
J. Johnson
Laboratory of Food Safety and Toxicology, Alabama A and M University, Normal Al, 35872, United States
S. Ogutu
Laboratory of Food Chemistry, Alabama A and M University, Normal Al, 35872, United States
J. Boateng
Laboratory of Food Safety and Toxicology, Alabama A and M University, Normal Al, 35872, United States
LiveDNA: 1.12516
Journal of Pharmacology and Toxicology
Year: 2021 | Volume: 16 | Issue: 1 | Page No.: 22-36







ABSTRACT
Background and Objective: Edible flowers such as the daylily (Hemerocallis spp.) have been shown to have beneficial effects against chronic diseases. This study determined antioxidant activity and polyphenol contents of two cultivars of daylily flowers Barbara Mitchell (BM) and Elegant Explosion (EE). The in vitro bioactivities of EE extract were further examined in HepG2 cell lines. Materials and Methods: BM and EE daylily flowers extracts were prepared with aqueous ethanol and used to determine Total Phenolic Content (TPC), Total Flavonoid Content (TFC), Total Monomeric Anthocyanins (TMA), Condensed Tannins (CT) and antioxidant assays. Selected phytochemicals were also determined via HPLC/DAD. In vitro toxicity, apoptosis and enzymatic activities of EE extracts were assessed in HepG2 cells. Results: Results indicated EE showed significantly (p<0.05) higher polyphenol contents and antioxidant activity compared to BM. In vitro cytotoxicity indicated that EE extracts possess moderately weak toxicity towards HepG2 cells. Significant (p<0.05) increases in antioxidant activities were noted. Conclusion: With limited studies on polyphenol content and bioactivity of EE daylily flower, these results provide preliminary evidence and contribute to the promotion of edible flowers as a potential dietary supplement and nutraceutical for use as functional foods.
PDF Abstract XML References Citation
How to cite this article
J. Johnson, S. Ogutu and J. Boateng, 2021. Antioxidant and in vitro Toxicity Assessment of Elegant Explosion Daylily Flower (Hemerocallis spp.) Extract in Hepatic Cell Line. Journal of Pharmacology and Toxicology, 16: 22-36.
DOI: 10.3923/jpt.2021.22.36
URL: https://scialert.net/abstract/?doi=jpt.2021.22.36
DOI: 10.3923/jpt.2021.22.36
URL: https://scialert.net/abstract/?doi=jpt.2021.22.36
INTRODUCTION
A primary knowledge of edible flower preparations has been documented from accounts described in ancient literature1-3. For example, in 16th century Central Europe, fried batter-coated black elder (Sambucus nigra) flowers were common, as were dandelion flowers boiled with ersatz honey2-4. Ancient Romans enjoyed roses and lavender in sauces4,5. In Victorian-era England, roses were used as flavouring agents in teas, sweets, pastries, conserves and sauces4,5. Besides the wholesomeness and suitability in most aspects of culinary applications, edible flowers are abundant sources of nutrients such as carbohydrates, protein, fat, minerals and vitamins6,7. Additionally, edible flowers contain varying quantities of several bioactive compounds8-11. These bioactive compounds with antioxidant properties contribute to the medicinal soundness of edible flowers and thus support increased worldwide demand.
The daylily (Hemerocallis spp.) is an edible flower, which dates back thousands of years in sub-tropical Asia. The main diversity of the genus is centred in China, Korea and Japan, where they are used as medicine and food12. For culinary and consumption purposes, different parts of the flower are used as vegetables and incorporated into soups, stews, or eaten raw. Daylilies (Hemerocallis spp.) have shown strong antioxidant activities in vitro and vivo13-15. These beneficial effects are believed to be due to several novel bioactive compounds including naphthalene glycoside, stelladerol, caffeoylquinic acid derivatives, as well as other antioxidants such as gallic acid, rutin and catechin16,17. According to Rodriguez-Enriquez and Grant-Downton17, Hemerocallis spp. could have significant potential as a nutraceutical and the source of new compounds with biomedical activities. Thus, the daylily could be considered as a functional food and used in natural products and supplements to improve health.
Although the consumption of flowers is on the rise, it is concluded that there are still numerous aspects that need to be evaluated. This is especially important if it is to be considered as a functional food or nutraceutical. As highlighted by Granato et al.18, assigning a functional label to a potential food product or nutritional supplement requires completion of a proper assessment to attest its functionality using in vitro and in vivo methods. Given this, Granato et al.18 indicated that a wide variety of tests, both in vitro and in vivo, must be performed to determine efficacy and bioactivity as well as to assess the physicochemical and chemical properties of any food before stating it is functional.
This study aims to determine the polyphenolic contents and antioxidative activity of daylily cultivars Barbara Mitchell (BM) and Elegant Explosion (EE) and also, to assess their acute in vitro toxicity potential (cytotoxicity, apoptotic and antiproliferative properties) in HepG2 hepatic cell line.
MATERIALS AND METHODS
Study area: The study was carried out in the Department of Food Science at Alabama A and M University, Normal AL, United States from September, 2016-December, 2018.
Materials and reagents
Flowers: Barbara Mitchell (BM) and Elegant Explosion (EE) daylily plantlets were obtained from a local nursery (Bennet Nurseries) that cultivates and sells edible flowers from organic production. (The plantlets were confirmed by Dr. Patti Coggins, who has a long history of working with Hemerocallis spp.). The plantlets were cultivated and cared for in the greenhouse at Alabama A and M University, United States until harvesting.
Chemicals: All chemicals and reagents that were used in this study were of pure analytical grade and purchased from Fisher Scientific (Suwannee, GA) and Sigma Aldrich (St. Louis, MO). Kits and other supplies were purchased from Promega, Thermo Fisher Scientific (Madison, WI) and Cayman Chemical Company (Ann Arbor, MI). Catechin, Rutin, Gallic acid and Chlorogenic acid standards were acquired from Indofine Chemical Company (Hillsborough, NJ).
Harvesting and extraction of polyphenols from daylily flowers: Barbara Mitchell and Elegant Explosion daylily flowers were harvested at full bloom in the morning for a week. The pulling method was used to combine several batches over the days of harvest. Flowers were placed in individual plastic containers with absorbent papers at the bottom to prevent any lymph leakage due to guttation. The harvested flowers were placed on ice and transported to the laboratory in refrigerator bags and stored at -20°C immediately. The flowers were analyzed within 1 week from the collection day.
Extraction procedure: Approximately 70 g of flowers were weighed and homogenized in 200 mL of 80% ethanol. The homogenate was stored in the dark overnight. The supernatant was removed by centrifugation (3000 rpm at 4°C for 20 min). The extraction procedure was repeated two more times and all the supernatants/extracts were pooled. The extracts were concentrated by evaporation and the concentrated extract was stored at -80°C until analysis. The extracts were prepared in triplicates.
Determination of total polyphenol contents in daylily flowers
Total Phenolics Contents (TPC) and Total Flavonoids Content (TFC): The TPC of daylily extracts was determined using the Folin-Ciocalteu method modified for the microplate Gajula et al.19 The TPC was expressed as mg Gallic acid equivalent per gram sample (mg GAE g–1 sample). The TFC was determined via colorimetric assay20 with modifications19 and was expressed as mg catechin equivalent per gram sample (mg CE g–1 sample).
Condensed tannins and Total Monomeric Anthocyanins (TMA) contents: Condensed tannins were determined by the Vanillin-HCl/vanillin H2SO4 assay method18 and expressed in mg catechin equivalent g–1 sample (mg CE g–1 sample). The TMA of daylily flowers was determined by the pH differential method with modification21,22. Daylily extracts were diluted with two buffers, potassium chloride buffer (0.025 M, pH 1.0) and sodium acetate buffer (0.4 M, pH 4.5). Diluted samples are read against a blank at 520 and 700 nm. The TMA was expressed as mg cyanidin-3-glucoside equivalent g–1 sample (mg C3GE g–1 dwb).
Determination of antioxidant activities in daylily flowers
Trolox Equivalent Antioxidant Capacity (TEAC) assay: TEAC was determined following previous methods23. The TEAC was expressed as a Mmol Trolox g–1 sample.
Ferric Reducing Antioxidant Power (FRAP) assay: The FRAP of daylily extracts was measured using methods described by Benzie and Strain24. The FRAP activity was expressed as an mM Fe2+ g–1 sample.
DPPH radical scavenging capacity assay: The DPPH test was performed following the method of Brand-Williams et al.25, with modifications26. The IC50 was calculated and the decrease in absorbance at 515 nm was monitored at T0 and T30, T60 and T90. The DPPH scavenging ability was calculated as a percentage using the following Eq.26:
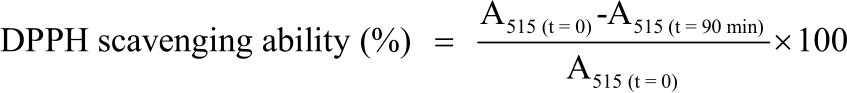 | (1) |
Metal chelating activity assay: The Metal chelating activity was measured using a modified method by Wong et al.27 Various concentrations of extracts were prepared and 180 μL of methanol (100%) and 5 μL FeCl2 (2 mM) were added and incubated for 3 min. About 10 μL of ferrozine (5 mM) was added to initiate the reaction and the mixture was incubated for 10 min at room temperature after shaking. Absorbance was read at 562 nm. The percentage of Fe2+-ferrozine complex formation was calculated as Fe2+ (%) chelating ability27.
Determination of selected polyphenols in daylily flowers via HPLC: High-performance liquid chromatography (HPLC) analysis of polyphenols in daylily flowers was carried out on a Varian Modular Analytical HPLC System. Polyphenol’s identification was determined using methods of Schutz et al.28 with modifications. Samples and standards were prepared using 80% ethanol. Mobile phase A was 0.5% (v/v) acetic acid in the water and mobile phase B was 0.5% (v/v) acetic acid in acetonitrile. Separations of polyphenols were conducted at a flow rate of 0.8 mL min–1 at 25°C. Catechin and gallic acid were eluted at 284 nm, chlorogenic acid at 320 nm and rutin at 350 nm. Spectral data was collected for each standard and sample. Polyphenols (i.e., rutin, catechin, chlorogenic acid and gallic acid) were selected based on the previous studies29-32.
Determination of in vitro cytotoxicity of Elegant Explosion (EE) extract
Cell line and culture conditions of Hep2G cells: Human hepatocellular carcinoma cell line (HepG2), (ATCC, Manassas, VA) was cultured following previous methods and conditions33. HepG2 was seeded at 104 cells cm–2 density and allowed to adhere for 48 hrs and cells were used for cytotoxicity determinations on the third day after seeding.
Determinations of cell viability of EE extract in HepG2 cells: Cell viability was determined by measuring the metabolic activity of living cells using the PrestoBlue® (Life Technologies Corporation, Grand Island, NY) assay reagent. After the seeding period specified above, HepG2 cells were incubated with different concentrations (0.2, 0.4, 0.8, 1.0, 2.0, 4.0 and 8.0 mg mL–1) of EE extract for 24 and 48 hrs. After the incubation period, the culture medium (supernatant) was removed and cell viability was determined.
Morphological evaluation of EE extract on HepG2 cells: HepG2 cells (1×104) were grown on coverslips and treated with or without daylily extracts for 48 hrs. The medium was discarded and cells were washed once with PBS. Cells were fixed with MeOH: acetic acid (3:1) and stained with NucBlue® Live reagent (Hoechst 33342) and NucGreen® Dead reagent (Life Technologies). After 15 min of incubation, the dye was removed and the cells were rinsed with PBS and imaged with the EVOS® FL Auto system (Thermo Fisher Scientific, Waltham, MA).
Determination of antioxidant/detoxification enzymes
Extraction of protein from cells: After treatment with daylily extract, cell lysate was collected from HepG2 cells following a protocol from our lab33,34. Protein in the cell lysate was determined using a BCA protein assay kit from Pierce (Rockford, IL). The cell lysate was used for enzyme analysis.
Determination of endogenous antioxidant enzymatic activity: Glutathione (GSH) was determined according to Griffith35 and expressed as μmol mg–1 protein. Superoxide Dismutase (SOD) was determined by the method outlined by Fernández-Urrusuno et al.36. Xanthine oxidase was utilized to generate a superoxide flux and samples were measured at 440 nm after the addition of xanthine oxidase. Data were expressed as units of SOD activity per mg protein (U mg–1 protein). Glutathione-S-transferase (GST) activity was based on the method of Habig et al.37 and 1-chloro-2, 4-dinitrobenzene was used as the substrate. GST was expressed as μmol mg–1 protein. All samples were analyzed in triplicates.
Statistical analysis: All experiments were repeated three independent times and results were conducted in triplicates. ANOVA was used to determine any significant differences among daylily cultivars. Data for the antioxidant assays are presented as the mean±standard deviation of the mean (SD) and those for the cytotoxicity assays are presented as the mean percentages relative to the control±SD. Significant (p<0.05 or 0.1), means were separated using Tukey’s Studentized Range Test. Statistical analysis was conducted using SAS, 9.4 tools.
RESULTS AND DISCUSSION
Total polyphenol contents in daylily flowers: The results of TPC, TFC, TMA and CTC are shown in Table 1. The TPC was significantly (p≤0.05) higher in EE (2.13 mg GAE g–1 dwb) compared to BM (1.11 mg GAE g–1 dwb). TFC showed a parallel trend between the BM and EE flowers. The TFC was approximately two times higher in EE (1.47 mg CE g–1 dwb) than in BM (0.64 mg CE g–1 dwb). While the results for TFC in EE and BM are within the range reported elsewhere for edible flowers38-41, there is high variability in TPC. The TPC reported in this study was lower compared to other studies. This variation may be due to the extraction methodology used or the types of solvent employed. Additionally, several factors such as genotype, environment, geographical location and altitude, UV exposure, season, soil, maturity stage at harvest, post-harvest conditions, etc. could all impact TPC variation. It should be noted that the daylily cultivars utilized in this study were cultivated in the AAMU greenhouse under controlled culture conditions. Flavonoids are well-known antioxidants with importance in human health. It is reported that due to their strong antioxidant capacity, most flavonoids outperform natural and well-known antioxidants including ascorbate (vitamin C) and α-tocopherol (vitamin E) because of their strong ability to donate electrons or hydrogen atoms to neutralize free radicals42. This indicates their potential as important therapeutic agents and also for improving the quality of foods43. The contents of anthocyanins in edible flowers have been well documented7,44,45. Among the most prominent anthocyanins, cyanidin-3-glucoside (C3G) is the major anthocyanin found in most flowering plants. The data of Table 1 shows the TMA content in EE was significantly (p<0.05) higher (1.73 mg C3G g–1 dwb) compared to BM (1.20 mg C3G g–1 dwb). These results are comparable to other studies46-50. Anthocyanins, besides their importance as natural colourants or pigments in foods, have various beneficial health benefits46,49,51-56. A similar trend was observed whereby CTC was significantly (p<0.05) higher in EE (0.024 mg CE g–1 dwb) than in BM (0.004 mg CE g–1 dwb) (Table 1). Condensed tannins (proanthocyanidins) have found various applications in foods. They are used as additives, they prevent oxidation in foods and enhance the shelf life of beverages including teas, fruit juices and wine57. For health purposes, condensed tannins or proanthocyanidins are noted for their antibacterial activity against urinary tract infections, for beneficial regulation of gut microbiota58,59 as well as anti-inflammatory enhancing properties57,60,61.
Antioxidant activities of daylily flowers: The result of Table 2 show the FRAP, TEAC, DPPH and Fe2+ chelating antioxidant activities of EE and BM flowers. When it comes to determining the antioxidant activity of plant extracts there’s not a standardized method for evaluation since the extracts may exhibit different antioxidant mechanisms. As such, it is important to perform more than one type of antioxidant activity measurement to take into account the various mechanisms of antioxidant action62,63.
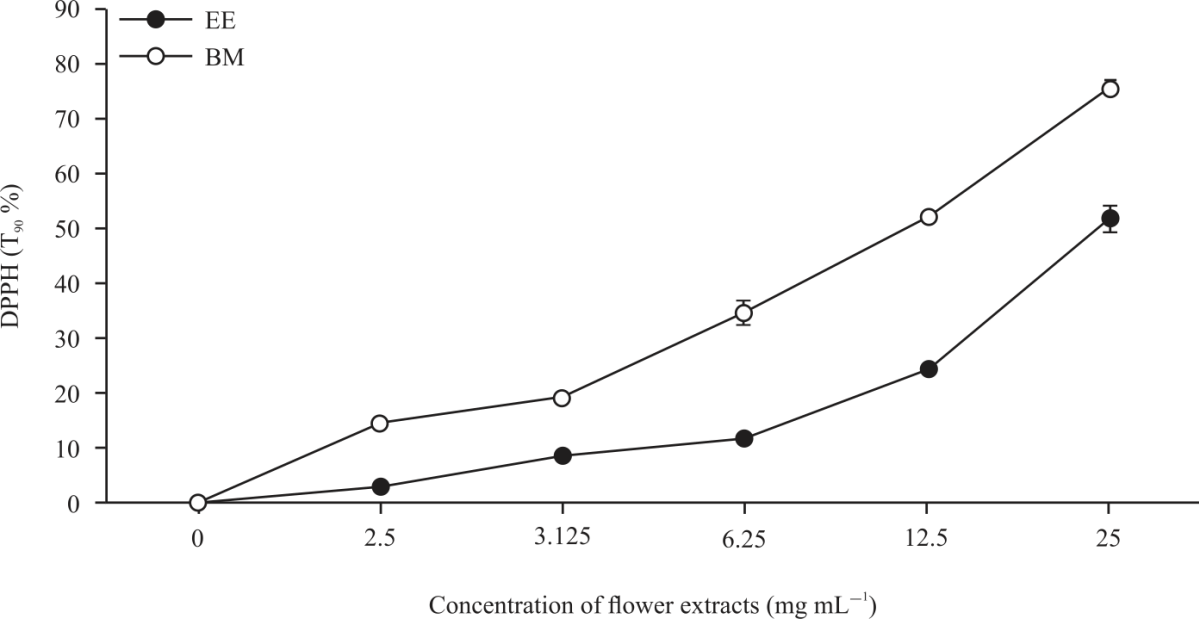 |
| Fig. 1: | DPPH (%) of daylily cultivars at T90 min Values are expressed as means±SD and were conducted in triplicates in three independent experiments. Statistical significance was determined by one-way ANOVA followed by Tukey’s range test. EE: Elegant explosion, BM: Barbara mitchell |
| Table 1: Total polyphenol contents in daylily cultivars | ||||
| Samples | TPC (mg GAE g–1 dwb) | TFC (mg CE g–1 dwb) | TMA (mg C3GE g–1 dwb) | CTC (mg CE g–1 dwb) |
| BM | 1.11±0.06b | 0.64±0.01b | 1.20±0.15b | 0.004b |
| EE | 2.13±0.01a | 1.47±0.02a | 1.73±0.08a | 0.024a |
Values (n = 3) are means±SD, means in the column (ab) letter differ (p≤0.05) using Tukey’s Studentized range test. BM: Barbara mitchell, EE: Elegant explosion, GAE: Gallic acid equivalent, DWB: Dry weight basis (dry flowers), TFC: Total flavonoid, TPC: Total phenolic, TMA: Total monomeric anthocyanin content, CTC: Condensed tannins content, CE: Catechin equivalent, GAE: Gallic acid equivalent, C3G: Cyanidin-3-glucoside equivalent | ||||
| Table 2: Antioxidant activities of daylily cultivars | ||||
| Samples | FRAP | TEAC | DPPH (IC50 mg mL–1) | Fe2+ chelating activity (IC50 mg mL–1) |
| BM | 0.97±0.04b | 4.82±0.02b | 24 | 43.08 |
| EE | 2.48±0.15a | 16.15±0.24a | 14.31 | 37.05 |
Values (n = 3) are expressed as means±SD means in column (ab) letter differ (p≤0.05) using Tukey’s Studentized range test. (1) FRAP is expressed in μmol FeSO4 per g of dry flowers, (2) TEAC is expressed in mM Trolox per g of dry flowers. BM: Barbara mitchell, EE: Elegant explosion, DPPH: 2,2-diphenyl-1-picrylhydrazyl, FRAP: Ferric reducing antioxidant power, Fe2+: Ferrous ion, TEAC: Trolox equivalent antioxidant capacity | ||||
The results in Table 2 show EE had a significantly (p<0.05) higher FRAP value (2.48 μmol FeSO4 g–1 dwb) compared to BM (0.97 μmol FeSO4 g–1 dwb). TEAC activity was higher (p<0.05) in EE (16.15 mM Trolox g–1 dwb) compared to BM (4.82 mM Trolox g–1 dwb). The DPPH assay is a rapid, direct and reproducible procedure widely used to determine the radical scavenging activity, via inhibition (%) or IC50 of plant extracts. The IC50 was 14.31 mg mL–1 (R2 = 0.94171) in EE compared to 24 mg mL–1 (R2 = 0.99363) in BM in Table 2. The data of Fig. 1 shows the DPPH expressed as percentage (%) inhibition after 90 min (T90) was higher (p<0.05) in EE (75.37%) than in BM (51.56%). These results are comparable with other studies investigating the antioxidant activities of edible flowers3,64,65. The IC50 for Fe2+ capacity was indicated at 43.08 (R2 = 0.82275) and 37.05 mg mL–1 (R2 = 0.70796) for BM and EE, respectively. In Fig. 2 the Metal chelating (Fe2+) capacity in the presence of BM and EE extracts showed a linear activity with increasing concentration. Although this study did not record the statistical correlation between polyphenol contents and antioxidant activity, many authors6,66,67 have indicated a strong linear relation between the polyphenol contents and the antioxidant activities of edible flower species.
Selected polyphenols in daylily flowers: HPLC chromatogram of selected polyphenol standards are shown in Fig. 3. The standards are chlorogenic acid (RT = 6.62 min), (+)-catechin (RT = 4.43 min), gallic acid (RT = 4.45 min) and rutin (RT = 15.04 min). The peaks were confirmed by UV/Vis spectra obtained at λmax 284 nm for (+)-catechin (Fig. 3a) and gallic acid (Fig. 3b), 320 nm for chlorogenic acid (Fig. 3c) and 350 nm for rutin (Fig. 3d). The chromatogram in Fig. 4 shows the peaks for the polyphenolic compounds identified in daylily flower extracts. In Fig. 4a, the chromatogram show (+)-catechin, chlorogenic acid and rutin were identified in EE.
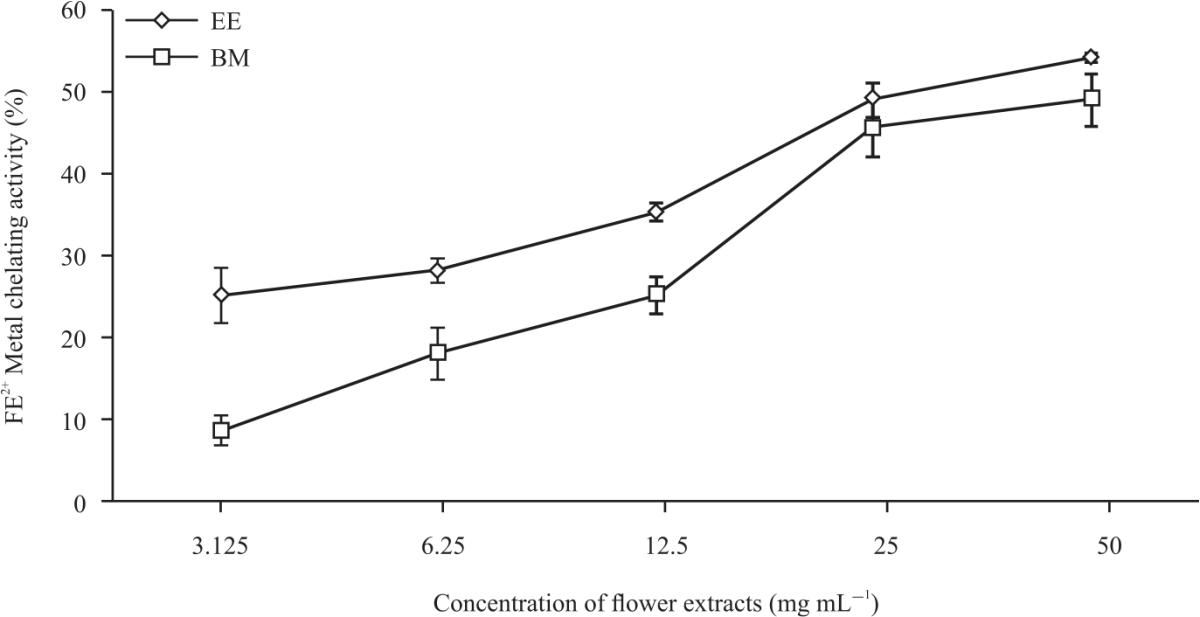 |
| Fig. 2: | Fe2+ chelating activity of daylily cultivars Values are expressed as means±SD and were conducted in triplets in three independent experiments. Statistical significance was determined by one-way ANOVA followed by Tukey’s range test. EE: Elegant explosion, BM: Barbara mitchell |
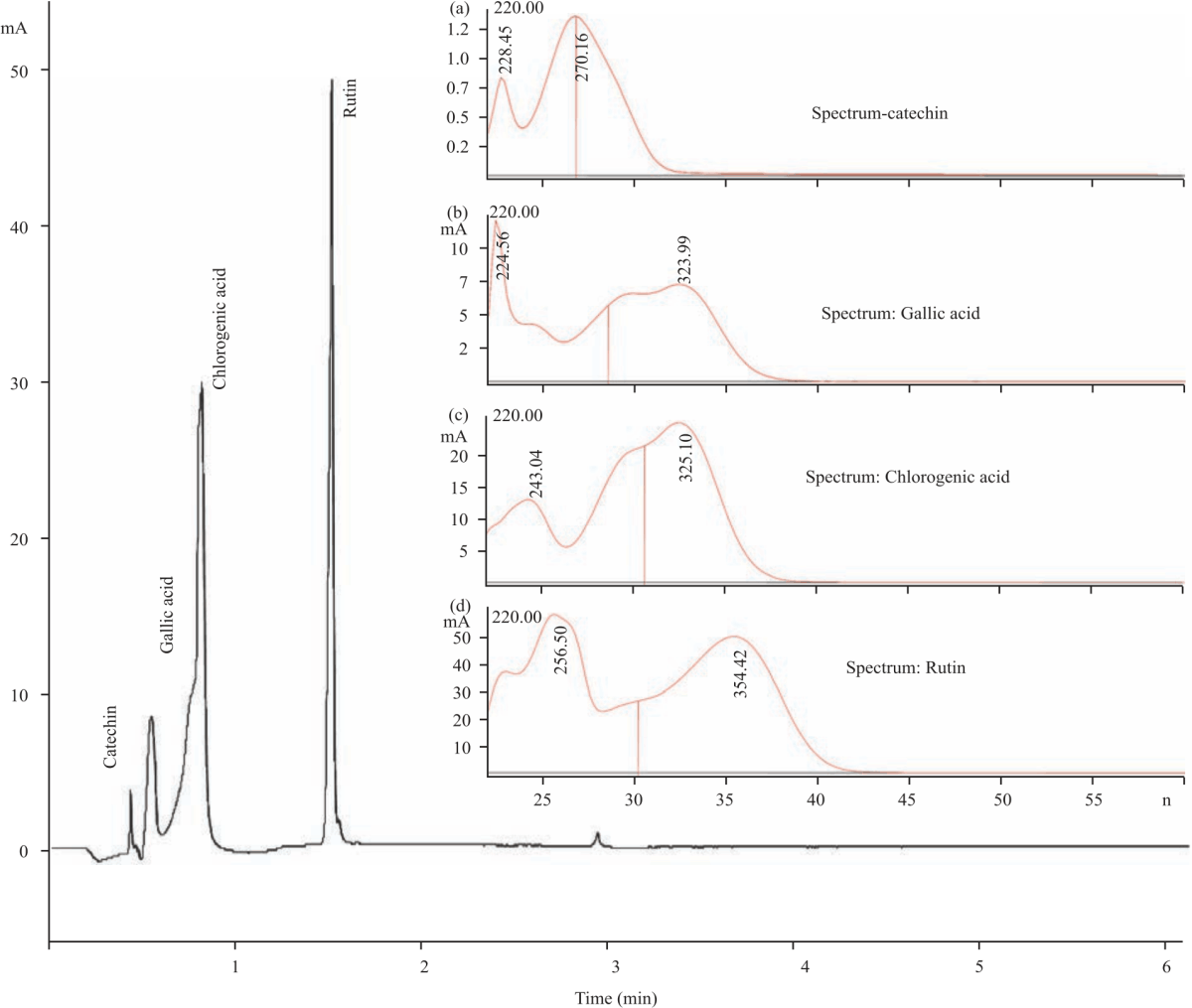 |
| Fig. 3(a-d): | HPLC Chromatographs of polyphenolic standards catechin, gallic acid, chlorogenic acid and rutin UV spectrum was recorded for (a) Catechin (284 nm), (b) Gallic Acid (284 nm), (c) Chlorogenic acid (320 nm) and (d) Rutin (350 nm) |
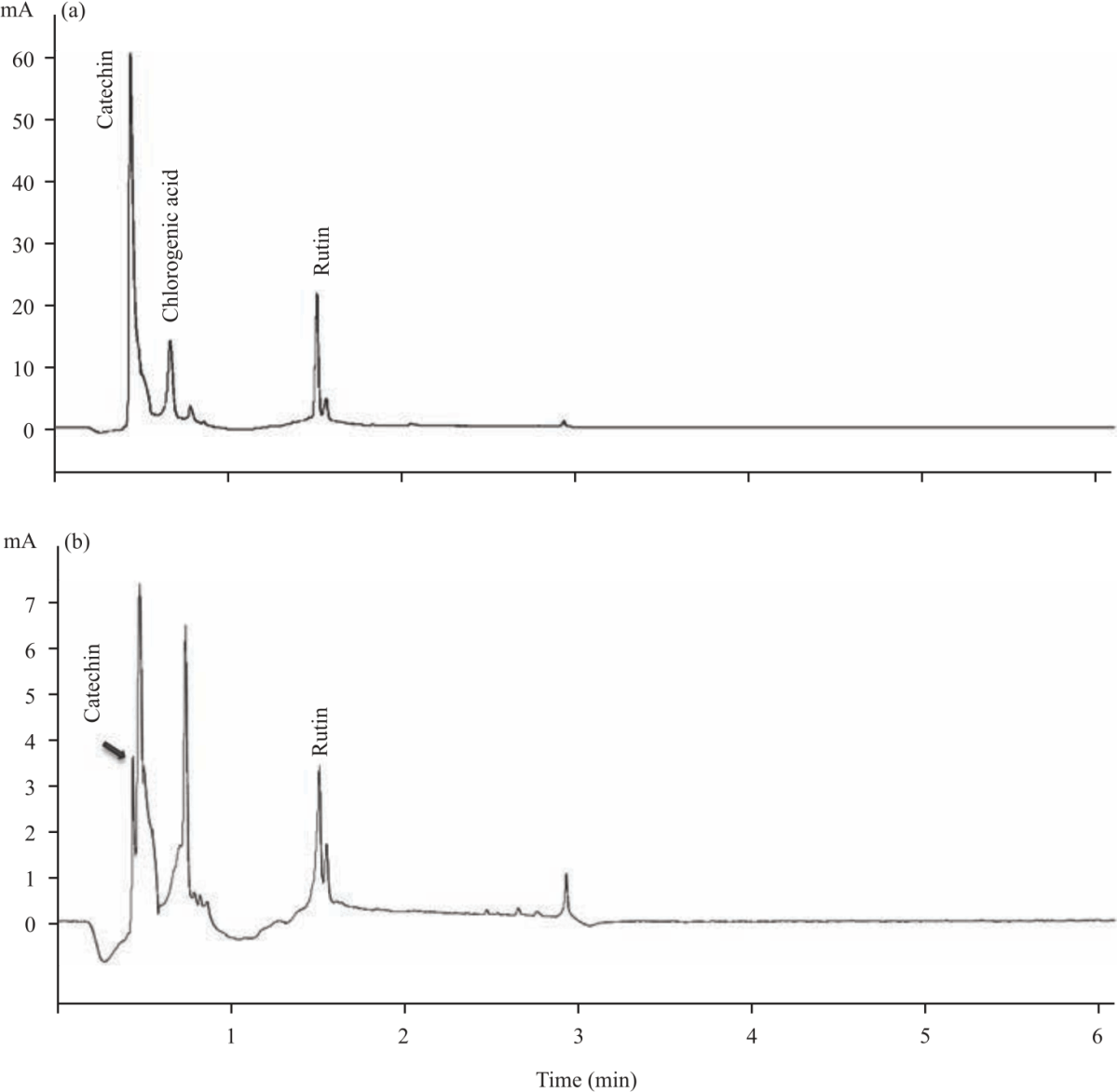 |
| Fig. 4(a-b): | HPLC Chromatograms and UV spectrums of selected polyphenols identified in daylily cultivars (a) Catechin, chlorogenic acid and rutin were identified in Elegant Explosion and (b) Catechin and rutin were identified in Barbara Mitchell |
| Table 3: Concentrations of selected polyphenol compounds (μg g–1 dwb) identified in daylily cultivars | ||||
Catechin (μg g–1) | Rutin (μg g–1) | Chlorogenic acid (μg g–1) | Gallic acid (μg g–1) | |
| BM | 13b | 11b | ND | ND |
| EE | 83a | 42a | 37 | ND |
Values (n = 3) are expressed as means±SD of three replicates. Means in column (ab) letter differ (p≤0.05) using Tukey’s Studentized range test. EE: Elegant explosion, BM: Barbara mitchell, ND: Not detected, dwb: Dry weight basis | ||||
The data of Fig. 4b shows (+)- catechin and chlorogenic were identified in BM. Surprisingly; gallic acid was below the quantification limit and was not detected in any of the flowers in our study. The data in Table 3 shows concentrations of polyphenols in EE and BM. (+)-Catechin and rutin in EE were six times and four times higher compared to BM. Chlorogenic acid was only identified in EE (37 μg g–1 dwb). The results show that (+)-catechin was the most represented polyphenol in both the daylily cultivars (83 and 13 μg g–1 for EE and BM, respectively). It is reported that in daylily flowers, (+)-catechin represents nearly 75% of all polyphenols14,68. The high (+)-catechin, chlorogenic acid and rutin concentration in EE may be responsible for the demonstrated antioxidant activity. Polyphenols such as chlorogenic acid, rutin and (+)-catechin possess several physiological activities. These compounds are shown to inhibit the proliferation of cancer cells69-72. They also have cardioprotective73,74, anti-obesity75 and anti-hypertensive properties76-79. Due to the polyphenol contents, the daylily flowers used in this study could be considered as ingredients for several types of functional food applications with health benefits or as a nutraceutical in dietary supplements. Even though there were some variations in polyphenolic composition compared to other studies80-82, these variabilities might be due to the daylily flower cultivars, experimental conditions and the type of solvent used for polyphenolic extractions.
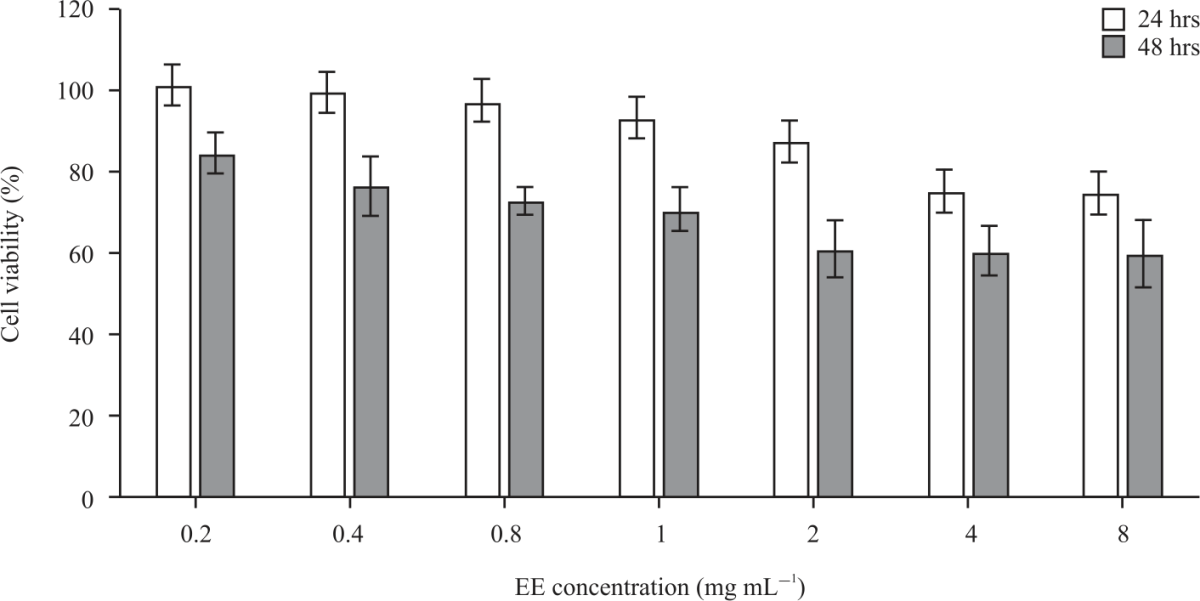 |
| Fig. 5: | Cytotoxicity of elegant explosion extracts in HepG2 cells after 24 and 48 hrs exposure Values (n = 3) are expressed as means±SD of three replicates. EE: Elegant explosion |
All the same, the polyphenol contents estimated in our study are in the range or comparable to those reported by some investigators14,30,41,66,83,84.
While gallic acid is one of the most common phenolic acids in edible plants, we did not detect this phenolic compound in our samples. Gallic acid and catechin share some structural similarities and as a result, both compounds could have co-eluted together under the HPLC conditions in this study. In any case, our results correspond with Zheng et al.85 and Barros et al.84 who reported similar findings in edible flowers from China and Brazil, respectively. Similar results were also observed with Calendula officinalis L., Chrysanthemum morifolium, Jasminum sambac (L.) Ait and other edible flowers83. Wang et al.86 reported that good separation of gallic acid and catechin is best achieved if the concentration of ethanol in the final sample solution for HPLC analysis is less than 15%. In this study, the final sample solution of ethanol was more than 15%.
Overall, the results show EE flowers contained high TPC, TFC, TMA CTC and strong antioxidant properties compared to BM flowers. As such, EE flowers were selected and used to perform in vitro toxicity experiments.
Cytotoxicity of Elegant Explosion (EE) extracts in HepG2 cells: The cytotoxicity of EE extract in Hep2G cells was achieved by the PrestoBlue® test and results are presented in Fig. 5. The PrestoBlue® assay is a simple and fast live assay to monitor cell viability and cytotoxicity87. It is a resazurin-based compound that is quickly reduced by metabolically viable cells88,89. Based on the results in Fig. 5, EE extract was non-cytotoxic activity towards HepG2 cells after 24 hrs exposure, as cytotoxicity ranged from 73-100% (8-0.2 mg mL–1). The results also show cell viability was slightly decreased with prolonged exposure (48 hrs incubation). After 48 hrs incubation cytotoxicity was 59% at the highest (8 mg mL–1) and the IC50 was recorded at 10.13 mg mL–1. The present results indicate that hydroethanolic EE extract possesses weak to moderate toxicity towards the HepG2 cell line used in this study. This assertion is based on ISO 10993-590 which specifies that cell viability above 80% is non-cytotoxic; 80-60% is weakly cytotoxic; 60-40% is moderately cytotoxic and <40% is strongly cytotoxic. The present results are comparable to the previous observations7,91,92. The effect could be attributed to the specific phenolic compounds and antioxidant activity of EE. Studies conducted by Cichewicz et al.93 and Lin et al.94 showed that polyphenols compounds anthraquinones and caffeoylquinic acid derivatives from daylily flowers showed weak cytotoxic activities and prevented oxidative stress in the HepG2 cell line. Recent research indicated caffeoylquinic acid derivatives and flavanone glycosides in Chrysanthemum morifolium displayed hepatoprotective effects in HepG2 cells95. Kwon et al.96 also reported quercetin and kaempferol in Begonia semperflorens flowers proved to be protective against oxidative stress in t BHP induced HepG2.
Morphological changes in Hep2G cells after 48h incubation: The result of Fig. 6a-d show microscopic observations in Hep2G cells exposed to EE extract after 48 hrs incubation. The results show EE extract induced high apoptosis with increasing concentration (0-4 mg mL–1). Chromatin condensation and fragmentations were observed in the HepG2 cells treated with EE extract, while the control untreated cells displayed minimal intact nuclei or apoptosis-inducing effects. Besides, microscopic observation showed cell confluence was markedly reduced with increasing concentration.
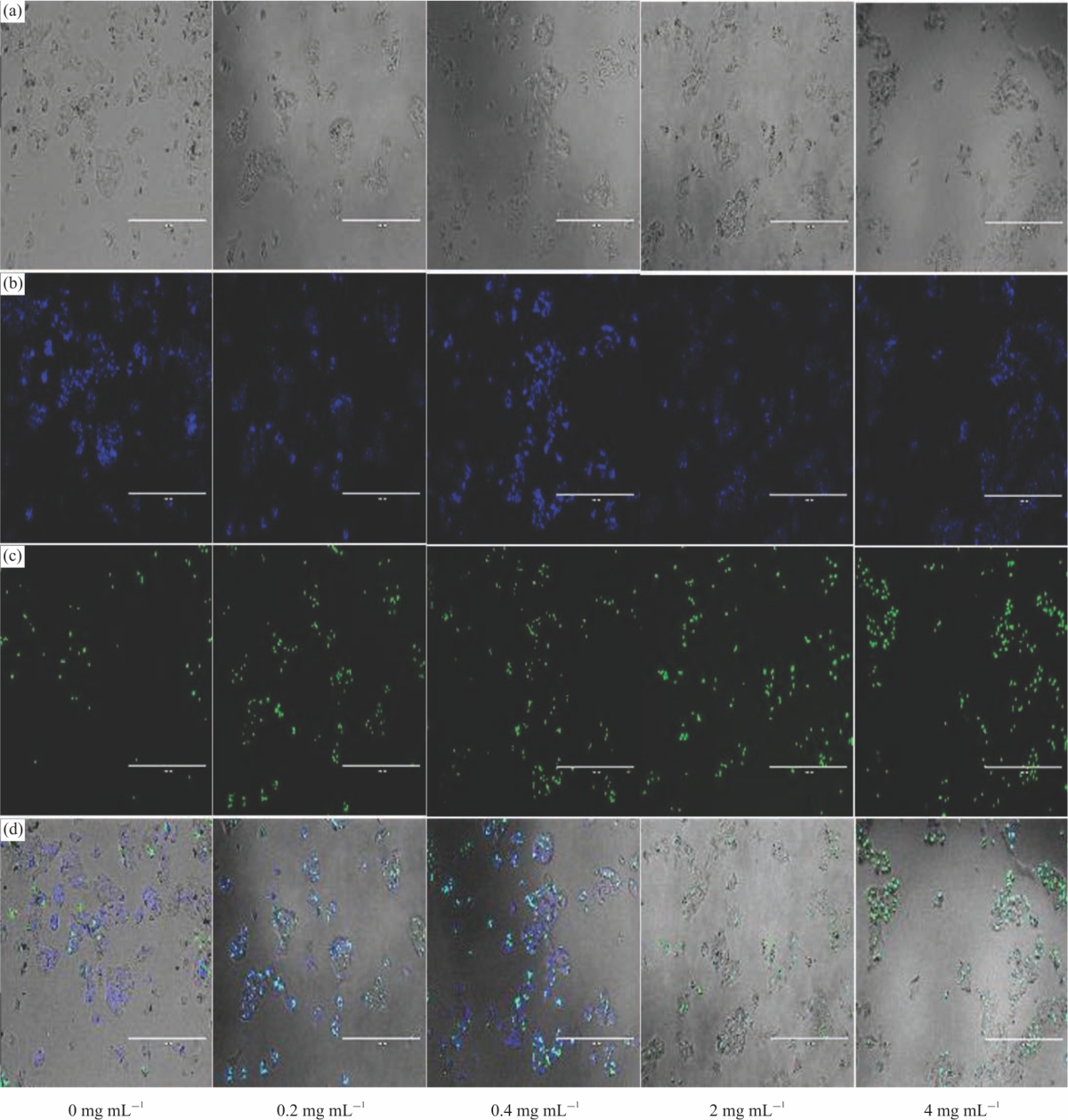 |
| Fig. 6(a-d): | Elegant explosion (EE) extract induced morphological changes in Hep2G cells following 48 hrs incubation (a) Phase-contrast photomicrographs of Hep2G cells treated with EE extract at 0 (control)-4 mg mL–1 for 48 hrs, (b-c) Cytotoxicity of EE extract analyzed by live/dead assay. NucBlue® Live (Hoechst 33342) reagent stains the nuclei of all cells while NucGreen® Dead reagent stains only the nuclei of dead cells. A higher percentage of apoptotic cells (live (blue)/dead (green)) are observed in the treated cells than in the control. Green channel signal intensity (dead cell stain) is strong in the cells treated with a high concentration of EE extract. Images are representative of three independent experiments. Cells were imaged on the EVOS® FL Auto system. Images were captured at ×10 magnification. Scale bars: 400 |
Furthermore, EE extract induced obvious morphology changes including cell shrinkage, abnormity and death as compared with control untreated cells (Fig. 6a). It is important to point out that maintenance of membrane integrity is a standard criterion for cell viability. NucBlue® Live (Hoechst 33342) reagent stains the nuclei of all cells (Fig. 6b). Live cells react with the fluorescent reactive dye only on their surface to yield weakly fluorescent cells. The nuclei of dead cells with compromised membranes react with NucGreen® Dead reagent stains only and produce brightly stained green cells (Fig. 6c). The results from cell viability analysis, i.e., presto blue are in line with cell images. The data of Fig. 6d depicts the merge or overlay of Fig. 6a-c.
| Table 4: Effects of elegant explosion daylily flowers extracts on endogenous antioxidant enzymes in HepG2 cells | |||
SOD (U mg–1 protein) | GST (μmol mg–1 protein) | GSH (μmol mg–1 protein) | |
| 24 hrs endogenous enzyme activity | |||
| Control | 1.11±0.06c | 6.36±0.31a | 0.27±0.01b |
| 0.1 | 1.4±0.04a | 5.84±0.07b | 0.40±0.02a |
| 0.2 | 1.48±0.03a | 4.60±0.13bc | 0.30±0.01a |
| 0.4 | 1.46±0.03a | 4.11±0.12bc | 0.32±0.05a |
| 0.8 | 1.47±0.02a | 4.0 ±0.16bc | 0.40±0.01a |
| 1 | 1.45±0.05a | 3.57±0.15c | 0.34±0.02a |
| 2 | 1.43±0.02a | 3.0 ±0.11c | 0.35±0.02a |
| 4 | 1.32±0.07b | 3.54±0.23c | 0.37±0.03a |
| 48 hrs endogenous enzyme activity | |||
| Control | 0.88±0.07c | 4.08±0.53ab | 0.14±0.01b |
| 0.1 | 1.15±0.01b | 4.68±1.05a | 0.40 ±0.01a |
| 0.2 | 1.32±0.03a | 3.87±0.38ab | 0.38 ±0.01a |
| 0.4 | 1.28±0.04a | 3.43±0.24b | 0.35 ±0.01a |
| 0.8 | 1.22±0.06a | 3.11±0.18b | 0.31±0.01a |
| 1 | 1.17±0.05b | 3.01±0.05b | 0.41±0.01a |
| 2 | 1.13±0.03b | 2.83±0.15b | 0.48 ±0.01a |
| 4 | 1.02±0.06bc | 2.97±0.12b | 0.46 ±0.01a |
Values are expressed as means±SD and were conducted in triplicates. abcLetters demonstrate the significant difference (p≤0.05), comparing the concentrations of the assay at 24 and 48 hrs. Statistical significance was determined by one-way ANOVA followed by Tukey’s range test. SOD: Superoxide dismutase, GSH: Glutathione, GST: Glutathione-S-Transferase | |||
Effect of EE extract on redox biomarkers in HepG2 cells: The result of Table 4 shows the effect of EE extract on endogenous enzyme activity. SOD activity (U mg–1 protein) was increased (p<0.05) in HepG2 cells exposed to EE extract (1.32-1.48 U mg–1 protein after 24 hrs incubation and 1.02-1.32 U mg–1 protein after 48 hrs incubation) compared to control untreated cells (1.11 U mg–1 protein and 0.88 U mg–1 protein, respectively for 24 and 48 hrs). The results show SOD activity was significantly (p<0.05) different after 24 and 48 hrs incubation. While the present data show a high SOD activity after treatment with EE extract, there was no linearity regarding concentrations. Other studies have indicated that SOD activity increase when treated with polyphenols97,98. SOD is a family of antioxidant enzymes that convert harmful superoxide radicals into H2O2, which is subsequently metabolized to water and oxygen by catalase and glutathione peroxidase. The present results could be due to the high antioxidant activity of the EE extract directly retarding the superoxide radical’s formation. It is also possible that the EE extract activated the SOD enzyme that catalyzes the conversion of superoxide radicals into hydrogen peroxide and oxygen for subsequent detoxification to water and oxygen.
GST activity (μmol mg–1 protein) decreased (p<0.05) in HepG2 cells exposed to EE extract for 24 hrs compared with control shown in Table 4. However, the exposure of cells for 48 hrs produced no significant changes compared to the control. The results are in line with others who reported decreased GST activity in HepG2 cells after acute incubation with basil and ginger extracts33,97 and grape pomace extract99. Also, previous studies have detected no change in GST activity after prolonged incubation with polyphenolic compounds in HepG2 cells100-102.
The results of the GSH level shown in Table 4 followed a similar pattern observed for SOD activity. The results demonstrated no significant difference in GSH levels as EE extract concentration increased. A study by Franco and Cidlowski103 indicated that GSH depletion is a common feature of apoptotic cell death. GSH participates in the disposal of potentially harmful electrophiles and protects cells from the damaging effects of free radicals104,105. Therefore, its depletion renders cells particularly vulnerable to oxidative stress. Since these results indicated no significant associations between cell viability and cellular GSH level, this phenomenon could be due to the tendency for cultured cells to increase protection as the concentration of extracts is increased106. Some studies utilizing high concentrations of plant extracts have reported increased GSH levels97. Others have suggested that GSH synthesis could be up-regulated during oxidative stress by phenolic compounds if their concentration does not compromise cell viability107. This might finally contribute to the paradox that is associated with the protective mechanisms of GSH.
CONCLUSION
A preliminary study of herbal extracts from daylily flower cultivars Elegant Explosion (EE) and Barbara Mitchell (BM) were evaluated for their polyphenol content and antioxidant activity. Also, the in vitro assessment of the safety (cytotoxicity/cell viability) and bioefficacy (effect on endogenous antioxidant enzymes) was evaluated for EE extract. According to results EE extracts exhibited higher TPC, TFC, TMA, polyphenol contents and strong free radical scavenging inhibitory activities. Furthermore, EE extracts induced weak to moderate cytotoxicity.
SIGNIFICANCE STATEMENT
The extent of this study is to utilize daylilies for use as functional food ingredients. The results generated from this study will help in gathering baseline information on the potential of daylily flowers to be utilized in the preparation of nutraceutical, functional food products and dietary supplements.
ACKNOWLEDGMENT
This study was funded by the USDA capacity Building grant No. 11343134, Evans Allen project No: ALAX-012-0812 and Alabama A and M University Agriculture Research Station, Normal AL 35762.
REFERENCES
- Friedman, H., O. Agami, Y. Vinokur, S. Droby and L. Cohen et al., 2010. Characterization of yield, sensitivity to Botrytis cinerea and antioxidant content of several rose species suitable for edible flowers. Sci. Hortic., 123: 395-401.
CrossRefDirect Link - Rop, O., J. Mlcek, T. Jurikova, J. Neugebauerova and J. Vabkova, 2012. Edible flowers-A new promising source of mineral elements in human nutrition. Molecules, 17: 6672-6683.
CrossRefDirect Link - Fernandes, L., S. Casal, J.A. Pereira, J.A. Saraiva and E. Ramalhosa, 2017. Edible flowers: A review of the nutritional, antioxidant, antimicrobial properties and effects on human health. J. Food Compos. Anal., 60: 38-50.
CrossRefDirect Link - Newman, M. and C.L. Kirker, 2016. Edible Flowers: A Global History. 1st Edn., Reaktion Books Ltd., London, pp: 19-21.
Direct Link - Stefaniak, A. and M.E. Grzeszczuk, 2018. Nutritional and biological value of five edible flower species. Not. Bot. Horti. Agrobo. Cluj-Napoca, 47: 128-134.
CrossRefDirect Link - Navarro-González, I., R. González-Barrio, V. García-Valverde, A.B. Bautista-Ortín and M.J. Periago, 2015. Nutritional composition and antioxidant capacity in edible flowers: Characterisation of phenolic compounds by HPLC-DAD-ESI/MSn. Int. J. Mole. Sci., 16: 805-822.
CrossRefDirect Link - Pires, T.C.S.P., M.I. Dias, L. Barros, R.C. Calhelha and M.J. Alves et al., 2018. Edible flowers as sources of phenolic compounds with bioactive potential. Food Res. Int., 105: 580-588.
CrossRefDirect Link - Pinedo-Espinoza, J.M., J. Gutiérrez-Tlahque, Y.O. Santiago-Saenz, C.L. Aguirre-Mancilla and M. Reyes-Fuentes et al., 2020. Nutritional composition, bioactive compounds and antioxidant activity of wild edible flowers consumed in semiarid regions of mexico. Plant Foods Hum. Nutr., 75: 413-419.
CrossRefDirect Link - Cunningham, E., 2015. What nutritional contribution do edible flowers make? J. Acad. Nut. Diet., Vol. 115.
CrossRefDirect Link - Loizzo, M.R., A. Pugliese, M. Bonesi, M.C. Tenuta, F. Menichini, J. Xiao and R. Tundis, 2016. Edible flowers: A rich source of phytochemicals with antioxidant and hypoglycemic properties. J. Agric. Food Chem., 64: 2467-2474.
CrossRefDirect Link - Shi, J., J. Gong, J. Liu, X. Wu and Y. Zhang, 2009. Antioxidant capacity of extract from edible flowers of Prunus mume in China and its active components. LWT-Food Sci. Technol., 42: 477-482.
CrossRefDirect Link - Kao, F.J., W.D. Chiang and H.M. Liu, 2015. Inhibitory effect of daylily buds at various stages of maturity on nitric oxide production and the involved phenolic compounds. LWT-Food Sci. Technol., 61: 130-137.
CrossRefDirect Link - Liu, X.L., L. Luo, B.B. Liu, J. Li, D. Geng, Q. Liu and L.T. Yi, 2014. Ethanol extracts from Hemerocallis citrina attenuate the upregulation of proinflammatory cytokines and indoleamine 2,3-dioxygenase in rats. J. Ethnopharmacol., 153: 484-490.
CrossRefDirect Link - Fu, M., Z. He, Y. Zhao, J. Yang and L. Mao, 2009. Antioxidant properties and involved compounds of daylily flowers in relation to maturity. Food Chem., 114: 1192-1197.
CrossRefDirect Link - Que, F., L.C. Mao and X.J. Zheng, 2007. In vitro and vivo antioxidant activities of daylily flowers and the involvement of phenolic compounds. Asia Pac. J. Clin. Nutr., 16: 196-203.
CrossRefDirect Link - Mueller, D.S., J.L. Williams-Woodward and J.W. Buck, 2003. Resistance of daylily cultivars to the daylily rust pathogen, Puccinia hemerocallidis. HortScience, 38: 1137-1140.
CrossRefDirect Link - Rodriguez-Enriquez, M.J. and R.T. Grant-Downton, 2013. A new day dawning: Hemerocallis (daylily) as a future model organism. Ann. Bot. Plants, Vol. 5.
CrossRefDirect Link - Granato, D., J.S. Santos, L.G. Maciel and D.S. Nunes, 2016. Chemical perspective and criticism on selected analytical methods used to estimate the total content of phenolic compounds in food matrices. TrAC Trends Anal. Chem., 80: 266-279.
CrossRefDirect Link - Gajula, D., M. Verghese, J. Boateng, L.T. Walker, L. Shackelford, S.R. Mentreddy and S. Cedric, 2009. Determination of total phenolics, flavonoids and antioxidant and chemopreventive potential of basil (Ocimum basilicum L. and Ocimum tenuiflorum L.). Int. J. Cancer Res., 5: 130-143.
CrossRefDirect Link - Kim, D.O., O.K. Chun, Y.J. Kim, H.Y. Moon and C.Y. Lee, 2003. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem., 51: 6509-6515.
CrossRefDirect Link - Luna-Vital, D., Q. Li, L. West, M. West and E.G. de Mejia, 2017. Anthocyanin condensed forms do not affect color or chemical stability of purple corn pericarp extracts stored under different pHs. Food Chem., 232: 639-647.
CrossRefDirect Link - Lee, J., R.W. Durst and R.E. Wrolstad, 2005. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int., 88: 1269-1278.
CrossRefDirect Link - Montoro, P., A. Braca, C. Pizza and N. De Tommasi, 2005. Structure-oxidant activity relationship of flavonoids isolated from different plant species. Food Chem., 92: 349-355.
CrossRef - Benzie, I.F.F. and J.J. Strain, 1996. The ferric reducing ability of plasma (FRAP) as a measure of "antioxidant power": The FRAP assay. Anal. Biochem., 239: 70-76.
CrossRefPubMedDirect Link - Brand-Williams, W., M.E. Cuvelier and C. Berset, 1995. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol., 28: 25-30.
CrossRefDirect Link - Muñoz-Espada, A.C., K.V. Wood, B. Bordelon and B.A. Watkins, 2004. Anthocyanin quantification and radical scavenging capacity of concord, norton and marechal foch grapes and wines. J. Agric. Food Chem., 52: 6779-6786.
CrossRefDirect Link - Wong, F.C., A.L. Yong, E.P.S. Ting, S.C. Khoo, H.C. Ong and T.T. Chai, 2014. Antioxidant, metal chelating, anti-glucosidase activities and phytochemical analysis of selected tropical medicinal plants. Iran. J. Pharm. Res., 13: 1409-1415.
Direct Link - Schutz, K., D. Kammerer, R. Carle and A. Schieber, 2004. Identification and quantification of caffeoylquinic acids and flavonoids from artichoke (Cynara scolymus L.) heads, juice and pomace by HPLC-DAD-ESI/MSn. J. Agric. Food Chem., 52: 4090-4096.
CrossRefPubMedDirect Link - Kaisoon, O., S. Siriamornpun, N. Weerapreeyakul and N. Meeso, 2011. Phenolic compounds and antioxidant activities of edible flowers from Thailand. J. Funct. Foods, 3: 88-99.
CrossRefDirect Link - Kaisoon, O., I. Konczak and S. Siriamornpun, 2012. Potential health enhancing properties of edible flowers from Thailand. Food Res. Int., 46: 563-571.
CrossRef - Xiong, L., J. Yang, Y. Jiang, B. Lu and Y. Hu et al., 2014. Phenolic compounds and antioxidant capacities of 10 common edible flowers from China. J. Food Sci., 79: C517-C525.
CrossRefDirect Link - Liu, W., Y. Zhao, J. Sun, G. Li, Y. Shan and P. Chen, 2017. Study the effects of drying processes on chemical compositions in daylily flowers using flow injection mass spectrometric fingerprinting method and chemometrics. Food Res. Int., 102: 493-503.
CrossRefDirect Link - Yang, M., R. Hardin, S. Ogutu, M. Verghese and J. Boateng, 2016. Preliminary analysis of in vitro digestion and bioactivity assessment of basil and ginger in human liver cancer cell line. J. Biol. Sci., 16: 202-214.
CrossRefDirect Link - Boateng, J. and M. Verghese, 2012. Protective effects of the phenolic extracts of fruits against oxidative stress in human lung cells. Int. J. Pharmacol., 8: 152-160.
CrossRefDirect Link - Griffith, O.W. and A. Meister, 1985. Origin and turnover of mitochondrial glutathione. Proc. Nati. Acad. Sci. USA, 82: 4668-4672.
CrossRefDirect Link - Fernandez-Urrusuno, R., E. Fattal, J. Feger, P. Couvreur and P. Therond, 1997. Evaluation of hepatic antioxidant systems after intravenous administration of polymeric nanoparticles. Biomaterials, 18: 511-517.
CrossRefDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Barriada-Bernal, L.G., N. Almaraz-Abarca, E.A. Delgado-Alvarado, T. Gallardo-Velázquez and J.A. Ávila-Reyes et al., 2014. Flavonoid composition and antioxidant capacity of the edible flowers of agave durangensis (Agavaceae). CyTA J. Food, 12: 105-114.
CrossRefDirect Link - Tian, H., F.F. Yang, C.Y. Liu, X.M. Liu and R.L. Pan et al., 2017. Effects of phenolic constituents of daylily flowers on corticosterone- and glutamate-treated PC12 cells. BMC Comp. Altern. Med., Vol. 17.
CrossRefDirect Link - Wang, Y., T. Xu, B. Fan, L. Zhang, C. Lu, D. Wang and F. Wang, 2018. Advances in researches on chemical composition and functions of Hemerocallis plants. Med. Plant, 9: 16-21.
CrossRefDirect Link - Wu, W.T., M.C. Mong, Y.C. Yang, Z.H. Wang and M.C. Yin, 2018. Aqueous and ethanol extracts of daylily flower (Hemerocallis fulva L.) protect HUVE cells against high glucose. J. Food Sci., 83: 1463-1469.
CrossRefDirect Link - Hernandez, I., L. Alegre, F.V. Breusegem and S. Munne-Bosch, 2009. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci., 14: 125-132.
CrossRefDirect Link - Pérez-Jiménez, J. and F. Saura-Calixto, 2018. Fruit peels as sources of non-extractable polyphenols or macromolecular antioxidants: Analysis and nutritional implications. Food Res. Int., 111: 148-152.
CrossRefDirect Link - Landi, M., B. Ruffoni, L. Guidi, M. Savona and D. Salvi, 2015. Cold storage does not affect ascorbic acid and polyphenolic content of edible flowers of a new hybrid sage. Agrochimica, 59: 348-357.
CrossRefDirect Link - Yang, H. and Y. Shin, 2017. Antioxidant compounds and activities of edible roses (Rosa hybrida spp.) from different cultivars grown in Korea. Appl. Biol. Chem., 60: 129-136.
CrossRefDirect Link - Cendrowski, A., I. Ścibisz, M. Mitek, M. Kieliszek and J. Kolniak-Ostek, 2017. Profile of the phenolic compounds of Rosa rugosa petals. J. Food Qual., Vol. 2017.
CrossRefDirect Link - Chensom, S., Y. Shimada, H. Nakayama, K. Yoshida and T. Kondo et al., 2020. Determination of anthocyanins and antioxidants in ‘titanbicus’ edible flowers in vitro and in vivo. Plant Foods Hum. Nutr., 75: 265-271.
CrossRefDirect Link - Chaiyasut, C., B.S. Sivamaruthi, N. Pengkumsri, S. Sirilun, S. Peerajan, K. Chaiyasut and P. Kesika, 2016. Anthocyanin profile and its antioxidant activity of widely used fruits, vegetables and flowers in thailand. Asian J. Pharmaceut. Clin. Res., 9: 218-224.
Direct Link - Grajeda-Iglesias, C., M.C. Figueroa-Espinoza, N. Barouh, B. Baréa, A. Fernandes, V. de Freitas and E. Salas, 2016. Isolation and characterization of anthocyanins from Hibiscus sabdariffa flowers. J. Nat. Prod., 79: 1709-1718.
CrossRefDirect Link - Benvenuti, S., E. Bortolotti and R. Maggini, 2016. Antioxidant power, anthocyanin content and organoleptic performance of edible flowers. Sci. Hort., 199: 170-177.
CrossRefDirect Link - Wang, L.S. and G.D. Stoner, 2008. Anthocyanins and their role in cancer prevention. Cancer Lett., 269: 281-290.
CrossRefDirect Link - He, J. and M.M. Giusti, 2010. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol., 1: 163-187.
CrossRefDirect Link - Azzini, E., J. Giacometti and G.L. Russo, 2017. Antiobesity effects of anthocyanins in preclinical and clinical studies. Oxid. Med. Cell Longev., Vol. 2017.
CrossRefDirect Link - Tsang, C., N.F. Smail, S. Almoosawi, G.J.M. McDougall and E.A.S. Al-Dujaili, 2018. Antioxidant rich potato improves arterial stiffness in healthy adults. Plant Foods Hum. Nutr., 73: 203-208.
CrossRefDirect Link - Hester, S.N., A. Mastaloudis, R. Gray, J.M. Antony, M. Evans and S.M. Wood, 2018. Efficacy of an anthocyanin and prebiotic blend on intestinal environment in obese male and female subjects. J. Nutr. Metab., Vol. 2018.
CrossRefDirect Link - Vugic, L., N. Colson, E. Nikbakht, A. Gaiz, O.J. Holland, A.R. Kundur and I. Singh, 2020. Anthocyanin supplementation inhibits secretion of pro-inflammatory cytokines in overweight and obese individuals. J. Funct. Foods, Vol. 64.
CrossRefDirect Link - Rauf, A., M. Imran, T. Abu-Izneid, Iahtisham-Ul-Haq and S. Patel et al., 2019. Proanthocyanidins: A comprehensive review. Biomed. Pharmacother., Vol. 116.
CrossRefDirect Link - Tao, W., Y. Zhang, X. Shen, Y. Cao, J. Shi, X. Ye and S. Chen, 2019. Rethinking the mechanism of the health benefits of proanthocyanidins: Absorption, metabolism and interaction with gut microbiota. Compr. Rev. Food Sci. Food Saf., 18: 971-985.
CrossRefDirect Link - Unusan, N., 2020. Proanthocyanidins in grape seeds: An updated review of their health benefits and potential uses in the food industry. J. Funct. Foods, Vol. 67.
CrossRefDirect Link - Coleman, S.L. and O.M. Shaw, 2017. Progress in the understanding of the pathology of allergic asthma and the potential of fruit proanthocyanidins as modulators of airway inflammation. Food Funct., 8: 4315-4324.
CrossRefDirect Link - Lai, R., D. Xian, X. Xiong, L. Yang, J. Song and J. Zhong, 2018. Proanthocyanidins: Novel treatment for psoriasis that reduces oxidative stress and modulates Th17 and Treg cells. Redox Rep., 23: 130-135.
CrossRefDirect Link - González-Barrio, R., M.J. Periago, C. Luna-Recio, F.J. Garcia-Alonso and I. Navarro-González, 2018. Chemical composition of the edible flowers, pansy (Viola wittrockiana) and snapdragon (Antirrhinum majus) as new sources of bioactive compounds. Food Chem., 252: 373-380.
CrossRefDirect Link - Frankle, E.N. and A.S. Meyer, 2000. The problems of using one-dimensional methods to evaluate multifunctional food and biological antioxidants. J. Sci. Food Agric., 80: 1925-1941.
CrossRef - Teerarak, M., K. Changsawake, K. Pilasombut and C. Laosinwattana, 2018. Antioxidant activities and heat stability of edible flowers of Telosma minor and Sesbania javanica. J. Herbs Spices Med. Plants, 24: 87-98.
CrossRefDirect Link - Zheng, J., X. Yu, M. Maninder and B. Xu, 2018. Total phenolics and antioxidants profiles of commonly consumed edible flowers in China. Int. J. Food Prop., 21: 1524-1540.
CrossRefDirect Link - Li, A.N., S. Li, H.B. Li, D.P. Xu, X.R. Xu and F. Chen, 2014. Total phenolic contents and antioxidant capacities of 51 edible and wild flowers. J. Funct. Foods, 6: 319-330.
CrossRefDirect Link - Garzón, G.A., D.C. Manns, K. Riedl, S.J. Schwartz and O. Padilla-Zakour, 2015. Identification of phenolic compounds in petals of nasturtium flowers (Tropaeolum majus) by high-performance liquid chromatography coupled to mass spectrometry and determination of oxygen radical absorbance capacity (ORAC). J. Agric. Food Chem., 63: 1803-1811.
CrossRefDirect Link - Mlcek, J. and O. Rop, 2011. Fresh edible flowers of ornamental plants-A new source of nutraceutical foods. Trends Food Sci. Technol., 22: 561-569.
CrossRefDirect Link - Mayr, C., A. Wagner, D. Neureiter, M. Pichler and M. Jakab et al., 2015. The green tea catechin epigallocatechin gallate induces cell cycle arrest and shows potential synergism with cisplatin in biliary tract cancer cells. BMC Comp. Altern. Med., Vol. 15.
CrossRefDirect Link - Bimonte, S., M. Cascella, M. Leongito, R. Palaia, D. Caliendo, F. Izzo and A. Cuomo, 2017. An overview of pre-clinical studies on the effects of (-)-epigallocatechin-3-gallate, a catechin found in green tea, in treatment of pancreatic cancer. Recenti progressi Med., 108: 282-287.
CrossRefDirect Link - Hou, N., N. Liu, J. Han, Y. Yan and J. Li, 2017. Chlorogenic acid induces reactive oxygen species generation and inhibits the viability of human colon cancer cells. Anti-cancer Drugs, 28: 59-65.
CrossRefDirect Link - Yan, Y., N. Liu, N. Hou, L. Dong and J. Li, 2017. Chlorogenic acid inhibits hepatocellular carcinoma in vitro and in vivo. J. Nut. Biochem., 46: 68-73.
CrossRefDirect Link - Mills, C.E., A. Flury, C. Marmet, L. Poquet and S.F. Rimoldi et al., 2017. Mediation of coffee-induced improvements in human vascular function by chlorogenic acids and its metabolites: Two randomized, controlled, crossover intervention trials. Clin. Nut., 36: 1520-1529.
CrossRefDirect Link - Santana-Gálvez, J., L. Cisneros-Zevallos and D. Jacobo-Velázquez, 2017. Chlorogenic acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules, Vol. 22.
CrossRefDirect Link - Cai, E.P. and J.K. Lin, 2009. Epigallocatechin gallate (EGCG) and rutin suppress the glucotoxicity through activating IRS2 and AMPK signaling in rat pancreatic β cells. J. Agric. Food Chem., 57: 9817-9827.
CrossRefDirect Link - Khalesi, S., J. Sun, N. Buys, A. Jamshidi, E. Nikbakht-Nasrabadi and H. Khosravi-Boroujeni, 2014. Green tea catechins and blood pressure: A systematic review and meta-analysis of randomised controlled trials. Eur. J. Nutr., 53: 1299-1311.
CrossRefDirect Link - Onakpoya, I.J., E.A. Spencer, M.J. Thompson and C.J. Heneghan, 2015. The effect of chlorogenic acid on blood pressure: A systematic review and meta-analysis of randomized clinical trials. J. Hum. Hypertens., 29: 77-81.
CrossRefDirect Link - Ward, N.C., J.M. Hodgson, R.J. Woodman, D. Zimmermann and L. Poquet et al., 2016. Acute effects of chlorogenic acids on endothelial function and blood pressure in healthy men and women. Food Funct., 7: 2197-2203.
CrossRefDirect Link - Szewczyk, K., M. Miazga-Karska, W. Pietrzak, Ł. Komsta, B. Krzemińska and A. Grzywa-Celińska 2020. Phenolic composition and skin-related properties of the aerial parts extract of different Hemerocallis cultivars. Antioxidants, Vol. 9.
CrossRefDirect Link - Lin, S.H., H.C. Chang, P.J. Chen, C.L. Hsieh, K.P. Su and L.Y. Sheen, 2013. The antidepressant-like effect of ethanol extract of daylily flowers (金針花 Jīn Zhēn Huā) in rats. J. Traditional Complementary Med., 3: 53-61.
CrossRefDirect Link - Joshi, R. and A. Gulati, 2011. Biochemical attributes of tea flowers (Camellia sinensis) at different developmental stages in the Kangra region of India. Sci. Hortic., 130: 266-274.
CrossRefDirect Link - Chen, G.L., S.G. Chen, Y.Q. Xie, F. Chen, Y.Y. Zhao, C.X. Luo and Y.Q. Gao, 2015. Total phenolic, flavonoid and antioxidant activity of 23 edible flowers subjected to in vitro digestion. J. Funct. Foods, 17: 243-259.
CrossRefDirect Link - Barros, R.G.C., J.K.S. Andrade, U.C. Pereira, C.S. de Oliveira and Y.R.R.S. Rezende et al., 2020. Phytochemicals screening, antioxidant capacity and chemometric characterization of four edible flowers from Brazil. Food Res. Int., Vol. 130.
CrossRefDirect Link - Zheng, J., M. Meenu and B. Xu, 2019. A systematic investigation on free phenolic acids and flavonoids profiles of commonly consumed edible flowers in China. J. Pharm. Biomed. Anal., 172: 268-277.
CrossRefDirect Link - Wang, H., K. Helliwell and X. You, 2000. Isocratic elution system for the determination of catechins, caffeine and gallic acid in green tea using HPLC. Food Chem., 68: 115-121.
CrossRefDirect Link - Emter, R. and A. Natsch, 2015. A fast resazurin-based live viability assay is equivalent to the MTT-test in the KeratinoSens assay. Toxicol. in vitro., 29: 688-693.
CrossRefDirect Link - O'Brien, J., I. Wilson, T. Orton and F. Pognan, 2000. Investigation of the alamar blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem., 267: 5421-5426.
Direct Link - Boncler, M., M. Rozalski, U. Krajewska, A. Podsedek and C. Watala, 2014. Comparison of PrestoBlue and MTT assays of cellular viability in the assessment of anti-proliferative effects of plant extracts on human endothelial cells. J. Pharmacol. Toxicol. Meth., 69: 9-16.
CrossRefDirect Link - Wallin, R.F. and E.F. Arscott, 1998. A practical guide to ISO 10993-5: Cytotoxicity. Med. Dev. Diag. Ind., 20: 96-98.
Direct Link - Sagdic, O., L. Ekici, I. Ozturk, T. Tekinay and B. Polat et al., 2013. Cytotoxic and bioactive properties of different color tulip flowers and degradation kinetic of tulip flower anthocyanins. Food Chem. Toxicol., 58: 432-439.
CrossRefDirect Link - López-García, J., Z. Kuceková, P. Humpolíček, J. Mlček and P. Sáha, 2013. Polyphenolic extracts of edible flowers incorporated onto atelocollagen matrices and their effect on cell viability. Molecules, 18: 13435-13445.
CrossRefDirect Link - Cichewicz, R.H., Y. Zhang, N.P. Seeram and M.G. Nair, 2004. Inhibition of human tumor cell proliferation by novel anthraquinones from daylilies. Life Sci., 74: 1791-1799.
CrossRefDirect Link - Lin, Y.L., C.K. Lu, Y.J. Huang and H.J. Chen, 2011. Antioxidative caffeoylquinic acids and flavonoids from Hemerocallis fulva flowers. J. Agric. Food Chem., 59: 8789-8795.
CrossRefDirect Link - Yang, P.F., Y.N. Yang, C.Y. He, Z.F. Chen and Q.S. Yuan et al., 2019. New caffeoylquinic acid derivatives and flavanone glycoside from the flowers of Chrysanthemum morifolium and their bioactivities. Molecules, Vol. 24.
CrossRefDirect Link - Kwon, J.H., H.J. Oh, D.S. Lee, S.J. In and K.H. Seo et al., 2019. Pharmacological activity and quantitative analysis of flavonoids isolated from the flowers of Begonia semperflorens Link et Otto. Appl. Biol. Chem., Vol. 62.
CrossRefDirect Link - Offei-Oknye, R., J.L. Patterson, L.T. Walker, J. Boateng and M. Verghese, 2015. Chemopreventive potential of ginger on Hep2G cells. Int. J. Cancer Res., 11: 52-66.
CrossRefDirect Link - Cheraghi, O., G. Dehghan, M. Mahdavi, R. Rahbarghazi and A. Rezabakhsh et al., 2016. Potent anti-angiogenic and cytotoxic effect of conferone on human colorectal adenocarcinoma HT-29 cells. Phytomedicine, 23: 398-405.
CrossRefDirect Link - Goutzourelas, N., D. Stagos, A. Housmekeridou, C. Karapouliou and E. Kerasioti, 2015. Grape pomace extract exerts antioxidant effects through an increase in GCS levels and GST activity in muscle and endothelial cells. Int. J. Mol. Med., 36: 433-441.
CrossRefDirect Link - Lima, C.F., M. Fernandes-Ferreira and C. Pereira-Wilson, 2006. Phenolic compounds protect HepG2 cells from oxidative damage: Relevance of glutathione levels. Life Sci., 79: 2056-2068.
CrossRefPubMedDirect Link - Singh, S., D.K. Singh, A. Meena, V. Dubey, N. Masood and S. Luqman, 2019. Rutin protects t‑butyl hydroperoxide-induced oxidative impairment via modulating the Nrf2 and iNOS activity. Phytomedicine, 55: 92-104.
CrossRefDirect Link - Crespo, R., B.E. Rodenak-Kladniew, M.A. Castro, M.V. Soberón and S.M.L. Lavarías, 2020. Induction of oxidative stress as a possible mechanism by which geraniol affects the proliferation of human A549 and HepG2 tumor cells. Chem. Biol. Interact., Vol. 320.
CrossRefDirect Link - Franco, R. and J.A. Cidlowski, 2009. Apoptosis and glutathione: Beyond an antioxidant. Cell Death Differ., 16: 1303-1314.
CrossRefPubMedDirect Link - Anderson, M.E., 1998. Glutathione: An overview of biosynthesis and modulation. Chem. Biol. Interac., 111-112: 1-14.
CrossRefDirect Link - Du, Z.X., H.Y. Zhang, X. Meng, Y. Guan and H.Q. Wang, 2009. Role of oxidative stress and intracellular glutathione in the sensitivity to apoptosis induced by proteasome inhibitor in thyroid cancer cells. BMC Cancer, Vol. 9.
CrossRefDirect Link - Kasala, E.R., L.N. Bodduluru, R.M. Madana, K.V. Athira, R. Gogoi and C.C. Barua, 2015. Chemopreventive and therapeutic potential of chrysin in cancer: Mechanistic perspectives. Toxicol. Lett., 233: 214-225.
CrossRefDirect Link - Ortega, A.L., S. Mena and J.M. Estrela, 2011. Glutathione in cancer cell death. Cancers, 3: 1285-1310.
CrossRefDirect Link