
Hannah N. Tanner
Department of Biology, Eastern Kentucky University, Richmond 40475, Kentucky, United States of America
Devan E. Atkins
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Kimberly L. Bosh
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Grace W. Breakfield
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Sydney E. Daniels
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Makayla J. Devore
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Hailey E. Fite
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Landys Z. Guo
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Danielle K.J. Henry
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Alana K. Kaffenberger
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Katherine S. Manning
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Tatum E. Mowery
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Cecilia L. Pankau
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Malina E. Serrano
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Yamaan Shakhashiro
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Ruth A. Ward
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Aubrey H. Wehry
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
Robin L. Cooper
Department of Biology, University of Kentucky, Lexington 40506, Kentucky, United States of America
LiveDNA: 1.523
Journal of Pharmacology and Toxicology
Year: 2022 | Volume: 17 | Issue: 1 | Page No.: 14-27







ABSTRACT
Background and Objective: Pharmacological agents 4-aminopyridine (4-AP) and tetraethylammonium chloride (TEA) block different subsets of voltage-gated potassium (K+) channels. The potassium channels in the chordotonal organs in crab limbs are sensitive to both blockers. These organs serve as a model of proprioceptive sensation and have rapidly and slowly-adapting sensory neurons. Since 4-AP is used clinically in the treatment of amyotrophic lateral sclerosis and multiple sclerosis, a better understanding of its action on proprioceptive models will increase our knowledge of the potential effects in mammalian systems. Materials and Methods: To assess the action of these blockers on the function of proprioceptive sensory neurons, the neurons were evoked by movements associated with the joint while applying these compounds individually or in combination. The model organism was the blue crab (Callinectes sapidus). Results: Both 4-AP and TEA individually and in combination decreased the activity of evoked compound action potentials in isolated nerves. Conclusion: The blockers appeared to act on electrical induction and conduction within the axons, suggesting that the crab limb proprioceptive organs can be used as a model for future pharmacological investigations to increase our understanding of mechanosensitive channels and neuronal activity.
PDF Abstract XML References Citation
Received: December 20, 2021;
Accepted: March 08, 2022;
Published: April 12, 2022
How to cite this article
Hannah N. Tanner, Devan E. Atkins, Kimberly L. Bosh, Grace W. Breakfield, Sydney E. Daniels, Makayla J. Devore, Hailey E. Fite, Landys Z. Guo, Danielle K.J. Henry, Alana K. Kaffenberger, Katherine S. Manning, Tatum E. Mowery, Cecilia L. Pankau, Malina E. Serrano, Yamaan Shakhashiro, Ruth A. Ward, Aubrey H. Wehry and Robin L. Cooper, 2022. Effect of TEA and 4-AP on Primary Sensory Neurons in a Crustacean Model. Journal of Pharmacology and Toxicology, 17: 14-27.
DOI: 10.3923/jpt.2022.14.27
URL: https://scialert.net/abstract/?doi=jpt.2022.14.27
DOI: 10.3923/jpt.2022.14.27
URL: https://scialert.net/abstract/?doi=jpt.2022.14.27
INTRODUCTION
Pharmacological agents which alter neuronal function are useful for both human and veterinary clinical applications. The 4-Aminopyridine (4-AP) and tetraethylammonium (TEA), also known as Etamon1-3, work to block voltage-gated potassium (K+) channels that are involved in electrical excitability and conduction in neurons as well as certain types of tissues. The 4-AP was analyzed for use as a clinical agent in people suffering from Lambert-Eaton Myasthenic Syndrome (LEMS), but serious side effects were observed in the early trials. The 4-AP is still used in some countries and has seen an increase in use within the United States to help alleviate symptoms of Multiple Sclerosis (MS)2 and some types of poisoning3. The TEA was also assessed as a potential therapeutic agent, but these studies were not continued since other compounds were available that elicited fewer side effects. Both 4-AP and TEA may have benefits as a treatment for some forms of Parkinson’s disease4. The effects of these two pharmacological compounds on ion channels in various preparations, such as squid, amphibians and crayfish5 are well-established and have been used to demonstrate if 4-AP and/or TEA-sensitive channel types are present in a tissue of interest or to block these channels, so other channels could be examined. The differential distribution of 4-AP and TEA-sensitive channels accounts for the physiological differences in rapidly and slowly-adapting proprioceptive neurons in the muscle receptive organs (MRO) of the crayfish, which is a physiological model of the mammalian muscle spindle6.
Ion channel pathologies can cause severe neurological problems because these channels are required for normal electrical function and conduction7. Potassium channels often play a major role in the repolarization of the membrane potential back to the resting state. Typically, when open, they allow an outward flow8 of K+. However, if the channels are even slightly compromised, proprioception may be altered in the primary sensory neurons. The specific effect of these potential changes remains to be investigated.
Examining proprioceptive neurons in animal models by blocking 4-AP and/or TEA-sensitive K+ channels can illuminate the contribution of these channel types to a range of physiological functions in the whole proprioceptive organ. Chordotonal joint receptors are easily accessed in the propodite-dactylopodite (PD) chordotonal organ in the limb of a crab. The anatomical characteristics of the PD organ have been thoroughly described in earlier reports9-15. The PD organ monitors joint activity (the rate of movement and static positions of the joint) and has been used as a model to examine pharmacological agents on Stretch-Activated Channel (SAC) blockers to aid in determining the SAC receptor subtypes10-12, as well as the effect of raised potassium on electrical activity21. The direct access to the sensory endings isolated from muscle fibres facilitates the exploration of the action of diverse agents on the sensory endings as well as on axon excitability14,15.
The mechanism of action of 4-AP to help treat the disease state of MS in mammals is difficult to study since myelinated neurons become unmyelinated and ion channels change in their distribution and expression16. Understanding how unmyelinated neurons respond to therapeutic compounds in other model organisms may provide insight into their potential therapeutic efficacy. This study contributes to our understanding of the actions of 4-AP and TEA on crab proprioceptive neuron function10-15.
The objective of this study was to examine the effects of 4-AP and TEA on the sensory nerves of a crab model for neurophysiological studies. Potentially this model organism can be used for future studies in neurophysiological investigations building on these initial findings.
MATERIALS AND METHODS
Study area: The procedures used in this study are similar to those recently published12-15, but they are described briefly below. These studies were conducted from May to December, 2021 within the Department of Biology at the University of Kentucky in Lexington, Kentucky, USA.
Animals: Blue crabs (Callinectes sapidus) were obtained from a distribution centre in Atlanta, GA, USA and delivered to a local supermarket in Lexington, KY, USA. Crabs were maintained in a seawater aquarium before use for three to five days. All experiments were performed on adults with a carapace width (from point to point) of 10-15 cm. The crabs were fed frozen squid and the water temperature was maintained between 14-16°C. The crabs were caught from the wild and were two to three years old. Forty-five crabs were used in this study.
Physiological measures: Procedures for dissection and preparation of the crab PD organ can be found in video format17. The first or second legs were obtained from live animals using “autotomization,” which takes advantage of a natural process used by the animal as a response to a predator or when captured in a net (Fig. 1a). There is a natural fracture plane and the animals routinely regenerate the legs (similar to a lizard’s tail being lost and regenerating),there is very little bleeding associated with the process.
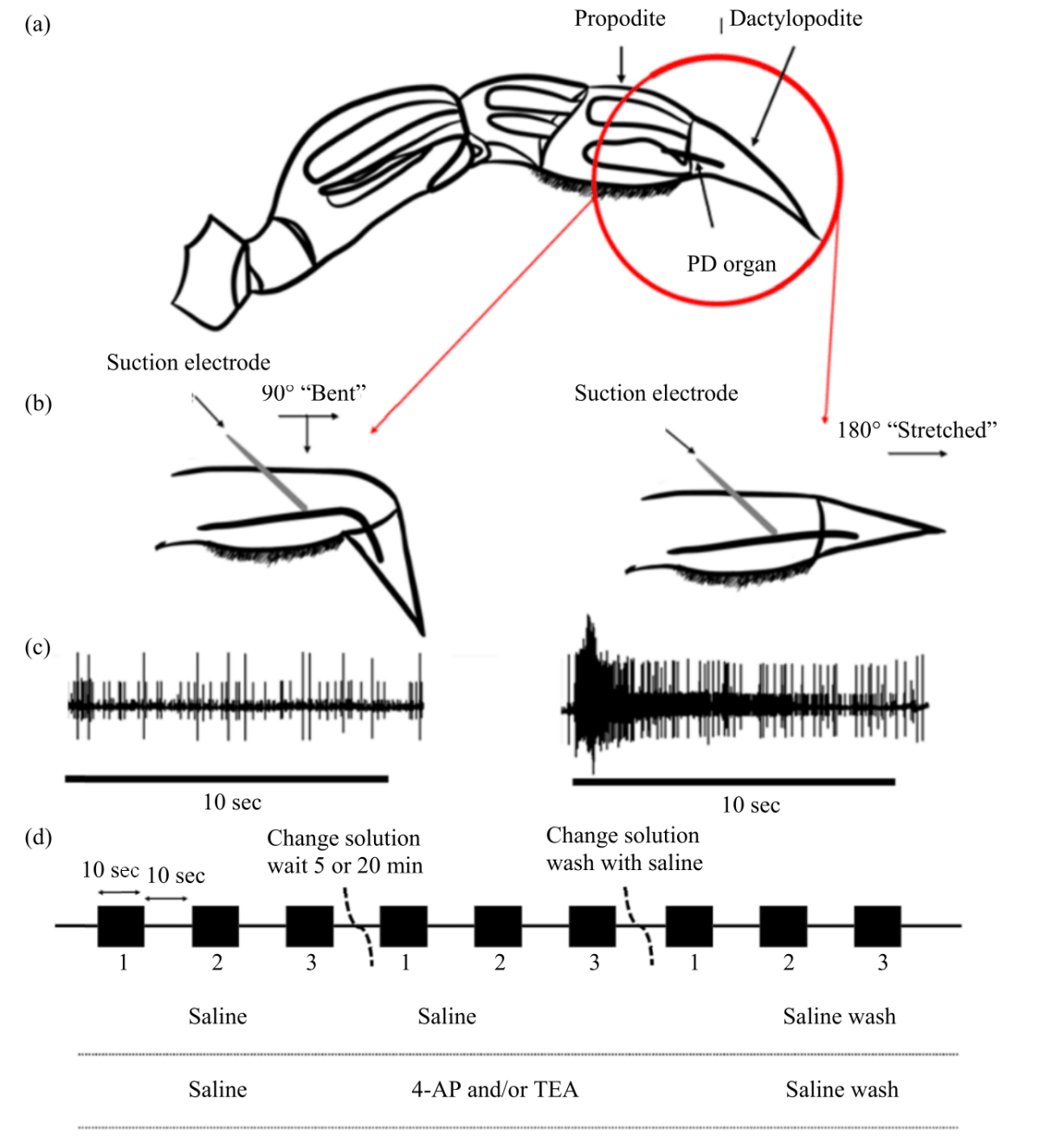 |
| Fig. 1(a-d): | Experimental paradigm for moving the joint in a crab limb while recording from the proprioceptive nerve, (a) An outline of a walking leg of the crab highlighting the apodemes (i.e., tendons) in each segment and the PD chordotonal organ in the most distal joint, (b) Illustration of the movements of the PD joint, (c) Neural activity in the bent and extended positions. The number of spikes within the first 10 sec (the 1 sec movement to the extended position followed by 9 sec in a static extended position) was used to quantify the amount of activity, (d) Paradigm for obtaining the average number of spikes for each preparation (saline bath, saline+compounds, saline wash). The displacement was repeated three times per saline bath and for each treatment |
Autotomization can be induced by lightly pinching at the base of the leg distal to the autotomy plane. On the day of experimentation, the two first walking legs were induced to be autotomized and the crabs were then euthanized by placing them in a -20°C freezer.
The autotomized leg was placed in a Sylgard-lined dish with crab saline. After the PD nerve was isolated the proximal end of the nerve was pulled into a suction electrode for recording. In the experimental paradigm to activate the proprioceptive neurons, the dactyl was moved from a flexed position to an extended position. The movement was conducted within a 1 sec time frame and held in the extended position for 10 sec before the joint was moved back to the starting position (Fig. 1b, c). An insect dissecting pin was used as a stop limit for each displacement to maintain consistency and each displacement period was marked on the computer recording file (Fig. 1c). To ensure accurate activity levels, the joint was extended 20 times without a hold before measurements were taken. Responses were recorded from the PD organ in saline, followed by saline containing 4-AP and/or TEA. After exposure to the compounds, the bath was exchanged with fresh saline without the pharmacological agents. Neural activity was assessed to compare the initial response to the compounds and then after their removal by exchanging them to fresh saline. For control preparations, saline was exchanged after the initial recordings for fresh saline without 4-AP or TEA. Three trials were performed for each condition (Fig. 1d) and the activity from the set of three trials was averaged.
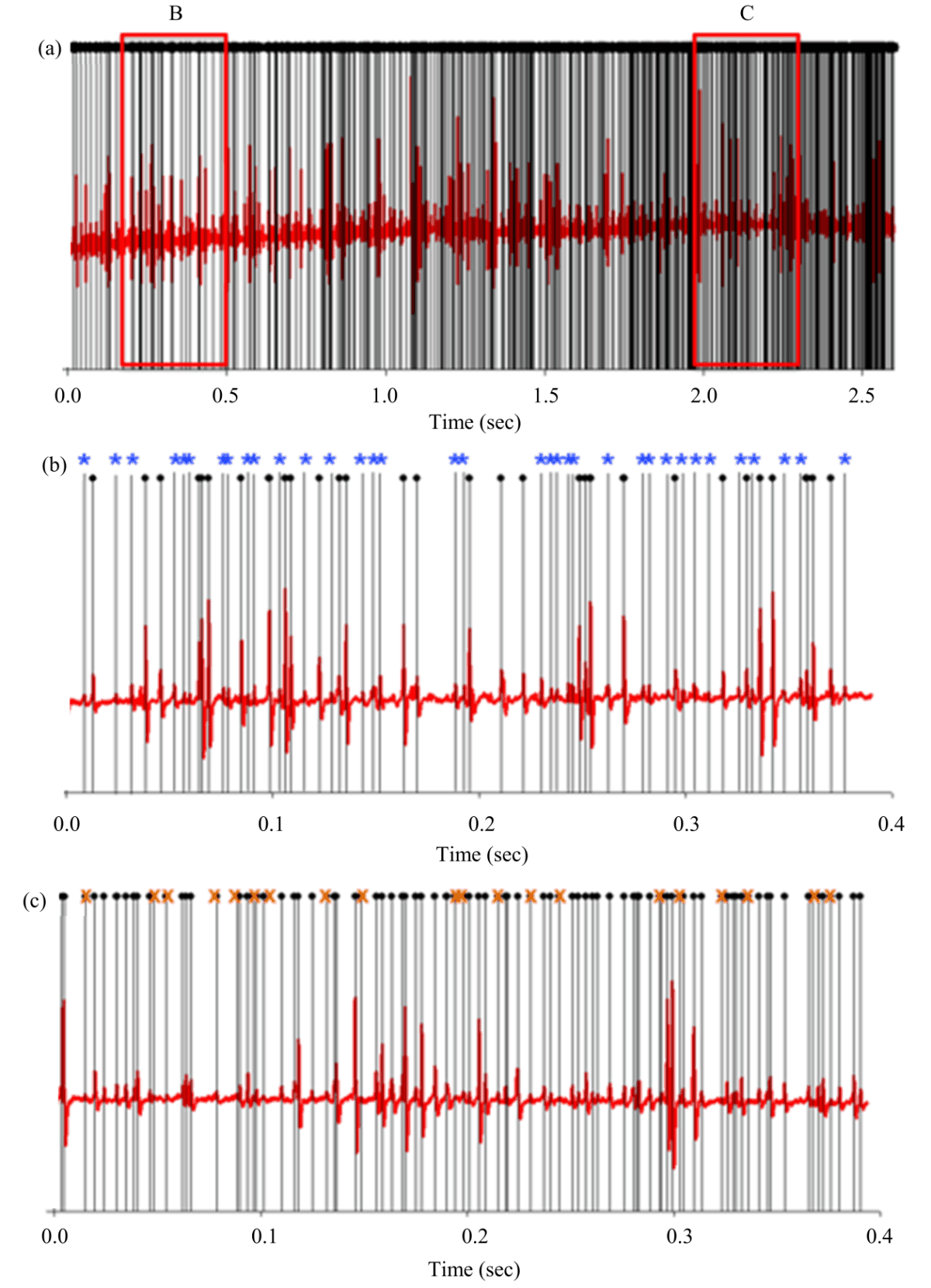 |
| Fig. 2(a-c): | Software was used to determine the number of spikes in each 10 sec during the joint movement and hold displacement, (a) Representative trace as well as where the program counted the spikes, (b) Portion of that trace where spikes were added (shown by a star) and (c) Portion of that trace where spikes were removed (shown by an x). A Standard Deviation (SD) between 0.05 and 9.00 was chosen and the number of spikes that fell above the SD of an averaged baseline was determined |
The saline used consisted of (in mM) 470 NaCl, 7.9 KCl, 15.0 CaCl2⋅2H2O, 6.98 MgCl2⋅6H2O, 11.0 dextrose, 5 HEPES acid and 5 HEPES base adjusted to pH 7.5. The 4-AP (100 mM) and TEA (50 mM) were used as reported in previous studies utilizing crayfish sensory nerves10. Preparation was incubated with TEA for 20 min before assessing any neural activity changes. The pH of the saline was adjusted after adding the compounds to maintain the pH at 7.5. All chemicals were obtained from Sigma-Aldrich, St. Louis, MO, USA.
Suction electrodes made from glass pipettes fitted with plastic tips were used to record extracellular signals from the cut nerves (details of making the suction electrodes is provided in18). A P-15 amplifier (Grass Instruments, Astro-Med West Warwick, Rhode Island, USA) in conjunction with a PowerLab/4s A/D converter and Lab Chart 7 software (ADI Instruments, Colorado Springs, CO, USA) was used to obtain the signals, which were recorded on a computer at a 10 or 20 kHz sampling rate.
Neural activity was determined by the number of spikes (i.e., extracellularly recorded action potentials) which occurred within a 10 sec period from the start of the joint movement (Fig. 1). The averaged values in the three trials for each bath condition are presented. The software analysis program can detect any electrical signal above a set standard deviation from the mean (Fig. 2a).
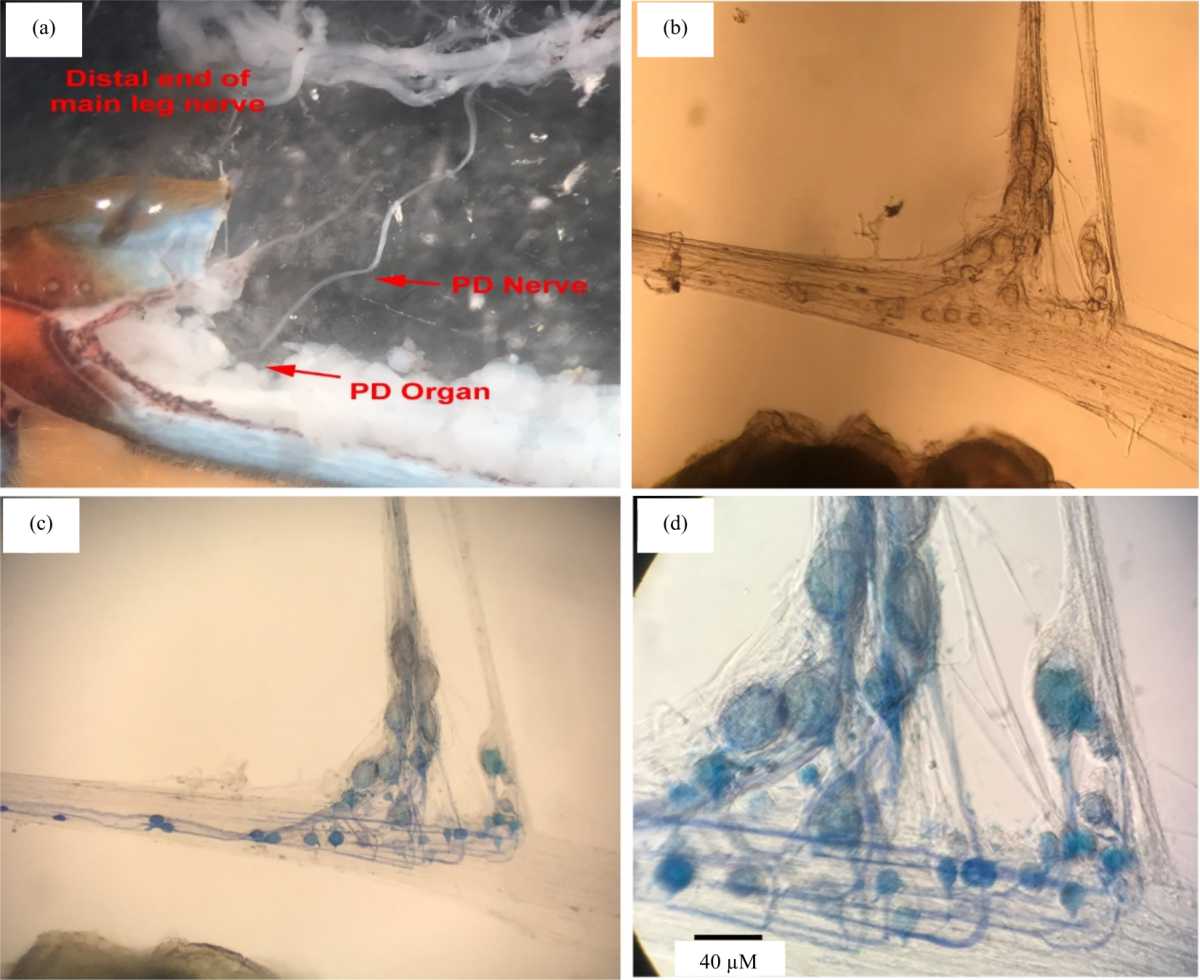 |
| Fig. 3(a-d): | PD nerve and PD organ, (a) PD nerve is left intact to the PD organ to record the activity from displacing the joint with an en passant recording of the nerve. The proximal end of the main leg nerve can be used to evoke compound action potentials in the PD nerve while recording the neural activity from the PD organ. However, the PD nerve can also be transected from the PD organ and isolated to induce and record compound action potentials (Modified from Atkins et al.19), (b) PD organ illustrates the unstained neuron cell bodies associated with the elastic strand, (c) Same preparation is shown in (b) but lightly stained with methylene blue which is selectively taken up in the neurons20 and (d) Enlarged view is shown in (c). The connective sheath over the cell bodies is observed. Sensory endings are embedded within the elastic strand |
However, the automated analysis was not always consistent with the actual number of events observed. Therefore, each 10 sec period in the traces was reviewed by eye and additional spikes were added (Fig. 2b) or removed (Fig. 2c). The final number of spikes was then used for statistical analysis.
To record compound action potentials (CAPs), the PD nerve to the PD organ was left intact during the dissection and was peeled approximately 1 cm away from the main leg nerve (Fig. 3). To illustrate how the detailed analysis was performed a typical trace in the spikes from a PD nerve are shown (Fig. 3a). Spikes that were not detected by the standard software analysis were added by direct analysis by an observer looking over the data file (Fig. 3b). Additionally, noise in the trace as determined by an observer was removed from the counts (Fig. 3c). The distal end of the nerve was cut close to the PD organ and pulled into a suction electrode, which was then sealed with petroleum jelly. The proximal end of the main leg nerve was dissected out of the leg and pulled into a suction electrode and stimulated at varying voltage to induce a maximum amplitude and shape of the CAP measured at the distal end. Leaving the main leg nerve associated with the PD nerve reduced the time needed for dissection and allowed the PD nerve to be fully exposed to the compounds while changing out the bathing media The isolated PD nerve from the PD organ was used for recording the refractory periods of the PD nerve.
Statistical analysis: All data are expressed as a mean (±SEM). A repeated-measures ANOVA and repeated measures T-test, as well as a Wilcoxon Signed Rank Test, was used to compare the differences in responses before and after exchanging solutions. The normality test was Brown-Forsythe or a Shapiro-Wilk depending on whether the initial analysis used an ANOVA or a T-Test (respectively). The analysis was performed with Sigma Stat software. A p-value <0.05 was considered statistically significant for determining changes in neural activity after exposure to the compounds. To examine the consistency and reproducibility of the data, groups of participants blinded to the specific settings of the analysis software were asked to supply their interpretations of the number of spikes for some of the same data sets.
RESULTS
Effect of TEA and 4-AP on activity with joint movements: To confirm that the alterations in the PD nerve activity resulted from changing the bathing media and maintaining the recording for the duration of the experimental paradigm, control experiments were conducted by exchanging the saline media with fresh saline (Fig. 4). The bath was exchanged twice to mimic the conditions used for the pharmacological agents. Representative traces are shown (Fig. 4). The initial trace of the nerve activity in saline (Fig. 4a), after exchanging the bath with fresh saline (Fig. 4b) and after rinsing the bath out again with fresh saline (Fig. 4c) to control for the effects of exchanging the bathing environment for the pharmacological studies. This paradigm was used as a control for exchanging the bathing media and movements of the joint over time.
When 100 mM 4-AP was included in the second saline bath, PD nerve activity was significantly altered, as shown in the representative traces (Fig. 5). The initial trace of the nerve activity in saline (Fig. 5a), during exposure to 4-AP (Fig. 5b) and after rinsing the bath out with fresh saline not containing 4-AP (Fig. 5c).
The delayed potassium blocker TEA also demonstrated an effect on PD nerve activity (Fig. 6). The initial trace of the nerve activity in saline (Fig. 6a), during exposure to TEA (Fig. 6b) and after rinsing the bath out with fresh saline not containing TEA (Fig. 6c). Exposure to either 4-AP or TEA resulted in changes in neural activity. When bathed in saline plus both compounds, six out of six preparations displayed a bursting in neural activity. Representative initial saline exposure traces and exposure of 4-AP plus TEA is shown in Fig. 7. The initial trace of the nerve activity in saline (Fig. 7a), during exposure to 4-AP combined with TEA (Fig. 7b) and after rinsing the bath out with fresh saline not containing the pharmacological agents (Fig. 7c).
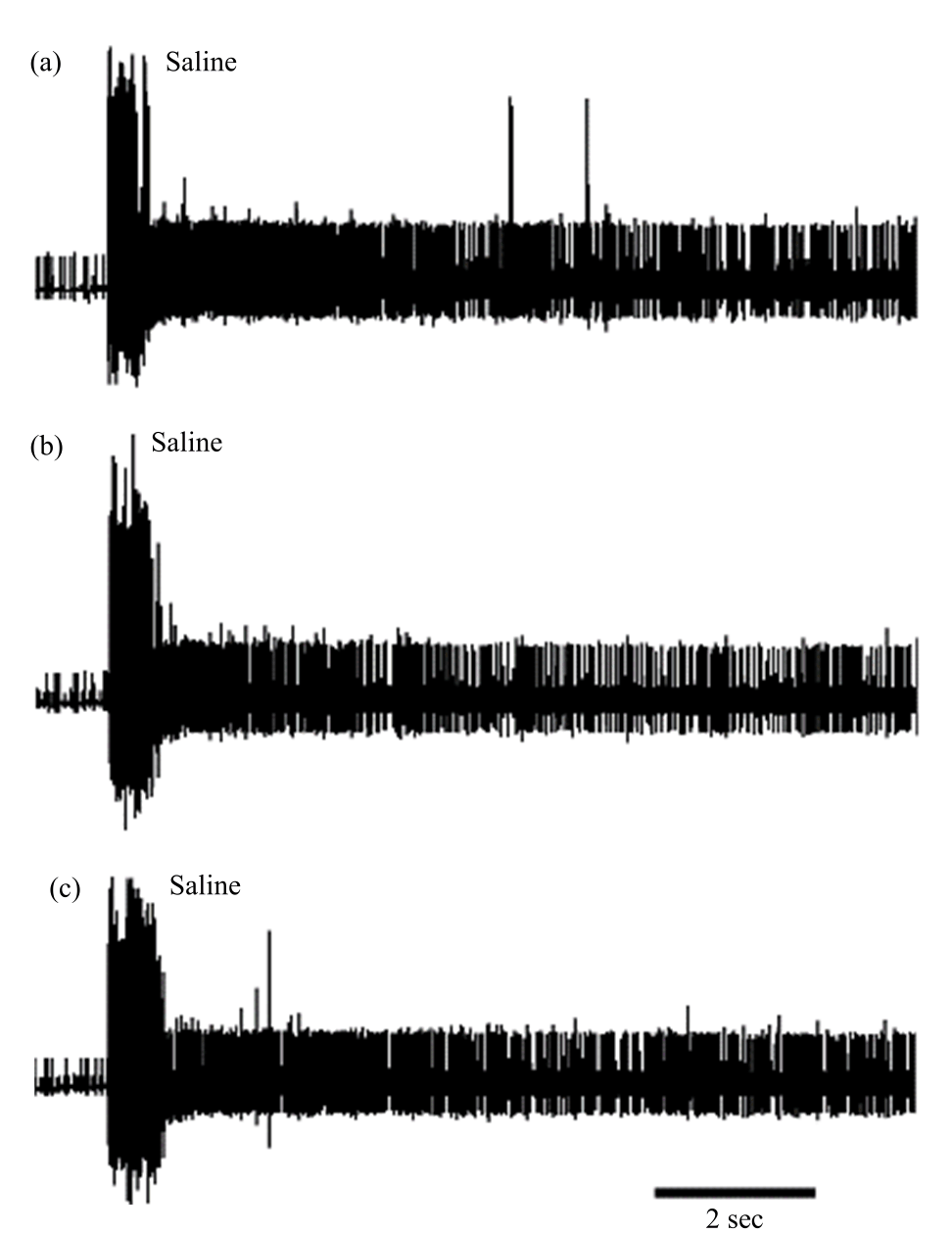 |
| Fig. 4(a-c): | Representation of the neural activity after exposure to the initial saline bath followed by two exchanges of the bathing media with fresh saline, (a) Initial saline bath, (b) After the first exchange in this case with saline only again and (c) Second exchanged with saline only |
Following the combined drug exposure and rinsing with fresh saline, the nerve did not regain a normal firing pattern. This bursting behaviour appears to be associated with exposure to 4-AP rather than to TEA, as some bursting patterns appeared in trials with only 4-AP.
For comparisons among drug treatments and variability among preparations the neural activity, as assessed by the number of spikes within the 10 sec from the start of the movement to a static position within 1 sec and held for the remaining 9 sec in the fully extended position, is shown for each of the six preparations under each experimental condition (Fig. 8a). Each line is an individual preparation and each symbol represents the average number of spikes for the three trials in each period of the experimental paradigm (Fig. 1). There was no significant effect in overall activity when exchanging initial saline for the second saline. However, from the initial saline to the final wash saline there was a significant decrease in overall activity. Treatment with 4-AP significantly decreased activity from initial saline to the 4-AP and from the initial saline exposure to the saline rinse after 4-AP exposure, (Fig. 8b).
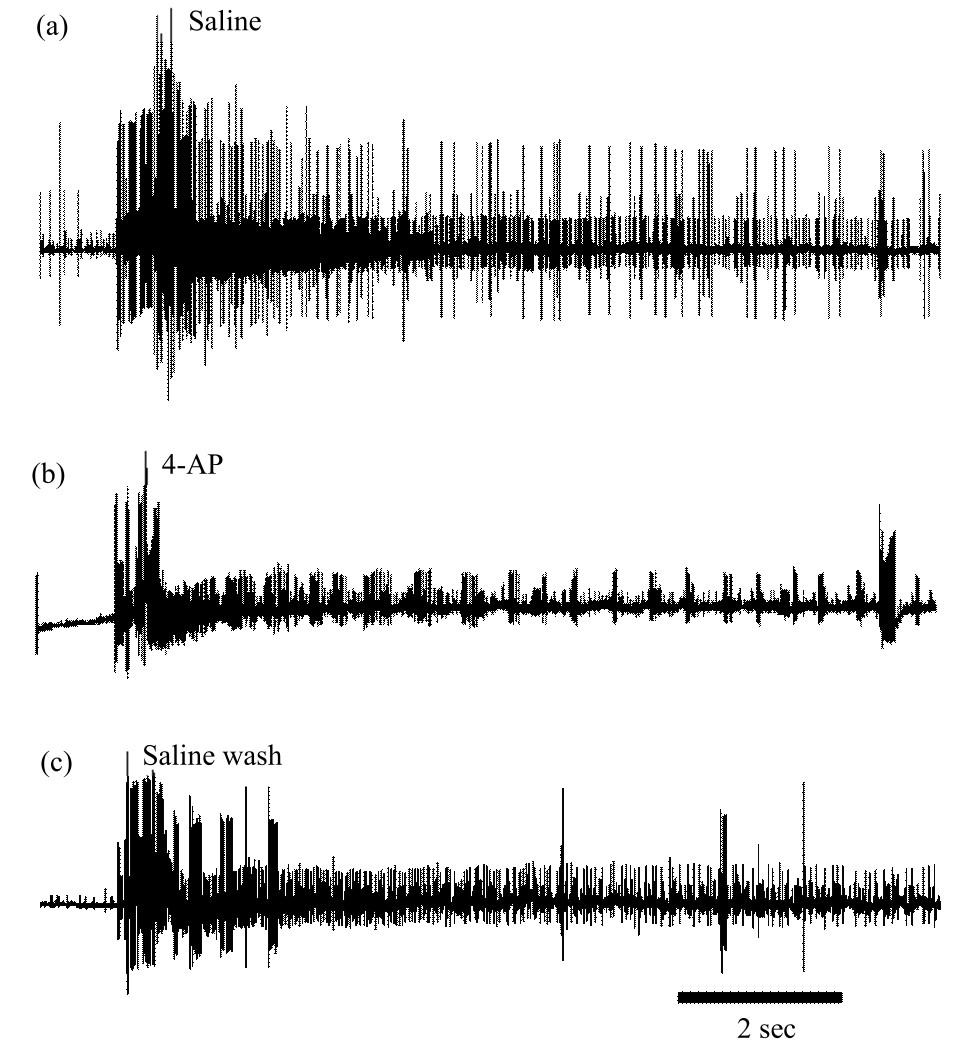 |
| Fig. 5(a-c): | Effect of 4-AP on neural activity during activation of the proprioceptive neurons, (a) Representation of the neural activity after exposure to the initial saline bath, (b) Followed by an exchange of the bathing media to saline+100 mM 4-AP and (c) A washout of the 4-AP with fresh saline. The neural activity of the PD nerve was dampened (b) when compared to the initial activity in saline (a). Exchanging the bath back to fresh saline increased but did not fully recover the activity (c) |
Treatment with TEA significantly decreased activity from initial saline to the TEA and from the initial saline exposure to the saline rinse after TEA exposure (Fig. 8c). The two drugs in combination also resulted in a significant overall decrease in neural activity. However, the data set for the paradigm for both 4-AP and TEA was not normally distributed. Thus, a Wilcoxon Signed Rank Test was used to compare the differences in responses before and after exchanging solution from the initial saline to the combined 4-AP and TEA exposure and again from initial saline to the washout saline. Both of these comparisons were significant (Fig. 8d).
The percent change in the number of spikes from the initial saline exposure was determined for each preparation under each experimental condition (Fig. 9). The preparations with 4-AP displayed a decrease of 66.91% in the number of spikes, the preparations with TEA displayed a decrease of 57.95% and the preparations with the combined TEA and 4-AP had a decrease of 80.48% when compared to the initial activity.
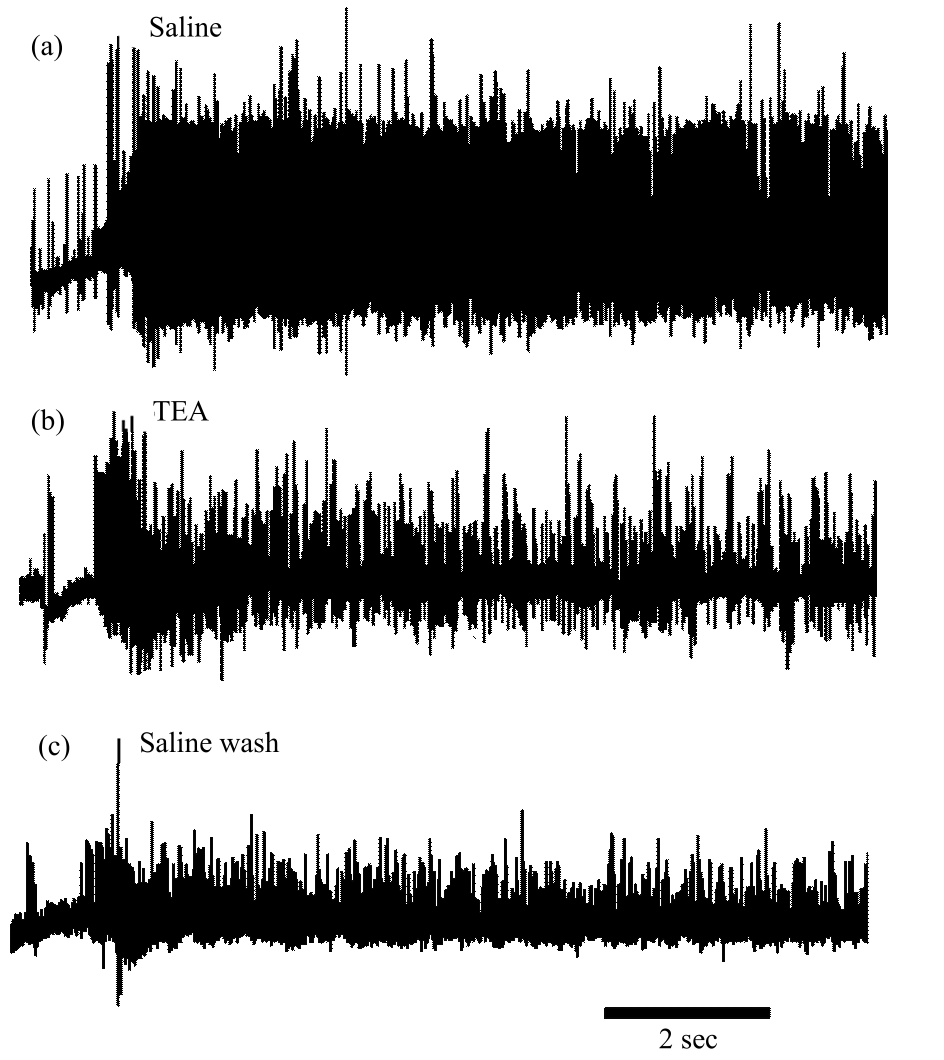 |
| Fig. 6(a-c): | Effect of TEA on neural activity during activation of the proprioceptive neurons, (a) Representation of the neural activity after exposure to the initial saline bath, (b) followed by an exchange of the bathing media to saline+50 mM TEA and (c) A washout of TEA with fresh saline. The neural activity after exposure to TEA is dampened (b) when compared to the initial activity in saline (a). Exchanging the bath back to fresh saline increased the activity but had lower amplitude spikes (c) |
Compound action potentials (CAPs): To understand how these compounds alter electrical conduction along the nerve, the preparation was exposed to TEA or 4-AP while the joint was held in a static position and evoked Compound Action Potentials (CAPs) were induced (Fig. 10). The effects on the evoked CAP and spontaneous activity with exposure to TEA (Fig. 10a) or 4-AP (Fig. 10b) is illustrated. Both 4-AP and TEA induced multiple firings of the nerve. This suggests that multiple neurons did not repolarize below the threshold but remained depolarized after the initial stimulus. This would also imply that the voltage-gated sodium channels did not remain in an inactive state after the initial depolarization. Likely, the membrane potentials were sufficiently repolarized to reactivate at least some sodium channels so that at the threshold potential these channels were able to fire again to produce these complex CAP waveforms.
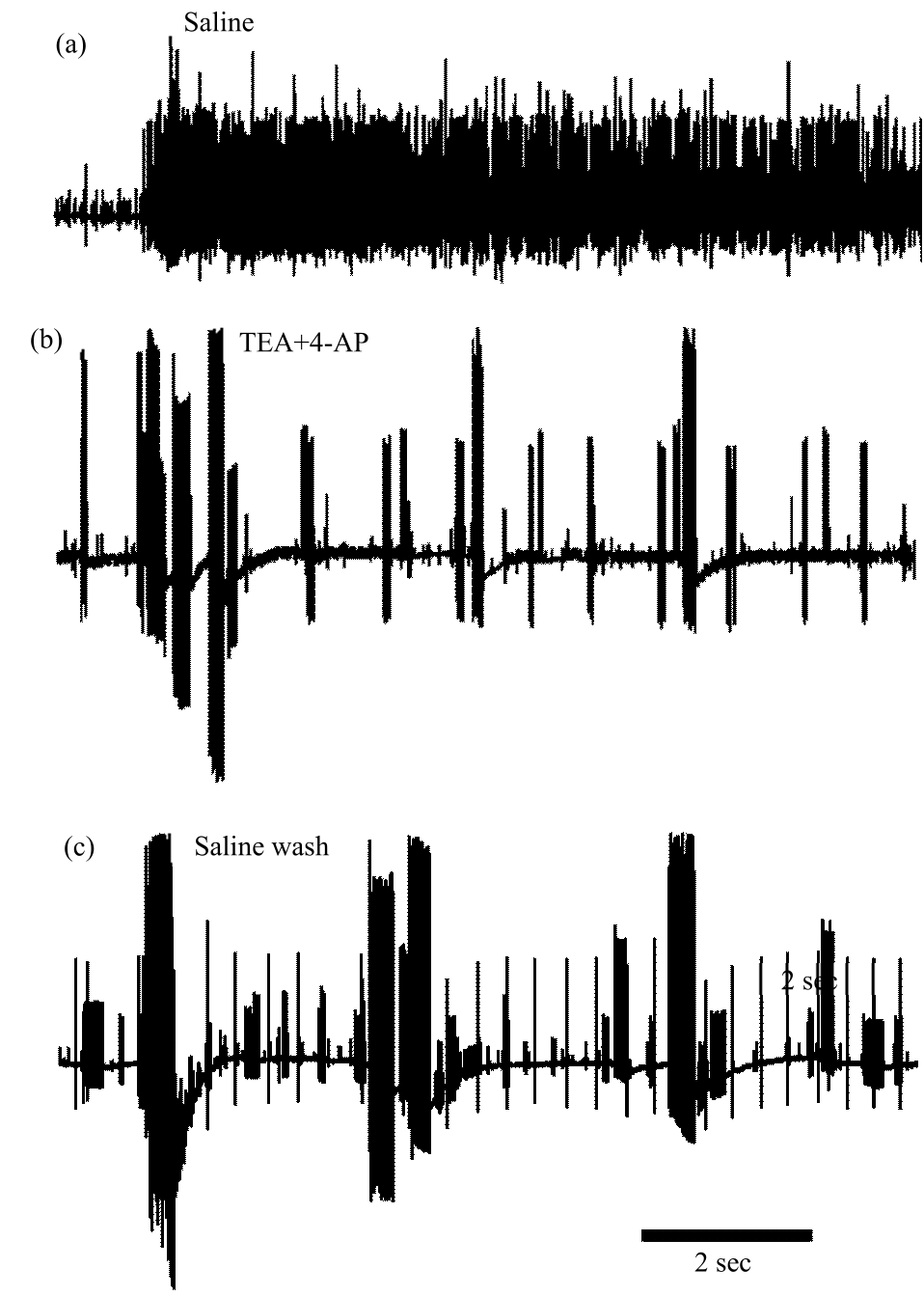 |
| Fig. 7(a-c): | Effect of combining 4-AP and TEA on neural activity during activation of the proprioceptive neurons, (a) Representation of the neural activity after exposure to the initial saline bath, (b) Followed by an exchange of the bathing media to a combination of 100 mM 4-AP and 50 mM TEA and (c) A washout of the compounds with fresh saline (c). The neural activity was dampened when compared with the initial activity in saline (a). Exchanging the bath back to fresh saline increased the activity (c). Bursting behaviour of the activity with exposure to the compounds |
Refractory period of the PD nerve: The absolute refractory period was difficult to obtain in most preparations due to the complex and prolonged duration of the CAPs (Fig. 10). However, when a relatively simple CAP waveform was obtained with a set stimulation voltage, the absolute refractory period was possible to determine (Fig. 11) with varying the delay between the two evoked stimuli as shown in Fig. 11a-c, as the delay for the 2nd stimulus becomes shorter and shorter.
However, with either 4-AP or TEA exposure, the nerve repetitively produced action potentials and CAPs which were not associated with the evoked stimulus (Fig. 12), even with varying the delay between the two evoked stimuli as shown in Fig. 12a-c, the nerve continued to produce 3 CAPS not related to the delay for the 2nd stimulus.
DISCUSSION
Both 4-AP and TEA suppressed the ability of the nerve to conduct electrical signals, although when in combination, they did not have a greater effect on neural activity than the individual effects of the compounds alone. Removal of the compounds and exposure to fresh saline generally increased activity. Given that the stimulation of the nerve was accomplished by activating the stretch-activated channels (SACs) within the sensory endings embedded within the PD chordotonal organ, the pharmacological agents could have had direct effects on the SACs themselves. This seems unlikely since previous studies have reported that there are no effects on PD activity with low pH (pH = 5) or applications of ruthenium red or amiloride on the PD organ using the same displacements of the joint as used in the current study. It appears the SAC subtype in crustacean chordotonal organs is not PIEZO1 nor a DEG/ENaCs (Degenerin/epithelial sodium)-dependent channel20. Only Gd3+ application reduced activity, which was reversible when washed out. The SACs might be a type of Ca2+ ion channel that will also conduct Na+, since a bath without Ca2+ ions still produced neural activity with PD joint movement, However, these SACs are blocked19-20 by Mn2+.
The CAPs evoked in the isolated PD nerve changed shape with exposure to 4-AP and TEA. The axons of the neurons were too narrow to obtain intracellular recordings of discrete action potentials. The cell bodies also appeared to have a connective sheath covering them which broke the fine microelectrode glass tips. Attempts to obtain intracellular recordings from the somas are ongoing.
We have not found any reports on the action of 4-AP and TEA on sensory neurons in Blue crabs. These compounds have been used previously in a freshwater crustacean species (crayfish) and it has been shown that TEA broadened the action potential shape in a motor neuron innervating the opener muscle in the walking leg in this species11. It is likely to have the same effect on the crab PD neurons. The broadening of the action potential would prevent the neuron from repolarizing as rapidly and prevent removal of the inactivation of the voltage-gated Na+ channels.
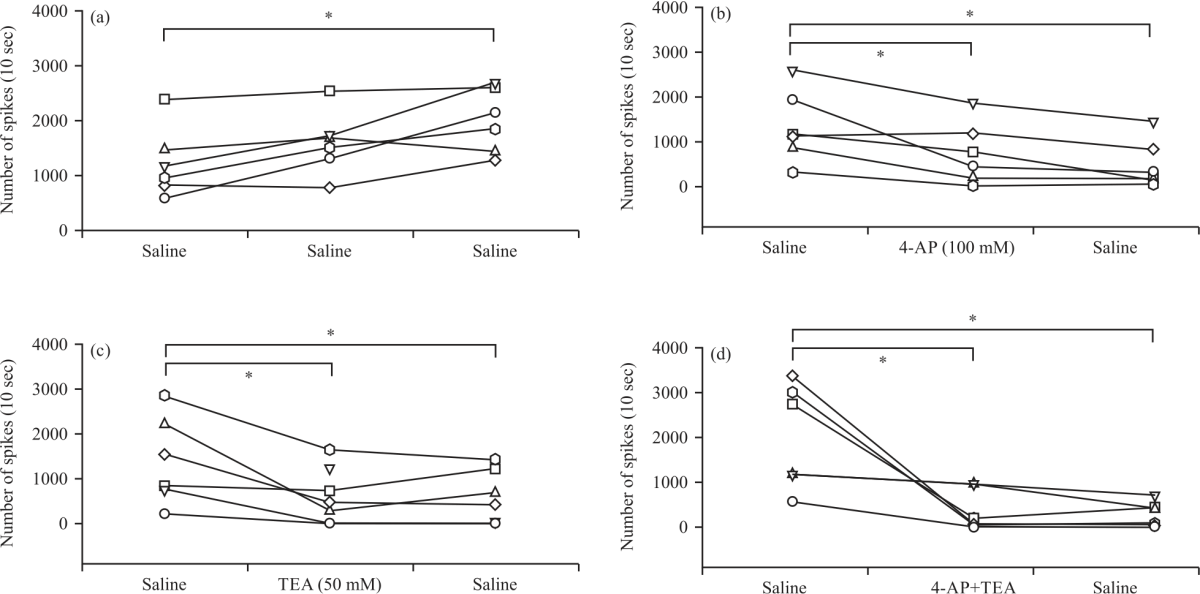 |
| Fig. 8(a-d): | Activity of the PD nerve with a displacement of the PD joint from the flexed position to an extended position. The mean number of spikes (average of three displacements in each bathing media) for 10 sec with the 1st sec moving the joint from a flexed position to an extended position for the remaining 9 sec (static position). Six preparations were used for each condition. (a) Saline only, (b) 100 mM 4-AP, (c) 50 mM TEA and (d) 100 mM 4-AP+50 mM TEA The different symbols represent individual preparations that were used only once for a given condition. The thicker lines above each graph represent the statistical comparisons for the different groups. The number of spikes was statistically compared from the initial saline to the subsequent change in the bathing environment as a repeated-measures ANOVA. However, the distributions were not normally distributed for the data set for the paradigm in which both 4-AP and TEA were combined. A Wilcoxon Signed Rank Test revealed a difference in responses before and after exchanging solution from the initial saline to the combined 4-AP and TEA exposure and again from initial saline to the washout saline and *represents a p<0.05 |
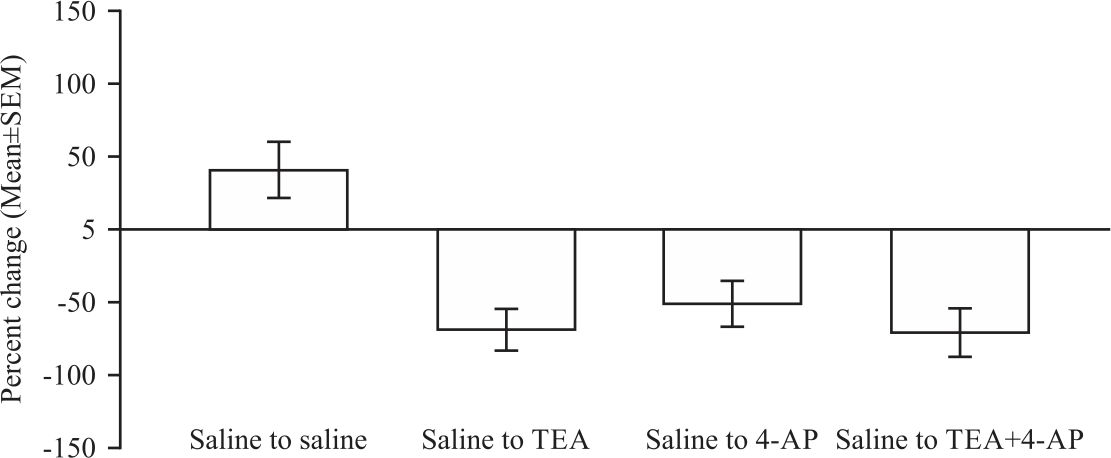 |
| Fig. 9: | Mean percent change from the initial saline exposure to the following bath exchange |
Thus, the neurons may have a longer period of absolute and relative refractory periods with a broad action potential. If this occurs, then the overall activity of the PD nerve would be expected to have less activity upon displacements of the joint. However, if the axons had a high density of calcium-activated potassium channels (K(Ca)) and the membrane was depolarized longer due to the broad action potential from blocking the voltage-gated K+ channels, then one would also expect the voltage-gated Ca2+ channels to remain open longer, promoting the K(Ca). This would result in the membrane remaining hyperpolarized for a longer period and a decrease in the activity of individual neurons.
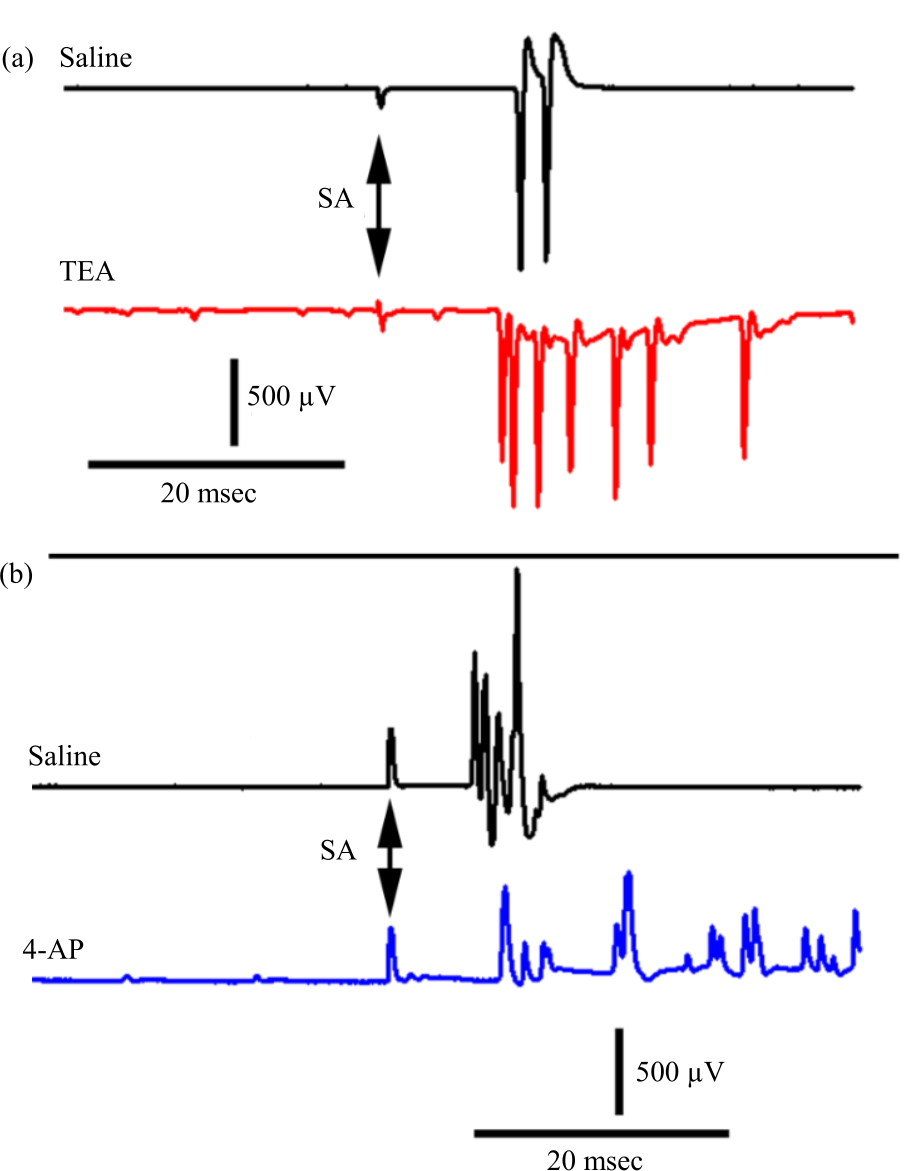 |
| Fig. 10(a-b): | Representative evoked compound action potentials in the PD nerve before and after exposure to, (a) TEA (50 mM) and (b) 4-AP (100 mM) Note the compound action potentials are mixed, meaning that the neuronal population assayed has differing conduction properties. Thus, the compound action potential spread out in duration compared to exposure to saline alone. Upon exposure to either 4-AP or TEA, the evoked compound action potentials dampened in amplitude and the nerve repetitively produced activity with the same given stimulus as in the saline alone exposure. Note the small compound action potentials in the traces after 4-AP or TEA was added. The stimulus artifact is depicted as SA |
It should be noted that the unmyelinated neurons of the crab sensory nerve may not respond in the same manner to treatments of 4-AP and TEA as myelinated amphibian and mammalian neurons due to the localized regions of the subtypes of voltage-gated potassium channels in the paranodal (4-AP sensitive) and nodal (TEA sensitive) regions7. Since the delayed potassium channel (TEA sensitive) is slow in the opening, it likely does not contribute as much to the repolarization of mammalian myelinated neurons as the rapidly-acting 4-AP-sensitive potassium channels. However, the TEA sensitive channels may account for the basal potential with repetitive firing and alter the responses during the high frequency of neural activity.
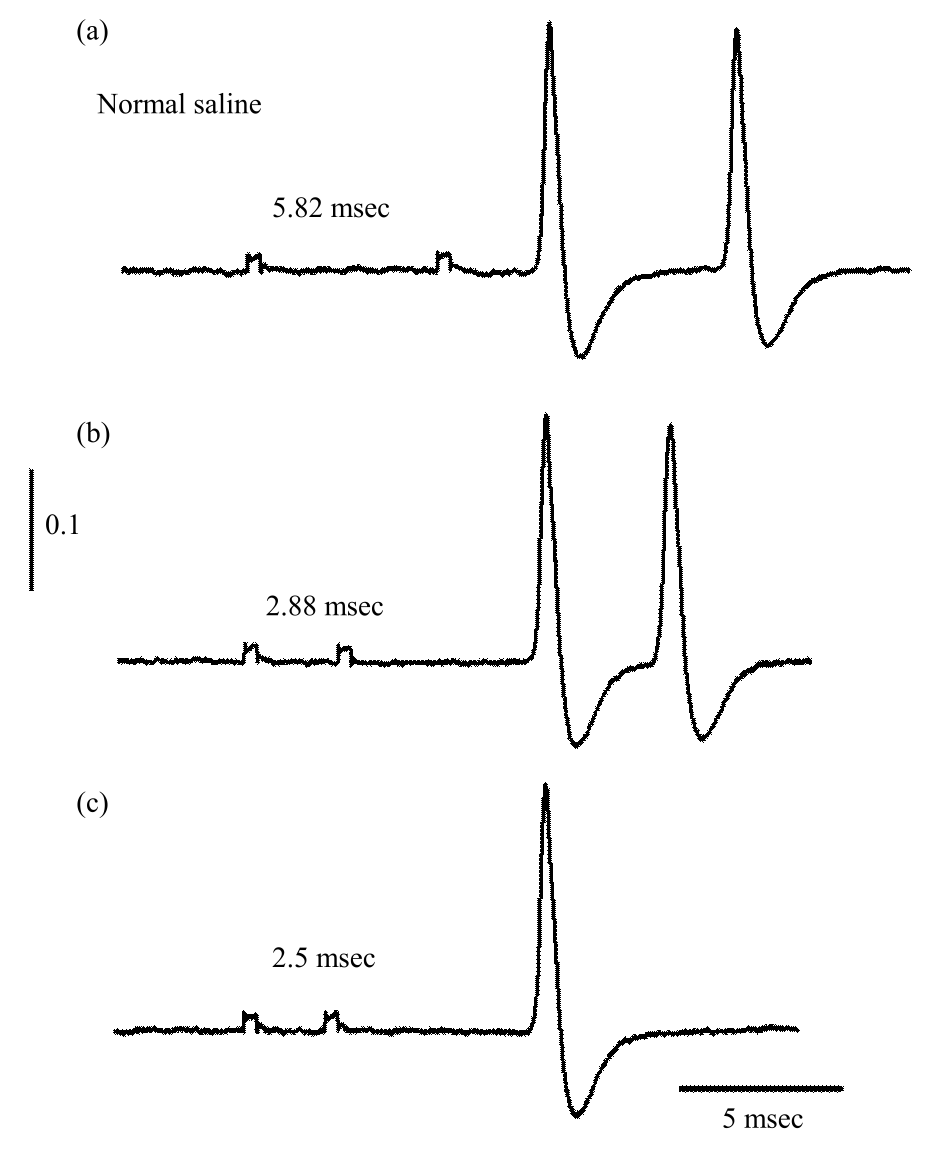 |
| Fig. 11(a-c): | Representative illustration of the absolute refractory period in the PD nerve when the evoked compound action potential (CAP) was simple in shape, (a) Two CAPS produced when the two evoked stimuli are 5.82 msec apart as compared to, (b) when the delay is 2.88 msec and (c) Delay with the second evoked stimulus is so short that the absolute refractory period is produced with a delay of 2.5 msec |
The action of TEA may also depend on the level of activity due to the opening and closing of ion channels as this compound may block when channels are in an open state. It has been shown that the clinical use of 4-AP in humans can induce seizures as a side effect which would suggest that some neurons are hyperexcitable. This could be due to a reduction in inhibitory control.
The mechanism of action for 4-AP-enhanced therapeutic outcomes for improving motor skills in some individuals with multiple sclerosis is unclear16,21. The complexity of myelination loss in the CNS, infiltration of microglia and inflammatory responses (i.e., activated T cells and macrophages) may be a direct result of 4-AP action, resulting in secondary responses on neurons22. However, understanding how 4-AP and TEA work individually and in combination in different neuronal models will address the action on unmyelinated nerves and the physiological activation of a sensory system.
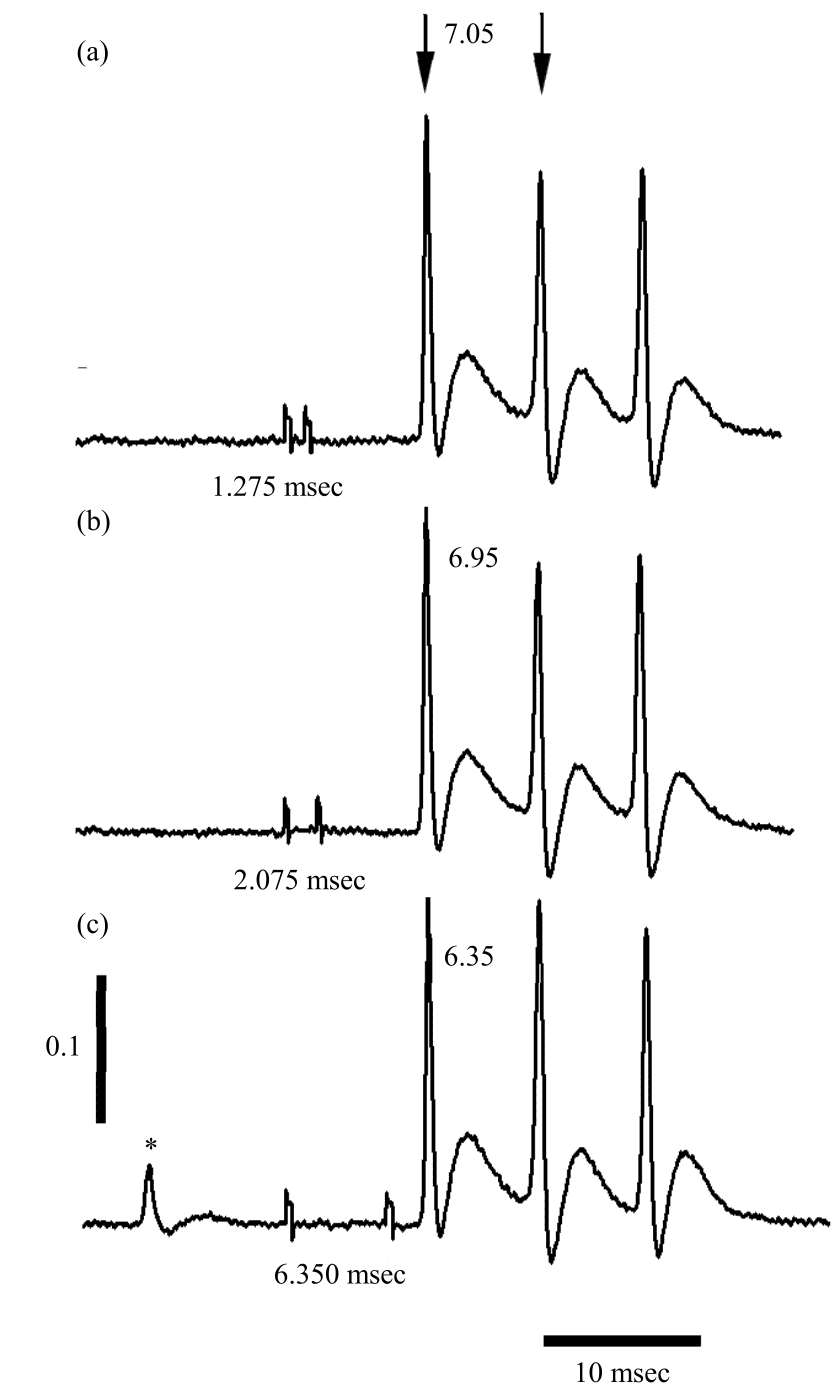 |
| Fig. 12(a-c): | Representative trace illustrating the difficulty in obtaining the absolute refractory period in the PD nerve. When the nerve was exposed to either 4-AP or TEA, the nerve became hyperexcitable and produced a volley of activity despite altering the duration of the second evoked stimulus. (a) Two evoked stimuli are 1.275 msec apart as compared to, (b) When the delay was 2.075 mec and the same three compound action potentials occur and (c) Delay with the second evoked stimulus was so long, 6.350 msec, that evoked compound action potentials should have spread out, but instead, the responses appear the same as with a very short delay between stimuli Note in (c) the * indicates a random compound action potential which occurs when evoking the nerve with exposure to 4-AP or TEA or with the two compounds combined |
CONCLUSION
This study presented a crustacean model of a proprioceptive organ and nerve which can be used to screen pharmacological compounds targeting sodium and potassium voltage-gated channels to determine the effects on electrical conduction and neural activity. Two commonly used potassium channel blockers (i.e., 4-AP and TEA) depressed sensory nerves to conduct electrical signals. However, if the nerve is electrically stimulated, random excitation of the nerve can occur. In the experimental paradigms described in this study, the combined application of TEA and 4-AP did not have a greater effect on neural activity than when the compounds were used in isolation at the same concentration. After removal of the compounds, the neural activity was not fully recovered, suggesting that either the compounds could not be completely removed by exchanging the saline bath and/or that there were long term effects on neural function.
SIGNIFICANCE STATEMENT
This study provided the usefulness of the crab model to address neurophysiological responses to the pharmacological compounds 4-AP and TEA. This illustrates that the neurons in the crab have ionic channels which are also sensitive to these blockers of voltage-gated potassium channels as for other model organisms. In addition, the hyperactivity of the evoked CAPs illustrates how application of these compounds in mammalian preparations likely also produces increased activity and may account for the seizure-like activity which can be induced by the application of these compounds therapeutically. In addition, an approach to provide a modification to correct oversights on standard software analysis is detailed.
ACKNOWLEDGMENTS
We thank Wendi Neckameyer, PhD for editorial assistance. Appreciation is given to the following participants in a previous course in which the concentrations of 4-AP and TEA to be used were determined for this present study and noting the preliminary results: Saadia Akhtar, Ross Basham, Bharath Chithrala, Bennett Collis, Bernardo Aguzzoli Heberle, Emma Higgins, Allison Lane, Saisindhu Marella, Matthew Ponder, Prachi Raichur, Aaron Silverstein, Alexandra E. Stanback, Maddie Stanback, Catherine Stanley and Kelsi Vela.
SUPPLEMENTARY MATERIALS
Reproducibility in the analysis of sensory activity: It is important to address the reproducibility in data analysis for the number of spikes, since the automated software for detecting the spikes of an average baseline value may produce some errors which are easily detected by visual observation (Fig. 2). In this study, spurious events were subtracted and missed events by the automated analysis were added. The first pass in the data analysis was with setting a threshold in the standard deviation of the mean values of the trace.
To assess if the visual adjustments were consistent, random data sets were provided to college students. Eight groups, consisting of two students per group, were given a data set labelled A through D. A movie was provided to course participants on how to use the automated software to analyze events in a recording and how to add or subtract events as deemed necessary. Since this was a blind study for the participants, the code was not provided until after the analysis was completed. The treatments were saline alone (Fig. S1, groups A1 and A2), 4-AP (Fig. S1, groups B1 and B2), TEA (Fig. S1, groups C1 and C2) as well as a combined solution of 4-AP and TEA (Fig. S1, groups D1 and D2). The general trends for each of the three trials in saline and the unknown are shown in Fig. S1a. The average response for the three trials in each condition is shown in Fig. S1b for each treatment by the course participants.
Several of the authors were students in a neurophysiology lab class, whose project was to address how 4-AP and TEA influence the activity of proprioceptive neurons in a crab model. This course project is part of a new trend in teaching science to undergraduates23. Course-based Undergraduate Research Experiences (CUREs) are relatively new and an approach being adopted by science educators in high schools and colleges24. Authentic research CUREs are referred to as ACUREs15,25.
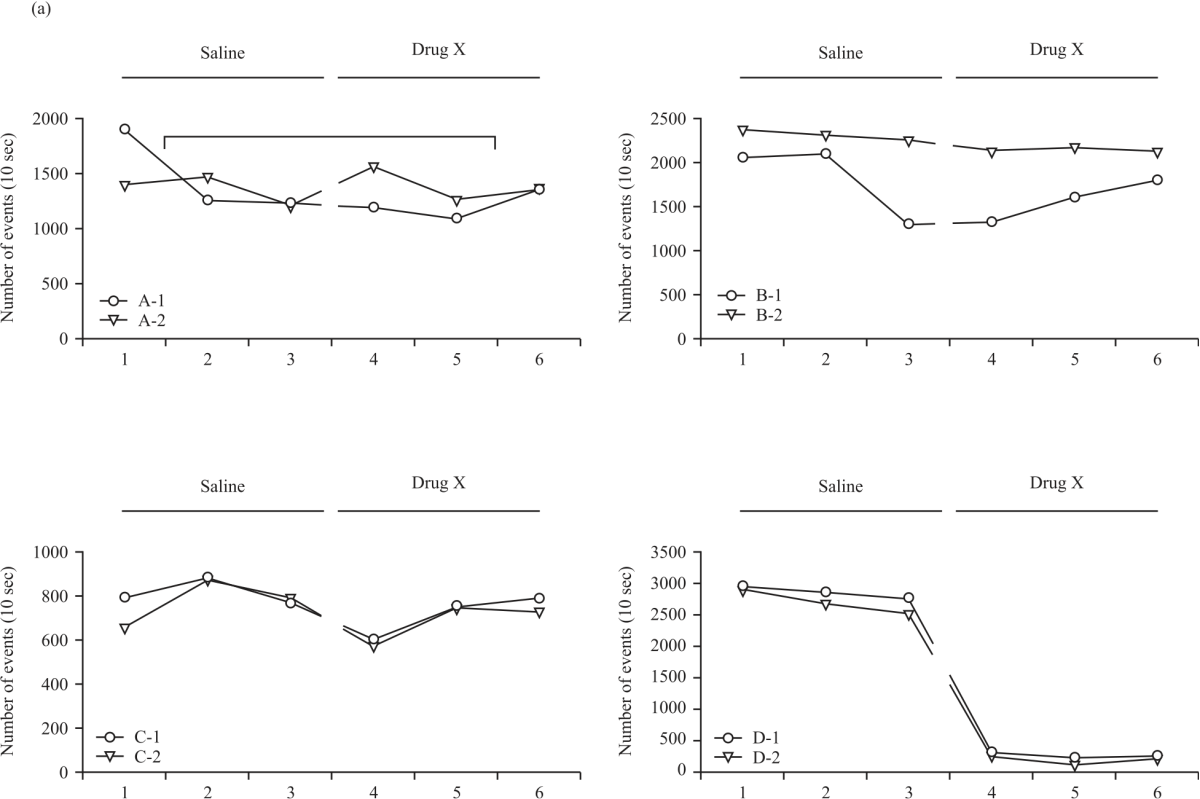 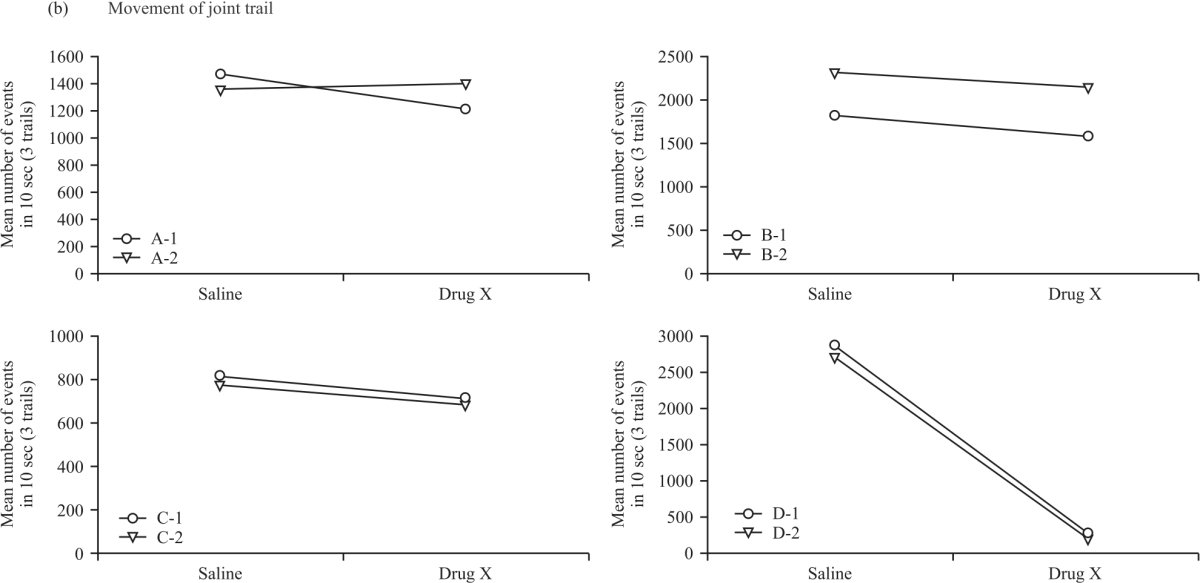 |
| Fig. S1(a-b): | Reproducibility of analysis by various groups of participants. The analysis was blind to the participants until completed. Eight groups, consisting of two students per group, were given a data set labelled A through D. The treatments were saline alone (groups A1 and A2), 4-AP (groups B1 and B2), TEA (groups C1 and C2) as well as a combined solution of 4-AP and TEA (groups D1 and D2). (a) General trends for each of the three trials in saline and the unknown are shown and (b) Average response for the three trials in each condition is shown for each treatment by the course participants |
REFERENCES
- Quartel, A., S. Turbeville and D. Lounsbury, 2010. Current therapy for Lambert-Eaton myasthenic syndrome: Development of 3,4-diaminopyridine phosphate salt as first-line symptomatic treatment. Curr. Med. Res. Opin., 26: 1363-1375.
CrossRefDirect Link - van Diemen, H.A.M., C.H. Polman, J.C. Koetsier, A.C.V. Loenen, J.J.P. Nauta and F.W. Bertelsmann, 1993. 4-aminopyridine in patients with multiple sclerosis. Clin. Neuropharmacol., 16: 195-204.
CrossRefDirect Link - Benton, B.J., S.A. Keller, D.L. Spriggs, B.R. Capacio and F.-C.T. Chang, 1998. Recovery from the lethal effects of saxitoxin: A therapeutic window for 4-aminopyridine (4-AP). Toxicon, 36: 571-588.
CrossRefDirect Link - Haghdoost-Yazdi, H., A. Faraji, N. Fraidouni, M. Movahedi, E. Hadibeygi and F. Vaezi, 2011. Significant effects of 4-aminopyridine and tetraethylammonium in the treatment of 6-hydroxydopamine-induced parkinson's disease. Behav. Brain Res., 223: 70-74.
CrossRefDirect Link - Vyshedskiy, A., T. Allana and J.W. Lin, 2000. Analysis of presynaptic Ca2+ influx and transmitter release kinetics during facilitation at the inhibitor of the crayfish neuromuscular junction. J. Neurosci., 20: 6326-6332.
CrossRefDirect Link - Leksrisawat, B., A.S. Cooper, A.B. Gilberts and R.L. Cooper, 2010. Muscle receptor organs in the crayfish abdomen: A student laboratory exercise in proprioception. J. Vis. Exp., Vol. 45.
CrossRefDirect Link - Kiernan, M.C., 2014. Impulse Conduction: Molecular Perspectives. In: Encyclopedia of the Neurological Sciences (Second Edition). Aminoff, M.J. and R.B. Daroff, Elsevier, Academic Press, ISBN 978-0-12-385158-1, Pages 688-690.
CrossRefDirect Link - Cooper, R.L., 2008. Mapping proprioceptive neurons on chordotonal organs in the crab, Cancer magister. Crustaceana, 81: 447-475.
CrossRefDirect Link - Dayaram, V., C. Malloy, S. Martha, B. Alvarez and I. Chukwudolue et al., 2017. The effect of CO2, intracellular pH and extracellular pH on mechanosensory proprioceptor responses in crayfish and crab. Am. J. Undergrad. Res., 14: 85-99.
CrossRefDirect Link - Dayaram, V., C.A. Malloy, S. Martha, B. Alvarez and I. Chukwudolue et al., 2017. Stretch activated channels in proprioceptive organs of crab and crayfish are sensitive to gadolinium but not amiloride, ruthenium red or low pH. Impulse, 2017: 1-17.
Direct Link - McCubbin, S., A. Jeoung, C. Waterbury and R.L. Cooper, 2020. Pharmacological profiling of stretch activated channels in proprioceptive neurons. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., Vol. 233.
CrossRefDirect Link - Malloy, C., V. Dayaram, S. Martha, B. Alvarez and I. Chukwudolue et al., 2017. The effects of potassium and muscle homogenate on proprioceptive responses in crayfish and crab. J. Exp. Zool. Part A: Ecol. Integr. Physiol., 327: 366-379.
CrossRefDirect Link - Stanback, M., A.E. Stanback, S. Akhtar, R. Basham and B. Chithrala et al., 2019. The effect of lipopolysaccharides on primary sensory neurons in crustacean models. IMPULSE, Vol. 16.
Direct Link - Stanley, C.E., R. Adams, J. Nadolski, E. Amrit and M. Barrett et al., 2020. The effects of tricaine mesylate on arthropods: Crayfish, crab and drosophila. Invertebr. Neurosci., Vol. 20.
CrossRefDirect Link - Judge, S.I.V., J.M. Lee, C.T.B.J. and P.M. Hoffman, 2006. Voltage-gated potassium channels in multiple sclerosis: Overview and new implications for treatment of central nervous system inflammation and degeneration. J. Rehabil. Res. Dev., 43: 111-122.
CrossRefDirect Link - Majeed, Z.R., J. Titlow, H.B. Hartman and R. Cooper, 2013. Proprioception and tension receptors in crab limbs: Student laboratory exercises. J. Visualized Exp., Vol. 80.
CrossRefDirect Link - Baierlein, B., A.L. Thurow, H.L. Atwood and R.L. Cooper, 2011. Membrane potentials, synaptic responses, neuronal circuitry, neuromodulation and muscle histology using the crayfish: Student laboratory exercises. J. Vis. Exp., Vol. 18.
CrossRefDirect Link - Atkins, D.E., K.L. Bosh, G.W. Breakfield, S.E. Daniels and M.J. Devore et al., 2021. The effect of calcium ions on mechanosensation and neuronal activity in proprioceptive neurons. NeuroSci, 2: 353-371.
CrossRefDirect Link - Pankau, C., J. Nadolski, H. Tanner, C. Cryer and J.D. Girolamo et al., 2022. Examining the effect of manganese on physiological processes: Invertebrate models. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., Vol. 251.
CrossRefDirect Link - Baker, M.D., 2013. Potential therapeutic mechanism of K+ channel block for MS. Multiple Sclerosis Relat. Disord., 2: 270-280.
CrossRefDirect Link - Rus, H., C.A. Pardo, L. Hu, E. Darrah and C. Cudrici et al., 2005. The voltage-gated potassium channel kv1.3 is highly expressed on inflammatory infiltrates in multiple sclerosis brain. Proc. Nat. Acad. Sci., 102: 11094-11099.
CrossRefDirect Link - Linn, M.C., E. Palmer, A. Baranger, E. Gerard and E. Stone, 2015. Undergraduate research experiences: Impacts and opportunities. Science, Vol. 347.
CrossRefDirect Link - Bakshi, A., L.E. Patrick and E.W. Wischusen, 2016. A framework for implementing course-based undergraduate research experiences (CUREs) in freshman biology labs. Am. Biol. Teach., 78: 448-455.
CrossRefDirect Link - Wycoff, S., K. Weineck, S. Conlin, C. Suryadevara and E. Grau et al., 2018. Effects of clove oil (eugenol) on proprioceptive neurons, heart rate, and behavior in model crustaceans. Impulse, Premier J. Undergraduate Publ. Neurosci., Vol. 2018.
Direct Link