyte, D., S. Uleckiene, J. Kalibatas, A.P. Viciene, N. Jascaniniene and M. Stosik, 2007. Experimental studies on effect of sodium fluoride and nitrate on biochemical parameters in rats. Bull. Vet. Inst. Pulawy., 51: 79-82.
Direct Link