
Nicole T. Marguerite
Department of Biology and Center of Muscle Biology, University of Kentucky, Lexington, 40506-0225 KY, USA
Shelby McCubbin
Department of Biology and Center of Muscle Biology, University of Kentucky, Lexington, 40506-0225 KY, USA
Robin L. Cooper
Department of Biology and Center of Muscle Biology, University of Kentucky, Lexington, 40506-0225 KY, USA
LiveDNA: 1.523
Journal of Pharmacology and Toxicology
Year: 2021 | Volume: 16 | Issue: 1 | Page No.: 37-46







ABSTRACT
Background and Objective: Invertebrates to mammals attempt to maintain heart rate under different physiological and pathological conditions by directly stretching myocytes as well as hormonal and neural regulation. Contractility increases with stretching of a heart tube in insect larvae as well as for the large ventricles of a mammal. This is a result of stretch activated ion channels, one type being transient receptor protein-ankyrin (TrpA) receptors. These TrpA receptors are known to increase in expression in cardiac pathologies of mammals, but it is not known if this is beneficial or detrimental. Hence the objective of the current study was to increase expression of the mechanosensitive channels (TrpA1) in the larval Drosophila heart to determine if they were responsible for maintaining the heart rate during pulsatile perfusion. Materials and Methods: Drosophila lines were constructed so the F1 generation selectively expressed TrpA1 in cardiac tissue. Larvae were dissected to expose the hearts to pulsatile perfusion for 4 hrs. The heart rates were counted and compared for significance using an ANOVA or Wilcoxon rank sum test. Results: Pulsatile supra-perfusion helps to maintain heart beats for over four hrs in situ which allows prolonged physiological and pharmacological studies. The parental (UAS-TrpA1) and over expressors (Tinc>TrpA1) of receptors relatively maintained heart rate at 20°C for about 4 hrs. At 30°C the parental line (UAS-TrpA1) was able to maintain heart rate better than the over expressors (Tinc>TrpA1). Conclusion: The TrpA1 blocker, HC-030031 at 30 μM, 100 μM and 1 mM, did not block the induced depolarization of body wall muscle at 30°C for larvae expressing TrpA1 receptors. Controls also did not show an effect with exposure to HC-030031.
PDF Abstract XML References Citation
How to cite this article
Nicole T. Marguerite, Shelby McCubbin and Robin L. Cooper, 2021. Mechanosensitive Proteins in Regulating Heart Rate in Larval Drosophila. Journal of Pharmacology and Toxicology, 16: 37-46.
DOI: 10.3923/jpt.2021.37.46
URL: https://scialert.net/abstract/?doi=jpt.2021.37.46
DOI: 10.3923/jpt.2021.37.46
URL: https://scialert.net/abstract/?doi=jpt.2021.37.46
INTRODUCTION
Transient receptor potenital (Trp) family of channels are a broad category, with some acting as stretch activated channels as well as serving as thermal receptors1. TrpA (TrpA - ankyrin) receptors are known to be present in vascular smooth muscle, endothelial cells and myocytes associated with the heart as well as pancreatic cells2, the spleen, and even in reproductive tissues3of mammals.
The mammalian heart has an amazing ability to self-regulate in physiological and pathological conditions by increasing contractility due to stretching of the ventricles (i.e., Frank Starling principle4), hormonal control and direct neural innervation. When a higher order pacing region fails, the heart will maintain electrical pacing in various regions to sustain some cardiac output5,6. There are also cellular changes of the heart which occur in pathological conditions, such as an increased expression of Trp proteins7,8. It is not known if this potential increase in expression in heart tissue is a beneficial compensation or whether it may contribute to the pathology9,10. However, there is some controversy in the functional expression in human pathological tissue and in rodent models of the Trp subtypes11.
The TrpA1 subtype of receptors are expressed in sensory neurons to detect thermal sensation, and when activated they elicit a behavioral response to internal as well as external environmental temperatures12. A recent study examined the effect of overexpression of TrpA1 in the larval heart of Drosophila and the effect of temperature on the heart rate. The heart rate increased substantially from 20-30°C in both control background strains and with the overexpression of TrpA1, however, the overexpressors stopped beating in a minute and remained in a diastolic state at 37°C. The heart rate recovered upon returning the temperature to 20°C13. Thus, the TrpA1 receptors can have a differential impact on heart rate depending on the temperature.
The larval Drosophila heart tube is myogenic and is regionally defined as the heart and aorta. Ostia are valves located in the heart to allow hemolymph to flow in from the body14. The heart pumps the hemolymph through a valve into the aorta, which directs flow to the anterior region of the larvae15. The master pacemakers are located on the most posterior region of the heart16. However, if the aortic tube is transected from the heart, then the two sections will pace on their own with the aortic tube pacing at a lower rate17.
In early stages of larval Drosophila, the heart is not neuronally innervated which allows one to examine the direct effects on heart tissue independent of neural regulation18. A modified physiological saline, with tightly controlled pH, maintains the larval heart to beat for an hour after in situ exposure to saline but will start to decrease beating in about 2 hrs19. The ability to dissect the larvae and replace the hemolymph with a defined saline allows one to examine the effects of pharmacological compounds on cardiac function. Thus, endogenous hormones and varied ionic composition in intact larvae is controlled for with an in situ preparation flushed with defined saline. In experiments examining various ionic compositions of saline to maintain the beating of the larval heart, it was noted that a pulsatile supra-perfusion significantly enhanced the duration of the heart to keep beating for up to 2 hrs19. This current study addressed pulsatile supra-perfusion for 4 hrs, which is the longest time reported maintaining cardiac function for in situ Drosophila larval preparations. The antagonist known to block the mammalian isoform of the TrpA1 receptors in rabbits at 10μM, HC-030031, was also examined in a line overexpressing TrpA1 in body wall muscles so that electrophysiological measures could be used to know if the ionic flux in the TrpA1 was blocked.
Considering that TrpA1 receptors also function as stretch activated channels and that a pulsatile perfusion of the heart helps to maintain the beating of the heart, the objective was to examine whether enhanced expression of TrpA1 receptors in the larval heart benefited the pulsatile influence in maintaining heart rate. This experimental manipulation in driving heart contraction with TrpA1 is similar in concept of being able to drive cardiac function with novel techniques such as optogenetics with light sensitive ion channels20-24.
MATERIALS AND METHODS
The experimental work was conducted in Lexington, Kentucky USA over years 2020 to 2021.
Drosophila lines: The over expression in the heart of the TrpA1 receptor was performed by crossing with female virgins of UAS-TrpA1 (BDSC stock # 26263). The heart-specific strain Tinc-Gal4 (on II)25 was used for expression of TrpA1 and is referred to as Tinc>TrpA1. UAS-TrpA1 and Tinc-Gal4 alone were used for control comparisons for heart rate measures. The overexpression in the body wall muscles (i.e., mesoderm) of the TrpA1 receptor was performed by crossing homozygous 24B-Gal4 (III) (BDSC stock # 1767) with female virgins of UAS-TrpA1 (BDSC stock # 26263). Progeny carrying one copy each of GAL4 driver and UAS-TrpA1, referred to as 24B>TrpA1 was used for physiological analysis. UAS-TrpA1 alone were used for control comparisons of the body wall muscles.
Dissection and solutions: To expose the larval heart tube in situ the procedures were followed as described in de Castro et al.26. The saline contained (mM): 1.0 CaCl2·2H2O, 70 NaCl, 20 MgCl2, 5 KCl, 10 NaHCO3, 5 trehalose, 115 sucrose, 25 5-N,N-bis(2-hydroxyethyl)-2-aminoethane sulfonic acid (BES).
The saline was aerated and the pH corrected to 7.1 with NaOH (1 M) or HCl (1 M) at room temperature of 20-21°C. Accumet model 10 pH meter (Fisher Scientific) and Ag/AgCl glass electrode was used. All chemicals listed above were obtained from Sigma-Aldrich (St. Louis, MO).
Pharmacological agents: HC-030031 (#H4415, (1,2,3,6-Tetrahydro-1,3-dimethyl-N-[4-(1-methylethyl)phenyl]-2,6-dioxo-7H-purine-7-acetamide, 2-(1,3-Dimethyl-2,6-dioxo-1,2,3,6-tetrahydro-7H-purin-7-yl)-N-(4-isopropylphenyl) acetamide), a known blocker of TrpA1 receptors was used at a concentration of 30 μM as this was shown to block TrpA127. Given the solubility is 10 mg mL–1 in DMSO, a stock was made with 10 mg mL–1 in DMSO and then diluted in HL3 modified saline to a concentration of 30 μM, 100 μM and 1 mM. All salts and compounds were obtained from Sigma (St. Louis, MO, USA).
Heart rate measurements: Heart Rate (HR) was counted by direct observation through a dissecting microscope as well as recordings captured on video (Movie 1). Experiments were performed with pulsatile saline perfusion.
A flow rate of 1.6 mL min–1 was delivered with a continuous perfusion pump (Master Flex, Vernon Hills, IL)19.
With the small recording dishes this rate of flow produces a substantial disturbance of fluid around the heart. For a visualization of the recording dish and dissection procedures see Cooper et al.28. To examine the effects of the antagonist, the bathing medium was exchanged from saline containing DMSO without HC-030031 and to saline with HC-030031 dissolved in DMSO and further recording at 21°C or 30°C.
The heart rates in each preparation and for each condition are shown. The raw values illustrate the variability among the preparations. It is important to illustrate the raw values as compared to a percent change in rates as a percent change can be large when the heart rate is low as compared to a high rate increasing by the same amount. In addition, it is easier to compare to past and future literature with when raw values are presented.
Electrical measures of body wall muscle: The effect of HC-030031 on blocking TrpA1 was examined by observing the effects on TrpA1 receptors expressed in body wall muscles. This was accomplished by changing the saline at 20-30°C without or with HC-030031 in the parental TrpA1-UAS line as well as 24B> TrpA1. This was performed as previously described13. In brief, body wall muscles (m6) of third instar larvae were impaled with a sharp intracellular electrode (30-40 M resistance) filled with 3 M KCl. An Axoclamp 2B (Molecular Devices, Sunnyvale, CA, USA) amplifier and 1 × LU head stage was used. The bathing saline was initially 20°C and then exchanged to 30°C while recording the membrane potential. The pH was monitored at 30°C at a pH of 7.1 noting that the pH was maintained, likely because of the high concentration of BES buffer used in this HL3-modified saline.
Statistical analysis: The alteration in heart rate were analyzed with an ANOVA using a Normality Test (Shapiro-Wilk) and a Kruskal-Wallis One Way Analysis of Variance on Ranks. The changes on the resting membrane potentials of the body wall muscles were analyzed with a rank sum non-parametric test.
RESULTS
To determine if the larvae heart would remain viable for a prolonged time with supra-perfusion, the heart beat was examined every 30 min for up to 4 hrs. This is the longest time we are aware of the in situ heart of Drosophila to be monitored. This was performed for the parental TrpA1-UAS line as well as Tinc>TrpA1 with and without activation of the TrpA1 channel by temperature.
20°C supra-perfusion: At 20°C with supra-perfusion the heart rate did gradually dimish over the 4 hrs for both the parental TrpA1-UAS line as well as Tinc>TrpA1 (Fig. 1 A1, B1). Due to the variation in the heart rates among the preparations, a percent change from the initial rate was used for normalization to make comparisons (Fig.1 A2, B2). There was no significant difference in the average percent change over time between time points for the parental TrpA1-UAS as compared to the Tinc>TrpA1 line (Fig. 1C). At 20°C there is little to no activation of the TrpA1 receptor as no differences in heart rate were observed in earlier studies with parental lines TrpA1-UAS and two heart specific lines for expression of TrpA1 (i.e., Hand4.2>TrpA1 and Tinc>TrpA1) as well as two different RNAi lines for TrpA1 (UAS-TrpA1-RNAi) and with w1118 combined with heterozygous 24B-Gal4 when examined at 20°C13.
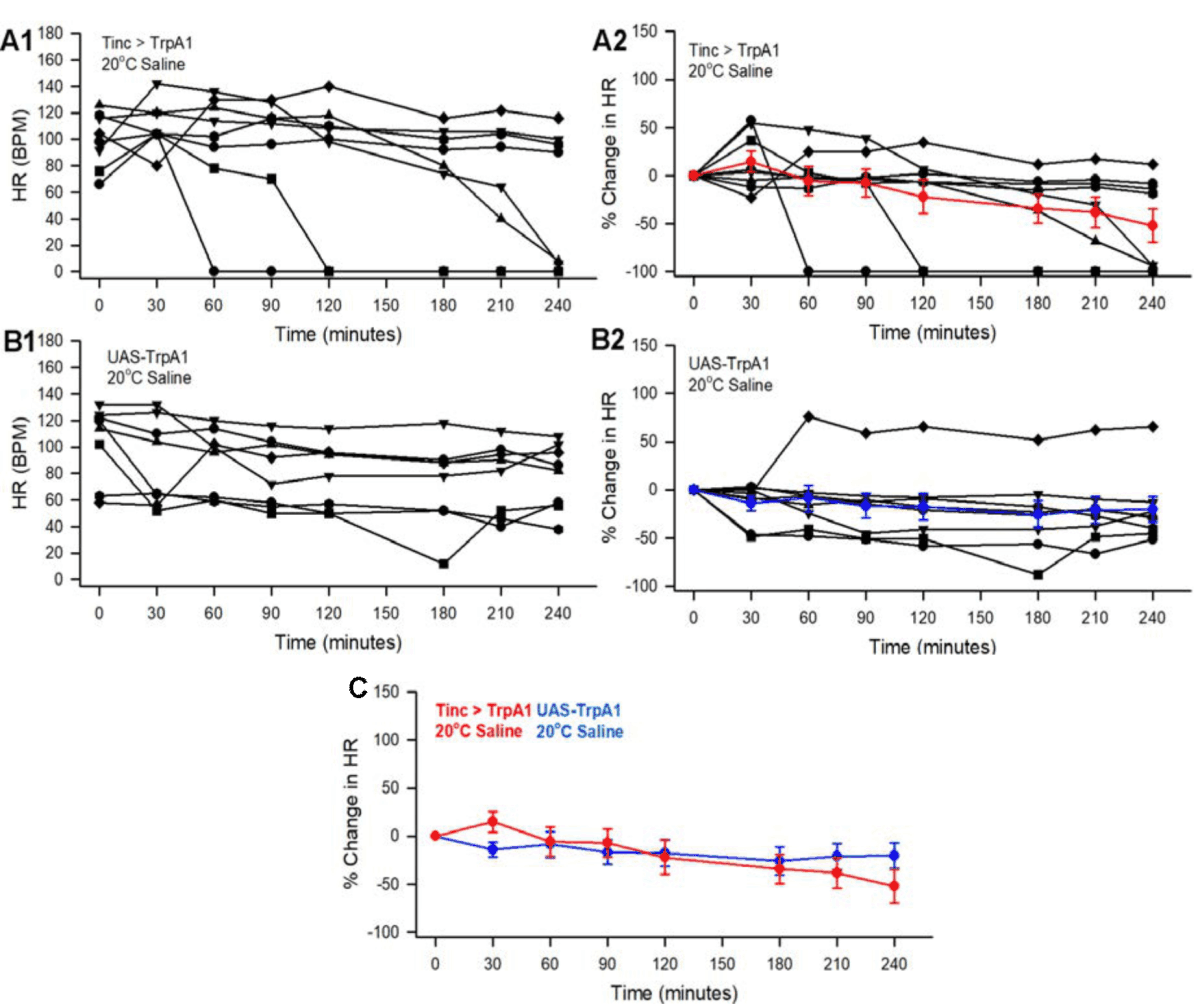 |
| Fig. 1(A-C): | Effect of 20°C on heart rate for larvae expressing TrpA1 with pulsatile supra-perfusion for 4 hrs. (A1) The heart rates for preparations monitored every 30 min for 4 hrs heart specific line (Tinc>TrpA1) expressing TrpA1 and the (B1) parental line the TrpA1 line (UAS-TrpA1). (A2 and B2) Percent change in heart rate from the initial reading for the rates shown in A1 and B1. The colored lines and symbols are the average (+/-SEM) of the recordings. (C) The average (+/-SEM) in the percent changes for all the preparations of the Tinc>TrpA1 and UAS-TrpA1 are illustrated together |
30°C supra-perfusion: The heart maintained beating for 4 hrs at 30°C for the Tinc>TrpA1 and parental line (UAS-TrpA1) with rates slowly decreasing over time more for the Tinc>TrpA1. To examine if the known antagonist HC-030031 (HC) altered the effect of temperature for the TrpA1 receptors, the solvent used for HC-030031 (i.e., DMSO) was used for the background control as well as Tinc>TrpA1 without exposure to HC-030031 (Fig. 2A1, B1, C1). Similar to the preparations monitored at 20°C, a percent change from the initial rate was used for normalization to make comparisons (Fig. 2A2, B2, C2). There were no significant differences for Tinc>TrpA1 preparations treated with HC-030031 as compared to the DMSO alone (Fig. 2D). However, there was a significant difference for both Tinc>TrpA1 with and without exposure to HC-030031 in comparison to UAS-TrpA1 after 3.5 and 4 hrs of perfusion (ANOVA, Normality Test (Shapiro-Wilk), Kruskal-Wallis One Way Analysis of Variance on Ranks, p<0.05. Fig. 2D). Within the first 30 min of monitoring heart rate, the rate increased for all preparations for Tinc>TrpA1 preparations treated with HC-030031 or with DMSO alone (P>0.05, non-parametric, Wilcoxon rank sum test, Fig. 2A1, A2, B1, B2) but not for the UAS-TrpA1 over time.
Body wall muscle: Since the contracting myocytes are difficult to maintain intracellular electrical recordings, the body wall muscles were used to examine the effect of raised temperature to 30°C and to examine if HC-030031 blocked the response of activating TrpA1 with temperature.
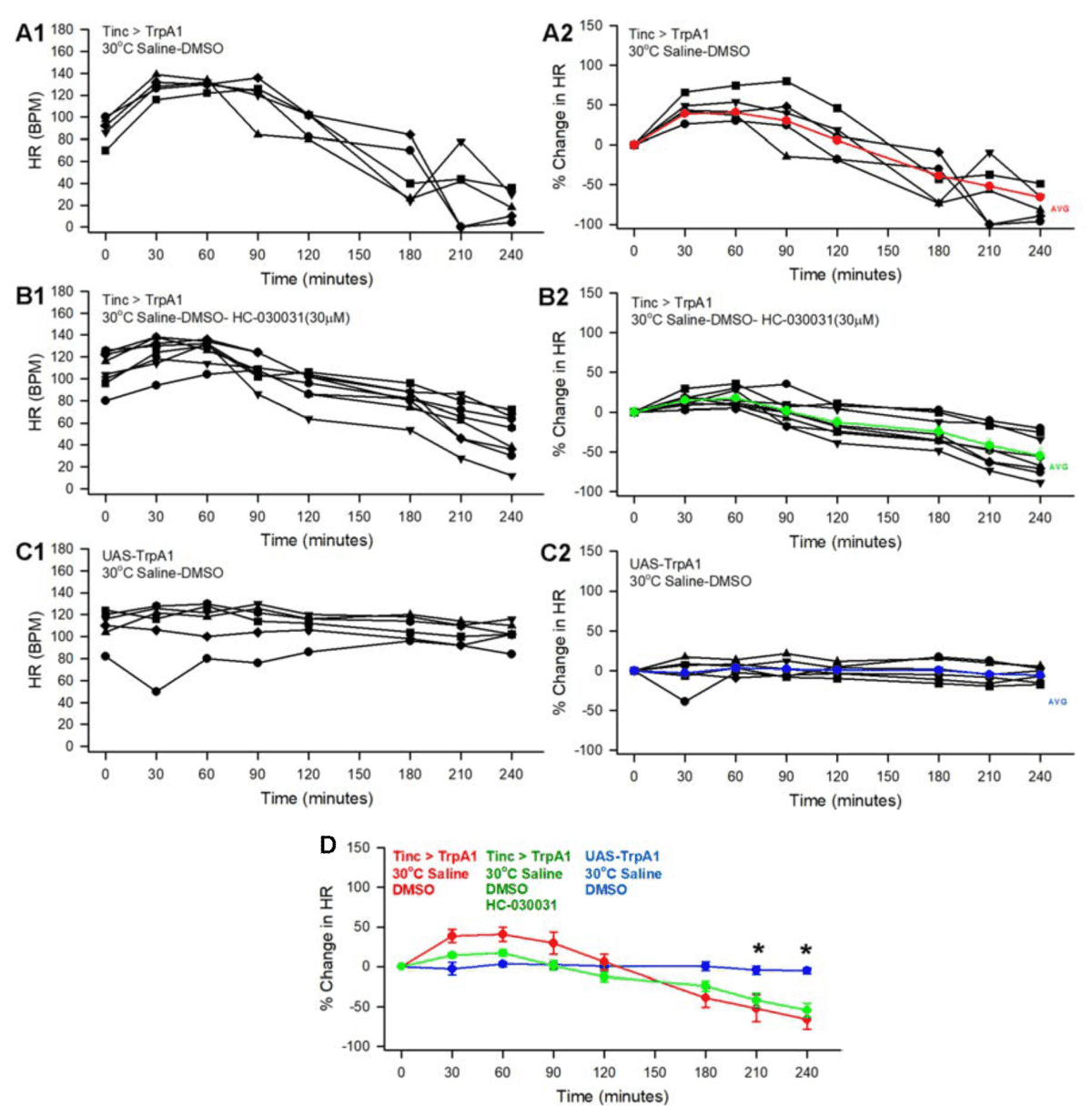 |
| Fig. 2(A-D): | Effect of 30°C on heart rate for larvae expressing TrpA1 with pulsatile supra-perfusion for 4 hrs and the effect of exposure to a HC-030031. (A1) The heart rates for preparations monitored every 30 min for 4 hrs heart specific line (Tinc>TrpA1) expressing TrpA1 with DMSO, (B1) and for the exposure to HC-030031 which was dissolved in DMSO as well as the (C1) parental line the TrpA1 line (UAS-TrpA1) in saline containing DMSO. (A2, B2 and C2) Percent change in heart rate from the initial reading for the rates shown in A1, B1 and C1. The colored lines and symbols are the average (+/-SEM) of the recordings. (D) The average (+/- SEM) in the percent changes for all the preparations of the Tinc>TrpA1with and without exposure to HC-030031and UAS-TrpA1 are illustrated together |
TrpA1 is readily expressed in skeletal muscle with use of the 24B line and has previously been examined for the effect on membrane potential at 27°C, 30°C and 37°C13 but not for the effect of TrpA1 antagonist with HC-030031.
The effect of DMSO, which was used to dissolve HC-030031 prior to adding to the physiological saline, had no effect on membrane potential as well as DMSO with HC-030031 at 20°C. The parental line (UAS-TrpA1) of the 24B>TrpA1 cross did not show any significant change in membrane potential at 20°C when switching the saline to DMSO containing saline (1 minute) followed by the same concentration of DMSO along with HC-030031 and returning to saline with DMSO (N = 6, p>0.05 rank sum non-parametric analysis).
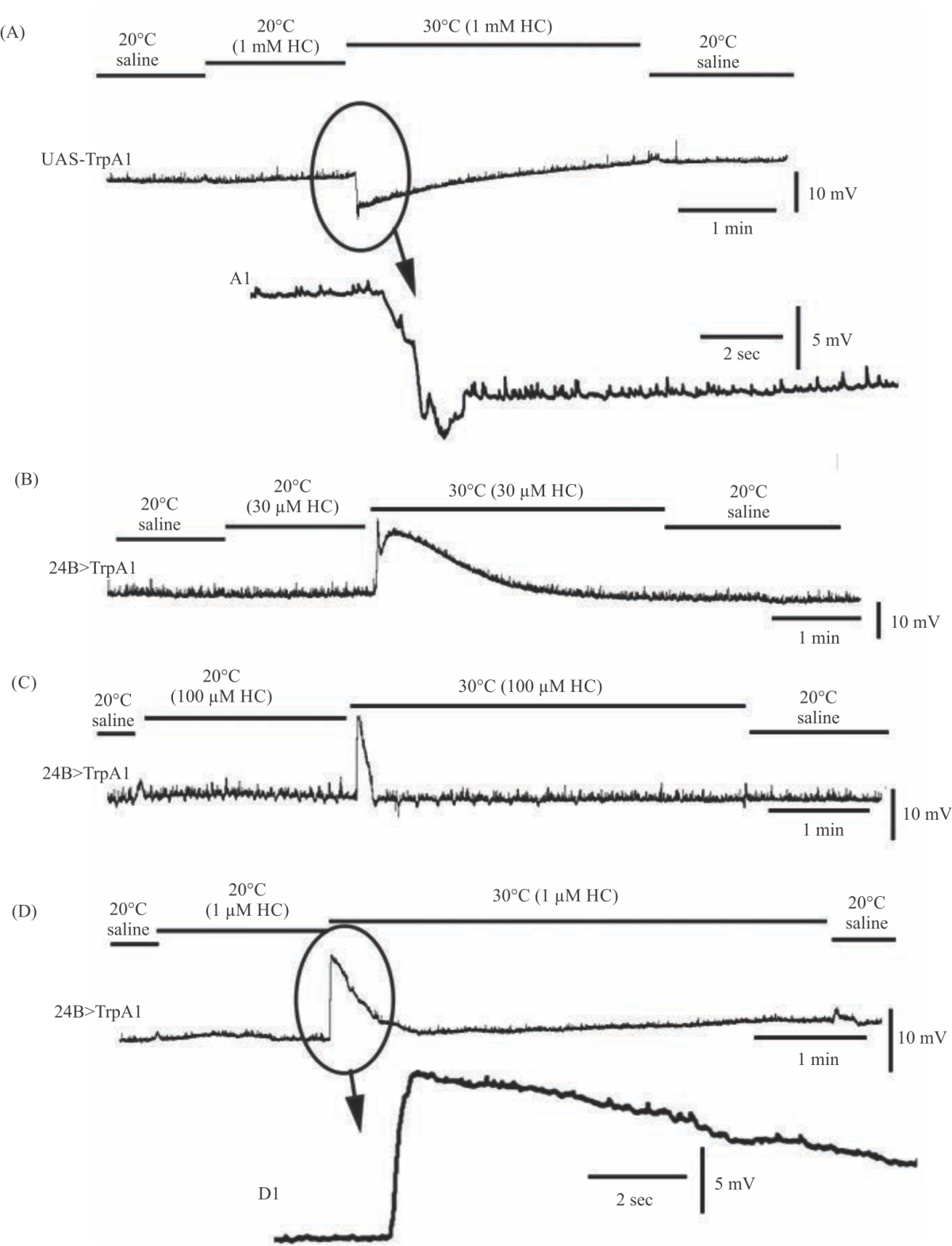 |
| Fig. 3(A-D): | Membrane potential of body wall muscles in TrpA1 overexpressing larvae at while activating TrpA1 and examining the putative blocker, HC-030031 (HC), of the TrpA1 channel. (A) The background line of UAS-TrpA1 at 30°C results in a rapid hyperpolarization of the membrane potential. The HC does not block this normal response. (B) The 24B>TrpA1 expressing line rapidly depolarized with exposure to 30°C and the HC did not block the depolarizing response at 30 μM, (C) 100 μM or even (D) 1 mM. Inserts A1 and D1 are enlarge segments of the indicated traces |
However, when exposed to 30°C the membrane did hyperpolarize with DMSO containing saline as well as with saline containing DMSO and HC-030031 (N = 6, p<0.05 rank sum non-parametric analysis). This is as expected considering increased temperature results in hyperpolarization of the muscle on its own13. There is variation among preparations in the effects of acute exposure to saline at 30°C, but are similar to earlier report13. To ensure HC-030031had time to be exposed to the muscle prior to changing the saline to 30°C, the saline with DMSO and HC-030031 at 20°C was exchanged in the bath for approximately 1 min and then the preparation was flushed with saline containing DMSO and HC-030031 at 30°C for about 3 min. Afterwards, the bath was exchanged back to saline without HC-030031 at 20°C (Fig. 3A). The same trends occurred in all preparations (N = 7).
The TrpA1 expressing larvae (24B>TrpA1) did not show any trends upon switching the saline to DMSO containing saline (1 minute) followed by the same concentration of DMSO along with HC-030031 and returning to saline with DMSO at 20°C (N = 6, p>0.05 rank sum non-parametric analysis). Preparations exposed to saline containing DMSO or DMSO and HC-030031 at 30°C also all depolarized in the same range. The same trends were observed for HC-030031 at 30 μM, 100 μM and 1 mM (Fig. 3B, C, D respectively).
DISCUSSION
This study showed that pulsatile supra-perfusion aided the larval Drosophila heart to maintain beating for over 4 hrs while in situ. This provides an opportunity for experimentation in future physiological and pharmacological studies over longer periods of time. In addition, it was demonstrated that when TrpA1 receptors are activated with heat at 30°C the heart rate will decrease likely due to Ca2+ overloading. Lastly, it was concluded that the established TrpA1 blocker, HC-030031, at various concentrations, did not block TrpA1 receptors when expressed in body wall muscles.
The heart of the genetical amenable model, Drosophila, has allowed one to examine the function of various altered genes which influence ionic properties and homeostatic mechanisms29-31 in the intact organism as well as in-situ. In addition, the Drosophila heart model serves as a testing ground for pharmacological compounds. Some applications use the larval or the pupal stage for assessing the effect of the heart rate. Considering that the heart is very susceptible to modulation by compounds released into the hemolymph, such as serotonin, octopamine and acetylcholine17,32-37, the dissected larvae bathed in a controlled physiological saline26,38 provides a more controlled environment than in vivo studies. However, it has been challenging in the past to be able to maintain a heartbeat in salines used for studying synaptic transmission and for culturing Drosophila cells. The heartbeat is particularly sensitive to pH. Therefore the development of a saline at pH 7.1 with strong buffering has allowed the heartbeat to be monitored for a longer period of time than with the use of the original HL3 saline and other previously examined salines26. In addition, there is quite a bit of variation in the initial rates of the heart upon dissection. The ability to also supra-perfuse the heart tubes of the larvae has also allowed more prolonged pharmacological studies to be performed in the defined saline devoid of endogenous modulators within intact larvae. Previously supra-perfusion of only 2 hrs was investigated for the larval hearts, however, this current study has now illustrated that the hearts are generally able to be maintained up to 4 hrs. Considering the pulsatile perfusion of the larval heart tube is key in maintaining the heart beat independent of endogenous modulators or added modulators, it is apparent the pulsatile stretching of heart is the mechanisms to maintenance of the beating rhythm. Considering that TrpA1 channels may up regulate in expression in mammalian hearts during pathological c onditions8 and that TrpA1 are also known to be activated by m echanical deformation1 as well as an increase in temperature, this study set out as a proof of concept to determine if the presence of the TrpA1 would have an effect on the heartbeat without a change in temperature.
The expression of TrpA1 in the heart tube with a heart specific driver (Tinc) illustrated that the heartbeat was better maintained over an extended period of time with pulsatile supra-perfusion. However, if the TrpA1 channels is activated by heat at 30°C continuously then the heartbeat is depressed as compared to preparation not expressing TrpA1 receptors. It is potentially feasible that expression of TrpA1 in mammalian hearts may be a compensatory mechanism to maintain contractility in pathological conditions by enhancing the Frank Starling effect4. However, if over stimulation such as by an increase in heat activation, there may be a negative impact in maintaining a consistent heartbeat. Considering the receptor is defined as transient in its action7,8, one might assume the change in temperature might only result in a transient effect on the heartbeat. The rapid rise in heart rate does occur upon increasing the temperature from 20-30°C as well as from 20-37°C but the effect is short lived with the heartbeat stopping in a shorter time than for control larvae and background strains13. Thus, it is likely the TrpA1 is still being activated during the continuous exposure to heat. Potentially the pulsatile action of the supra-perfusion may transitively activate the TrpA1 protein as the rate of the calcium exchanger or a calcium pump may provide time for the receptors to regain a transient function, however, if the receptors are able to be continuously activated throughout the supra-perfusion without the additional heat activation than it is likely they are helping to maintain the rhythm with enough time for the myocytes to regulate the homeostasis the ionic fluxes. If the flux is too large, due to the additional heat exposure at 30°C, the myocytes are slowly not able to maintain the ionic balance. At 37°C this dysfunctional state occurs more rapidly13.
The rundown of heart rate over time at 20°C for the UAS-TrpA1 and Tinc>TrpA1, even with pulsatile supra-perfusion, may well be due to the lack of modulators which naturally occur in the hemolymph within intact larvae. However, in the previous study with TrpA1 expression in the myocytes, serotonin was not able to stimulate the heart to beat even though it was in diastolic state while at 37°C13 and the same phenomenon of heart rate cessation and lack of sensitivity to serotonin occurs if the myocytes are over stimulated with optogenetic approaches24.
To examine the potential of the pharmacological agent HC-030031 in blocking the Drosophila isoform of TrpA1 receptor, the expression in the body wall muscles allowed us to readily measure the membrane potential of the cells while activating TrpA1 with warm saline. The activation of TrpA1 in the body wall muscle produces a transient depolarization of the membrane potential. When raising the saline temperature for UAS-TrpA1, 24B>TrpA1-RNA or Canton-S larvae the membrane potential hyperpolarizes13. The hyperpolarization is most likely due in part to the changes on the equilibrium potential of the potassium leak channels39. So, the depolarized state through the activating the TrpA1 receptors is likely due to a larger effect than just the depolarization as the drive to the equilibrium potential of the potassium leak channels of a greater negative potential is counteracted with depolarization above the resting membrane potential. Considering the hyperpolarizing membrane potential in muscle for the background controls with higher temperature is not altered by HC-030031, this indicates that the compound is not altering the drive of the potassium leak channels. In addition, the depolarization of the TrpA1 expressing line is not blocked by exposure to HC-030031even at high concentration of 1 mM. It is reported that HC-030031 blocks the mammalian human isoform TrpA1 when expressed40. However, this compound did not inhibit the frog TRPA1 (fTRPA1) or the zebrafish TRPA1 activity when activated by cinnamaldehyde40.
We assume that HC-030031 is not a strong antagonist of the Drosophila TrpA1 isoform. The Drosophila genome has sixteen various TRP channels41. It is not known how many types of TRP channels are expressed in the larval heart and the range in stimuli (i.e., thermal, mechanical, chemical) to activate them.
Thus, further pharmacological investigations into potential agonist and antagonist of the Drosophila TrpA isoforms is warranted to better characterized these receptor subtypes for more detailed studies in the physiological function of these types of receptors.
There are likely some form (s) of stretch activated proteins expressed in the heart tube of the larval heart. A limitation in this study was that it is still not known what type of mechanosensation is responsible for the heart to maintain beating with the pulsatile perfusion. It was demonstrated that the TRP channel Painless42 expressed in Drosophila larval heart resulted in cessation of the heartbeat with activation by heat or by mechanical stimulation43. Activation of Painless pauses the heart rate but does not promote an increase in heart rate, thus, it would be feasible a different isoform of TRP receptor might be present or another type of stretch activated channel is present to be responsible for the maintenance of the heart rate due to the sensitivity of the pulsatile supra-perfusion. It is likely not the subtype of PIZEO stretch activated channel since RNAi for PIEZO did not alter the heart rate and the antagonist ruthenium red, streptomycin and tarantula toxin did not have an effect, however, amiloride and gadolinium did reduce heart rate but the effect is likely through other mechanisms than stretch activated channels19. Experiments are underway to screening several relatively new pharmacological agents (i.e., YODA 1, JEDI 2, OB 1 and DOOKU) which block different forms of stretch activated channels44.
CONCLUSION
It was illustrated that pulsatile supra-perfusion of the larval heart maintains the heart to beat for 4 hrs, which is prolonged as compared to a static bath. Using a heart specific driver (Tinc) to express the TrpA1 proteins did not compromise the heartbeat. If the TrpA1 channels were also activated with heat, the heart rate declined quicker as compared to hearts without the expression of TrpA1. The depolarization in body wall muscles with over expression of TrpA1 channels indicated that the TrpA1 were activated at 30°C.
SIGNIFICANCE STATEMENT
This study was done to find out that an increase in expression of the mechanosensitive channels (TrpA1) in the larval Drosophila heart are responsible for maintaining the heart rate during pulsatile perfusion. These findings may aid in addressing whether the increased expression of TrpA1 in pathological states in mammals could be beneficial in maintaining an ionotropic as well as chronotropic function of the heart. However, even if the mammalian heart is not expressing a TrpA1 in physiological healthy hearts as a proof of concept the effect of activating the TrpA1 via pulsatile perfusion is of interest to determine the effects in potentially stimulating cardiomyocytes.
ACKNOWLEDGMENT
Stocks obtained from the Bloomington Drosophila Stock Center (NIH P40OD018537) were used in this study. We thank Dr. Anthony Cammarato for suppling the heart-specific line (Tinc-Gal4). Chellgren Endowed Professor fund from the University of Kentucky to R.L.C. for supplies. The NRPA NEURO Scholars Fellowship (University of Kentucky) to Shelby McCubbin.
REFERENCES
- Hof, T., S. Chaigne, A. Récalde, L. Sallé, F. Brette and R. Guinamard, 2019. Transient receptor potential channels in cardiac health and disease. Nat. Rev. Cardiol., 16: 344-360.
CrossRefDirect Link - Cao, D.S., L. Zhong, T.H. Hsieh, M. Abooj, M. Bishnoi, L. Hughes and L.S. Premkumar, 2012. Expression of transient receptor potential ankyrin 1 (TRPA1) and its role in insulin release from rat pancreatic beta cells. PLoS ONE, Vol. 7.
CrossRefDirect Link - Kunert-Keil, C., F. Bisping, J. Krüger and H. Brinkmeier, 2006. Tissue-specific expression of TRP channel genes in the mouse and its variation in three different mouse strains. BMC Genomics, Vol. 7.
CrossRefDirect Link - Rhodes, S.S., A.K. Camara, M. Aldakkak, J.S. Heisner and D.F. Stowe, 2015. Stretch‐induced increase in cardiac contractility is independent of myocyte Ca2+ while block of stretch channels by streptomycin improves contractility after ischemic stunning. Physiol. Rep., Vol. 3, No. 8.
CrossRefDirect Link - Armoundas, A.A., R. Wu, G. Juang, E. Marbán and G.F. Tomaselli, 2001. Electrical and structural remodeling of the failing ventricle. Pharmacol. Ther., 92: 213-230.
CrossRefDirect Link - Pappano, A.J. and W.G. Wier, 2013. Cardiovascular Physiology. 10th Edn., Elsevier Inc. Pages: 304.
CrossRefDirect Link - Bodkin, J.V., P. Thakore, A.A. Aubdool, L. Liang and E.S. Fernandes et al., 2014. Investigating the potential role of TRPA1 in locomotion and cardiovascular control during hypertension. Pharmacol. Res. Perspect., Vol. 2.
CrossRefDirect Link - Andrei, S.R., M. Ghosh, P. Sinharoy, S. Dey, I.N. Bratz and D.S. Damron, 2017. TRPA1 ion channel stimulation enhances cardiomyocyte contractile function via a CaMKII-dependent pathway. Channels, 11: 587-603.
CrossRefDirect Link - Falcón, D., I. Galeano-Otero, E. Calderón-Sánchez, R.D. Toro and M. Martín-Bórnez et al., 2019. TRP channels: current perspectives in the adverse cardiac remodeling. Front. Physiol., Vol. 10.
CrossRefDirect Link - Lu, Y., H. Piplani, S.L. McAllister, C.M. Hurt and E.R. Gross, 2016. Transient receptor potential ankyrin 1 activation within the cardiac myocyte limits ischemia–reperfusion injury in rodents. Anesthesiology, 125: 1171-1180.
CrossRefDirect Link - Hoebart, C., N.S. Rojas‐Galvan, C.I. Ciotu, I. Aykac and L.F. Reissig et al., 2021. No functional TRPA1 in cardiomyocytes. Acta Physiol.
CrossRefDirect Link - Hamada, F.N., M. Rosenzweig, K. Kang, S.R. Pulver, A. Ghezzi, T.J. Jegla and P.A. Garrity, 2008. An internal thermal sensor controlling temperature preference in Drosophila. Nature, 454: 217-220.
CrossRefDirect Link - Marguerite, N.T., J. Bernard, D.A. Harrison, D. Harris and R.L. Cooper, 2021. Effect of temperature on heart rate for Lucilia sericata (syn Phaenicia sericata) and Drosophila melanogaster with altered expression of the TrpA1 receptors. Insects, Vol. 12.
CrossRefDirect Link - Rotstein, B. and A. Paululat, 2016. On the morphology of the Drosophila heart. J. Cardiovasc. Dev. Dis., Vol. 3.
CrossRefDirect Link - Lammers, K., B. Abeln, M. Hüsken, C. Lehmacher and O.E. Psathaki et al., 2017. Formation and function of intracardiac valve cells in the Drosophila heart. J. Exp. Biol., 220: 1852-1863.
CrossRefDirect Link - Curtis, N.J., J.M. Ringo and H.B. Dowse, 1999. Morphology of the pupal heart, adult heart, and associated tissues in the fruit fly, Drosophila melanogaster. J. Morphol., 240: 225-235.
CrossRef3.0.co;2-v target='_blank' class='btn btn-sm btn-outline-primary mr-3 mt-3'>Direct Link - Majeed, Z.R., A. Stacy and R.L. Cooper, 2014. Pharmacological and genetic identification of serotonin receptor subtypes on Drosophila larval heart and aorta. J. Comp. Physiol. B., 184: 205-219.
CrossRefPubMedDirect Link - Dulcis, D. and R.B. Levine, 2003. Innervation of the heart of the adult fruit fly, Drosophila melanogaster. J. Comp. Neurol., 465: 560-578.
CrossRefDirect Link - De Castro, C., J. Titlow, Z.R. Majeed, C. Malloy, K.E. King and R.L. Cooper, 2019. Mechanical and chemical factors required for maintaining cardiac rhythm in Drosophila melanogaster larva. J. Entomol., 16: 62-73.
CrossRefDirect Link - Arrenberg, A.B., D.Y.R. Stainier, H. Baier and J. Huisken, 2010. Optogenetic control of cardiac function. Science, 330: 971-974.
CrossRefDirect Link - Govorunova, E.G., S.R. Cunha, O.A. Sineshchekov and J.L. Spudich, 2016. Anion channelrhodopsins for inhibitory cardiac optogenetics. Sci. Rep., Vol. 6.
CrossRefDirect Link - Kopton, R.A., J.S. Baillie, S.A. Rafferty, R. Moss and C.M. Zgierski-Johnston et al., 2018. Cardiac electrophysiological effects of light-activated chloride channels. Front. Physiol., Vol. 17.
CrossRefDirect Link - O’Shea, C., A.P. Holmes, J. Winter, J. Correia and X. Ou et al., 2019. Cardiac optogenetics and optical mapping-overcoming spectral congestion in all-optical cardiac electrophysiology. Front. Physiol., Vol. 10.
CrossRefDirect Link - Stanley, C.E., A.S. Mauss, A. Borst and R.L. Cooper, 2019. The effects of chloride flux on Drosophila heart rate. Methods Protocols, Vol. 2, No. 3.
CrossRefDirect Link - Blice-Baum, A.C., A.C. Zambon, G. Kaushik, M.C. Viswanathan, A.J. Engler, R. Bodmer and A. Cammarato, 2017. Modest overexpression of FOXO maintains cardiac proteostasis and ameliorates age-associated functional decline. Aging Cell, 16: 93-103.
CrossRefDirect Link - De Castro, C., J. Titlow, Z.R. Majeed and R.L. Cooper, 2014. Analysis of various physiological salines for heart rate, CNS function and synaptic transmission at neuromuscular junctions in Drosophila melanogaster larvae. J. Comp. Physiol. A, 200: 83-92.
CrossRefDirect Link - Legrand, C., J.M. Merlini, C. de Senarclens-Bezençon and S. Michlig, 2020. New natural agonists of the transient receptor potential Ankyrin 1 (TRPA1) channel. Sci. Rep., Vol. 10.
CrossRefDirect Link - Cooper, A.S., K.E. Rymond, M.A. Ward, E.L. Bocook and R.L. Cooper, 2009. Monitoring heart function in larval Drosophila melanogaster for physiological studies. J. Vis. Exp., Vol. 33.
CrossRefDirect Link - Gu, G.G. and S. Singh, 1995. Pharmacological analysis of heartbeat in Drosophila. J. Neurobiol., 28: 269-280.
CrossRefDirect Link - Mohr, S.E. and N. Perrimon, 2019. Drosophila melanogaster: A simple system for understanding complexity. Dis. Models Mech., Vol. 12.
CrossRefDirect Link - Reiter, L.T., L. Potocki, S. Chien, M. Gribskov and E. Bier, 2001. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res., 11: 1114-1125.
CrossRefDirect Link - Dasari, S. and R.L. Cooper, 2006. Direct influence of serotonin on the larval heart of Drosophila melanogaster. J. Comp. Physiol. B, 176: 349-357.
CrossRefDirect Link - Majeed, Z.R., C.D. Nichols and R.L. Cooper, 2013. 5-HT stimulation of heart rate in Drosophila does not act through cAMP as revealed by pharmacogenetics. J. Applied Physiol., 115: 1656-1665.
CrossRefPubMedDirect Link - Malloy, C.A., K. Ritter, J. Robinson, C. English and R.L. Cooper, 2015. Pharmacological identification of cholinergic receptor subtypes on Drosophila melanogaster larval heart. J. Comp. Physiol. B, 186: 45-57.
CrossRefPubMedDirect Link - Malloy, C., J. Sifers, A. Mikos, A. Samadi, A. Omar, C. Hermanns and R.L. Cooper, 2017. Using optogenetics to assess neuroendocrine modulation of heart rate in Drosophila melanogaster larvae. J. Comp. Physiol. A, 203: 791-806.
CrossRefPubMedDirect Link - Titlow, J.S., J.M. Rufer, K.E. King and R.L. Cooper, 2013. Pharmacological analysis of dopamine modulation in the Drosophila melanogaster larval heart. Physiol. Rep., Vol. 1, No. 2.
CrossRefDirect Link - Zhu, Y.C., E. Yocom, J. Sifers, H. Uradu and R.L. Cooper, 2016. Modulatory effects on Drosophila larva hearts: Room temperature, acute and chronic cold stress. J. Comp. Physiol. A., 186: 829-841.
CrossRefPubMedDirect Link - Stewart, B.A., H.L. Atwood, J.J. Renger, J. Wang and C.F. Wu, 1994. Improved stability of Drosophila larval neuromuscular preparations in haemolymph-like physiological solutions. J. Comp. Physiol. A., 175: 179-191.
CrossRefDirect Link - Zhu, Y.C. and R.L. Cooper, 2018. Cold exposure effects on cardiac function and synaptic transmission at the neuromuscular junction in invertebrates. Int. J. Zool. Res., 14: 49-60.
CrossRefDirect Link - Gupta, R., S. Saito, Y. Mori, S.G. Itoh, H. Okumura and M. Tominaga, 2016. Structural basis of TRPA1 inhibition by HC-030031 utilizing species-specific differences. Sci. Rep., Vol. 6.
CrossRefDirect Link - Christensen, A.P. and D.P. Corey, 2007. TRP channels in mechanosensation: direct or indirect activation? Nat. Rev. Neurosci., 8: 510-521.
CrossRefDirect Link - Tracey Jr., W.D., R.I. Wilson, G. Laurent and S. Benzer, 2003. Painless, a Drosophila gene essential for nociception. Cell, 113: 261-273.
CrossRefDirect Link - Sénatore, S., V.R. Reddy, M. Sémériva, L. Perrin and N. Lalevée, 2010. Response to mechanical stress is mediated by the TRPA channel painless in the Drosophila heart. PLoS Genet., Vol. 6, No. 9.
CrossRefDirect Link - McCubbin, S., A. Jeoung, C. Waterbury and R.L. Cooper, 2020. Pharmacological profiling of stretch activated channels in proprioceptive neurons. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., Vol. 233.
CrossRefDirect Link