
Oscar Istas
Department of Biology, Center for Muscle Biology, University of Kentucky, Lexington, KY, 40506-0225, USA
Abigail Greenhalgh
Department of Biology, Center for Muscle Biology, University of Kentucky, Lexington, KY, 40506-0225, USA
Robin L. Cooper
Department of Biology, Center for Muscle Biology, University of Kentucky, Lexington, KY, 40506-0225, USA
LiveDNA: 1.523
Journal of Pharmacology and Toxicology
Year: 2020 | Volume: 15 | Issue: 2 | Page No.: 65-72







ABSTRACT
Background and Objective: Gram-negative bacteria produce and release endotoxins in the form of lipopolysaccharides (LPS), which can have direct effects on neural synaptic communication. Since the effects of acute repetitive exposures of LPS have not been fully addressed previously, the objective of this study was to investigate the repetitive effects of acute applications of LPS on synaptic transmission. Materials and Methods: The membrane potential of cells and the amplitude of evoked synaptic responses were measured with intracellular recordings. The larval Drosophila neuromuscular junction was used as a glutamatergic synaptic model. Results: LPS depressed synaptic transmission in a dose-dependent manner. LPS blocked the glutamate receptors while hyperpolarizing the postsynaptic cell. LPS induced hyperpolarization desensitized the cell to the effects of subsequent exposures to LPS. The responses from repeating short one minute exposures to LPS with 5 min of removal and washing with saline were reproducible over three trials. Over twenty minutes of incubation with LPS, the muscle fibers depolarized and did not recover with the removal of LPS as well as the glutamatergic synaptic responses. Conclusion: LPS directly blocks glutamatergic synaptic transmission. Cells can recover from rapid exposures to LPS within 1 min. Longer than 5 min of LPS exposure produces long term irreversible effects. Additive effects with low to higher concentrations of LPS occur. The mechanism of LPS hyperpolarizing skeletal muscle is unknown but occurs in larval Drosophila muscle.
PDF Abstract XML References Citation
How to cite this article
Oscar Istas, Abigail Greenhalgh and Robin L. Cooper, 2020. Repetitive Exposure to Bacterial Endotoxin LPS Alters Synaptic Transmission. Journal of Pharmacology and Toxicology, 15: 65-72.
DOI: 10.3923/jpt.2020.65.72
URL: https://scialert.net/abstract/?doi=jpt.2020.65.72
DOI: 10.3923/jpt.2020.65.72
URL: https://scialert.net/abstract/?doi=jpt.2020.65.72
INTRODUCTION
An estimated 1.7 million adult cases of sepsis occur annually in the United States, contributing to 265,000 deaths each year1,2. Gram-negative bacterial strains, such as Escherichia coli, Pseudomonas aeruginosa and Serratia marcescens are linked to septicemia. Gram-negative bacteria produce and release endotoxins in the form of lipopolysaccharides (LPS). The different forms of LPS elicit varying immune responses from a host. The muscular system is greatly affected after contracting sepsis and is key in the body’s response to bacterial endotoxins by releasing proinflammatory cytokines (TNF-α, IL-1, or IL-6)3.
LPS appears at low levels systemically in mammals normally after high fat meals, compromised intestinal barrier and a low grade or during the initial onset of gram-negative induced septicemia. It is of interest to know the responsiveness of tissues with a secondary bout of LPS exposure or an additional exposure while initial responses are being mediated potentially by desensitization, by a receptor mediate action, or even actions by ion exchangers or pumps. In severe bacterial sepsis the use of high doses of antibiotics is common to kill the bacteria, but in doing so, a surge of LPS may occur and cause a cytokine storm.
In the CNS of rodents, LPS is known to depress glutamatergic synaptic communication which may be due to action on microglia to indirectly influence neurons as well as acting directly on neurons4,5. Glutamatergic synapses rapidly depress at larval Drosophila neuromuscular junction (NMJ) in the presence of LPS and can be partially recovered upon rapid flushing to remove the LPS. The mechanism behind the rapid hyperpolarization and gradual depolarization of the target muscle towards the initial membrane potential in the presence of LPS has not yet been elucidated. The gradual depolarization of the membrane potential while still exposed to LPS may be due to desensitization in the initial effect of LPS. This potential receptor-mediated response or action on a pump or exchanger is plausible considering the gradual and transitory natural in the onset and offset in responses. One postulated mechanism to account for the muscle membrane potential is an initial hyperactivation of the sodium-potassium pump followed by desensitization to LPS5. However, this would not explain the effect of the amplitude of the evoked EJPs. In some trials, the amplitudes of the EJPs continue to decrease while the membrane potential is recovering, but in other cases, the amplitude of the EJPs increases while the membrane slowly depolarizes back to the initial level. To examine if the initial effects can be additive, the second exposure of LPS was examined while the physiological changes were occurring from the initial exposure as well as 5 min after the removal of the initial LPS exposure.
There are no known mechanistic explanations to explain how LPS hyperpolarizes the muscle membrane. The potential mechanisms were presented in a past paper5; however they could not account for the changes. So, it remains an open question of how the membrane hyperpolarizes. What is shown herein is that the responses in muscles are repetitive and the cells do not die with one high-level exposure which was stated for the effect on neurons in other studies. High levels of LPS in this preparation is not immediately toxic to cells which contradicts dogma. Earlier publications do not address the effect of repetitive exposures and how the preparations can be repeatedly washed out with saline and be re-exposed to LPS as reviewed in Ballinger-Boone et al.5. The purpose of this study was to investigate if these responses are reproducible in the Drosophila model with acute repetitive exposures to LPS as one may gain insight into the actions and/or mechanisms of LPS effects in other animals including humans.
MATERIALS AND METHODS
Experimental site: Experiments were conducted at the University of Kentucky in Lexington, Kentucky, USA from January to May 2020.
Materials and research tools: The segmental nerves were cut and sucked into a suction electrode, which was filled with saline and stimulated. The segmental nerves were stimulated at 0.5 Hz (S88 Stimulator, Astro-Med, Inc., Grass Co., West Warwick, RI, USA). To monitor the transmembrane potentials of the body wall muscle (m6) of 3rd instar larvae, a sharp intracellular electrode (30 to 40 M resistance) filled with 3M KCl impaled the fiber. An Axoclamp 2B (Molecular Devices, Sunnyvale, CA, USA) amplifier and 1 X LU head stage was used.
Fly saline modified haemolymph-like 3 (HL3) was used: (in mmol/L) 70 NaCl, 5 KCl, 20 MgCl2, 10 NaHCO3, 1 CaCl2, 5 trehalose, 115 sucrose, 25 N,N-bis (2-hydroxyethyl)-2-aminoethane sulfonic acid (BES) and pH at 7.1. Canton S (CS) flies were used and have been isogenic in the lab for several years. The stock was originally obtained from the Bloomington Drosophila Stock Center (BDSC).
Research procedure: The overall protocol was to stimulate the segmental nerve at 0.5 Hz while recording the EJPs and mEJPS as well as the resting membrane potential before, during exposure to LPS and when LPS was removed from the bath.
The technique to dissect larvae was followed as described earlier6,7. In brief, a longitudinal dorsal midline cut was made in 3rd instar larvae to expose the CNS.
LPS was dissolved in saline prior to use and was readily exchanged over the dissected preparations during the recording of evoked EJPs and mEJPs. The total volume of the chamber was only 1 mL, which was fully exchanged when switching the media. One form of LPS used was Serratia marcescens (S.m.). LPS and the chemicals used for saline were obtained from Sigma-Aldrich (St. Louis, MO, USA). LPS concentration was used at 500 μg mL–1 to compare with previous studies using LPS on the larval Drosophila muscles as well as frog and crayfish muscles and rodent CNS5,8-11. The LD50 in rodents for LPS10 from S.m. is 650 μg mL–1 (6×106 CFU- colony-forming units)12. This was another reason to use a relatively high concentration for D. melanogaster, since they are likely exposed to gram-negative bacterial strains in their native environment.
Data collection: Electrical signals were recorded on-line to a Power Lab/4s interface (ADInstruments, Australia) and calibrated with the Powerlab Chart software version 7.
Experimental design: The effect on the membrane potential and amplitude of evoked EJPs were examined for brief repetitive exposures of 1 min of exposure to LPS followed by 5 min of flushed saline without LPS and was repeated two more times. In addition, repetitive exposures to LPS were examined without washing out the LPS between the exposures. To examine the effect of varying concentrations a low concentration (100 μg mL–1) was used followed by a higher concentration (500 μg mL–1). Long exposures of 25 min to LPS at high concentration followed by removal of LPS were examined for examining if the responses were reversible after prolonged exposure.
Parameters measured: The membrane potential and amplitudes of evoked transmission after changing bathing conditions to different compounds were measured.
Statistical analysis: Statistical analysis was performed as a Sign pairwise test for changes in membrane potential and amplitudes of evoked transmission after changing bathing conditions. Since some data sets are not normally distributed, (a number of zeroes in some groups) the non-parametric Sign test was used. A significant difference is considered p<0.05. Trends in the same direction were used to establish consistency.
RESULTS
Rapid effects of intermittent repetitive LPS exposure: To examine the effects of brief repetitive exposures, 1 min of exposure to LPS followed by 5 min of flushed saline without LPS was conducted and repeated. The subsequent responses of depressed evoked glutamatergic synaptic transmission were not able to recover fully during the removal of LPS (Fig. 1). The rapid hyperpolarization of the muscle was reproducible each time and obtained approximately the same level (Fig. 1). These trends were consistent in 6 out 6 preparations (p<0.05, Sign test, N = 6). LPS was presented for 1 min followed by 5 min of removal in normal saline for 3 consecutive trials.
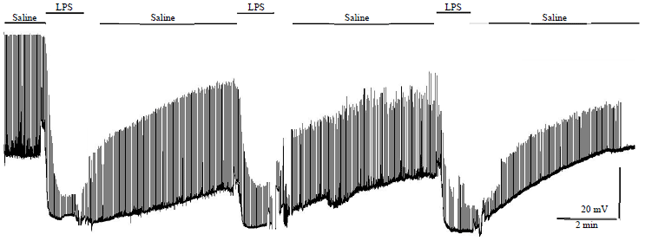 | |
| Fig. 1: | Repetitive exposure to LPS (500 μg mL–1) with the removal of LPS between exposures |
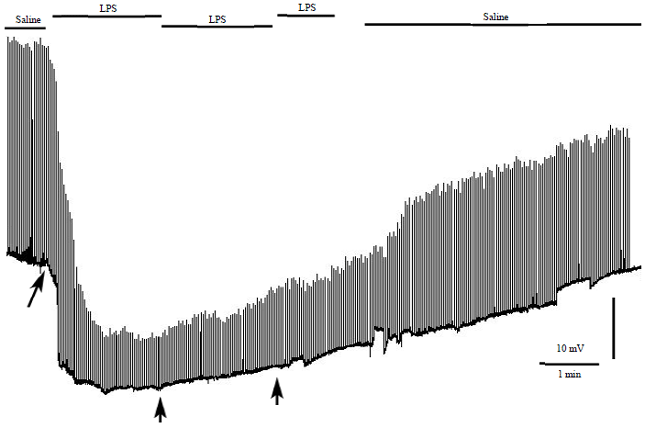 | |
| Fig. 2: | Repetitive continuous exposure to LPS (500 μg mL–1). |
LPS was presented for 1 min, followed by exchanging the bath to the same concentration of LPS for 3 consecutive trials, followed by removal of LPS | |
Each exposure resulted in rapid hyperpolarization of the membrane potential and a rapid reduction in the amplitude of the evoked Excitatory Junction Potential (EJP). The membrane potential drifted toward the initial membrane potential during the removal of LPS and the amplitude of the EJPs increased during the removal of LPS. With each exposure to LPS, the ability to recover to the initial amplitude of the EJP was reduced.
Repetitive constant LPS exposure: To examine if repetitive exposures recreated the initial response, LPS was repetitively introduced to the preparation without exchanging the bathing media (Fig. 2). The subsequent exposures every minute did not alter the response in producing an additional hyperpolarization or further dampening in the amplitude of the EJP. The effect of the overall 3 to 4 min of exposure to LPS was partially overcome by switching to saline without LPS (Fig. 2). The fact that a new exposure flushing the bath did not result in a subsequent hyperpolarization or had an effect on the amplitude of the EJP would suggest then that the LPS present from the initial exposure was not being degraded in the synapse. The constant bath exposure of LPS is sufficiently exposing the preparation without a flow though system during the exposure time. These trends were consistent in 6 out 6 preparations (p<0.05, Sign test, N = 6). The membrane potential drifted toward the initial membrane potential and amplitude of EJPs increased during LPS exposure. With each additional exposure to LPS (Fig. 2, arrows), the response appeared as if there was only single exposure to LPS. Thus, the response appears to show some desensitization to LPS. Removal of LPS promotes a quicker recovery toward initial conditions. This same trend was observed in six out of six preparations.
Dose dependent actions of LPS exposure: The fact that repetitive LPS exposures at 500 μg mL–1 did not produce an additional response and that the induced response was dampened indicated a saturating effect at a concentration of 500 μg mL–1. Thus, an initial exposure at a concentration of 100 μg mL–1 for 1 min was followed with subsequent exposure of 500 μg mL–1 LPS (Fig. 3). The increased concentration produced an additive effect with further hyperpolarization and rapid dampening in the amplitude of the evoked EJP (Fig. 3).
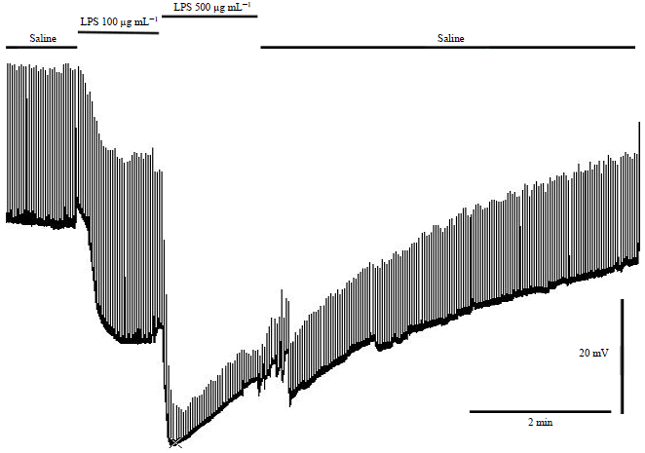 | |
| Fig. 3: | Repetitive exposure to LPS at an increasing concentration (100 to 500 μg mL–1) during continuous incubation with LPS |
These trends were consistent in 6 out 6 preparations (p<0.05, Sign test, N = 6). The additional exposure to 500 μg mL–1) had an additional effect on the membrane potential as well as the amplitude of the EJPs. The LPS exposure at 100 μg mL–1 predominantly affected the membrane potential while LPS exposure at 500 μg mL–1 rapidly affected both the hyperpolarization of the membrane potential as well as caused a reduction in the amplitude of the EJP. This same trend was observed in 6 out of 6 preparations.
Prolonged LPS exposure: The effect of a short term exposure of 1 to 3 min of LPS can be partially overcome with the removal of the LPS; however, the repetitive exposures with removing LPS resulted in a continuous depression in the amplitude of the EJPs. The removal of LPS did not revitalize the amplitude in the EJPs. To address the effect of relatively long-term exposure to LPS, preparations were bathed in LPS-containing-saline for 25 min while examining the membrane potential and the evoked responses (Fig. 4). The initial rapid hyperpolarization was slowly lost as the membrane depolarized above the initial resting membrane potential. The depressed amplitude of the EJP slowly recovered but in time depressed to the point of being undetectable despite the presence of a driving gradient (Fig. 4). After thoroughly exchanging the bathing media with fresh saline, not containing LPS, the membrane potential shows a slight recovery and the EJPs are present but still suppressed (Fig. 4). These trends were consistent in 9 out 9 preparations (p<0.02, Sign test, N = 9). In examining other muscles in the same preparation after 25 min of LPS exposure and saline flushing but not monitored with an intracellular electrode during the exposure, the membrane potentials were also depolarized and were easily damaged with the recording. When the bathing media was exchanged thoroughly three times, a gradual recovery of the membrane potential occurred along with the amplitude of the EJP. The lower enlarged trace (Fig. 4) depicts a spontaneous quantal event and the first EJP in the evoked responses. This same trend was observed in 9 out of 9 preparations.
Exposure to LPS (500 μg mL–1) for 25 min revealed the transient hyperpolarization and depression in EJP amplitude, with a gradual recovery of the membrane potential, but with a second delayed depression in the amplitude of the EJP, along with a loss of membrane potential.
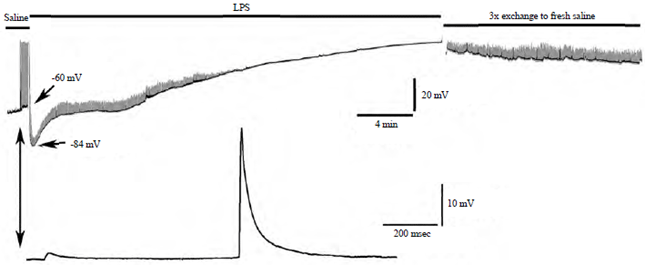 | |
| Fig. 4: | Long term continuous exposure to LPS |
DISCUSSION
This study illustrated that LPS can be repetitively applied with the effects partially reversed if only acute exposures occur. The rapid response in hyperpolarizing the membrane potential while reducing the evoked EJP suggests two different direct actions. One in hyperpolarizing the membrane while increasing the driving gradient for the EJP but due to the addition of blocking of glutamate receptors the amplitude of the EJPs was reduced. The transient effect of hyperpolarization and repolarization of the muscle membrane potential can be induced repetitively if LPS is removed but with continuous exposure the effect is transient. The long exposure to a high concentration of LPS of 25 min produces damaging effects and not readily reversed.
Direct actions of LPS on cells is an important topic to address as this is an initial step in triggering the secondary responses of cytokine release from multiple cell types. Blocking or modifying LPS response may lead to treatments of pathological ailments associated with gram-negative bacterial septicemia. Blocking one of the LPS induced responses in cultured rodent muscle was shown to block the induced atrophy13.
In mammals, it is known that LPS binds to a Toll-like receptor 4 (TLR4), now referred to as the CD14/TLR4/MD2 receptor complex in mammals14,15. Toll receptors were first discovered in Drosophila melanogaster, which led to the discovery of these receptors for LPS in mammals. These receptors are not the sole receptors for the gram-negative LPS to induce a response in Drosophila nor is it likely that the Immune deficiency (Imd) signaling pathway is the key for some of the reported responses. The peptidoglycan recognition proteins (PGRPs) are known to mediate an immune response in Drosophila. Of the 13 PGRPs identified by genome analysis of Drosophila, 3 were linked with the immune response16,17. Of these three PGRPs (PGRP-SA, PGRP-LC and PGRP-LE), PGRP-LC and PGRP-LE have been shown to respond to gram-negative bacteria18,19. However, using RNAi constructs of PGRP-LC and PGRP-LE for expression in Drosophila body wall and cardiac muscle, as well as neurons, did not alter the responses to direct exposure of LPS5.
The actions induced by LPS on the larval Drosophila muscle are two-fold. One response is the blocking of the glutamate receptors, which reduces the amplitude of evoked EJPs and was shown previously to gradually reduce the amplitude of the spontaneous quantal responses10,11. This indicated that the response is mediated on the postsynaptic cell. The glutamatergic synaptic responses in the rodent CNS is also reduced, but how this occurs is not well established. It appears there is a glial cell involvement (i.e., astrocytes, oligo) resulting in a release of cytokines (TNF-α and IL-1) through activated nuclear factor kappa B (NF-κB)20-23 and leading to cellular toxicity. However, in short, brief exposures of LPS, the synaptic responses can be recovered5. It has been shown that there are direct effects on cultured hippocampal neurons in culture neurons, independent from those on glial cells4. Separating out the direct and indirect responses of LPS on mammalian neurons is an important topic for future research. Interestingly, the glutamatergic receptor subtype at the larval Drosophila NMJ is of a quisqualate type, like in other insects and crustaceans24-26. It is of interest to know if other glutamatergic receptor subtypes are also blocked by LPS.
The other action of LPS on the muscle fibers is the rapid hyperpolarization and gradual depolarization. The mechanism to explain this phenomenon remains elusive and is also observed for the skeletal muscle of the crayfish27. The response is not likely due to an induction of calcium-activated potassium conductance, a TEA sensitive potassium channel or mediated through Nitric Oxide Synthase (NOS) as each of these possibilities has been addressed5,10. In addition, the response is not due to opening a chloride channel as the equilibrium potential for Cl– in larval Drosophila body wall muscles is more depolarized than the resting membrane potential28,29. Even the cardiac muscle cells of the larval Drosophila are directly affected by LPS exposure and are not innervated9. It would be of interest to know if neurons also demonstrate a transient hyperpolarization like the muscles in Drosophila and crayfish.
The LD50 in rodents for LPS from S. m. is 650 μg/ml (6x106 CFU- colony-forming units)12. Thus, we chose to use a relatively high concentration of 500 μg mL–1 for D. melanogaster since they are likely exposed to gram-negative bacterial strains in their native environment on decaying fruit in the outdoor environment. The concentration of LPS in humans for mild cases is lower; however, to accentuate the responses experimentally allows insight into what more subtle responses may be occurring at lower concentrations.
The hyperpolarization of the muscle is a more rapid and sensitive response to LPS than the reduction in the EJP amplitude, as lower concentrations of LPS induce the hyperpolarization while not producing a significant response on the evoked responses. In fact, it may increase the amplitude of the evoked EJP due to the large driving gradient for the ionotropic glutamate receptors as observed in Fig. 3 and reported for two different forms of LPS (i.e., Pseudomonas aeruginosa and Serratia marcescens) on Drosophila muscle10. This finding implies that the mechanism for inducing the hyperpolarization has a higher affinity than LPS blocking the glutamate receptor. Considering the same concentration and form of LPS causes hyperpolarization in crayfish muscle but not as pronounced whereas the effect on frog muscle is minimal suggest that there maybe a commonality in the mechanism of action on the muscle for arthropods but different for amphibians. However, the synaptic responses are enhanced for the glutamatergic synapses at the crayfish NMJ but dampen the response for the larval Drosophila NMJ and the cholinergic frog NMJ.
CONCLUSION
The effects of LPS can be reversed if acute exposures of a few minutes occur and then LPS is flushed away from the tissues. The rapid hyperpolarizing the muscle membrane potential is recovered quickly if LPS is removed but the effect is transient even with long exposure times. However, the membrane of the muscle will continue to depolarize indicating damage to the cell membrane integrity with long exposure. Repetitive exposure of a low concentration followed by a high concentration accentuates the effects of LPS. The blocking of the glutamate receptors is also able to be reversed with a short exposure of a few minutes.
SIGNIFICANCE STATEMENT
The findings of this study are important for understanding the potential effects and treatments in acute and longer-term exposure to LPS. A rapid removal of LPS is important for being able to reverse the effects on the membrane potential of the muscle as well as removing the blocking action of glutamate receptors. In speculating a potential mechanism for the hyperpolarization, transient activation of the Na-K pump could occur and then, as the membrane potential slowly recovers by desensitization of LPS on the Na-K pump. Future comparative studies examining more cell types and different species than those recently examined for frog, crayfish, rodent and Drosophila would help in determining the direct mechanisms of action by LPS on tissues for potential therapeutic actions.
ACKNOWLEDGMENTS
The author would like to thank Chellgren Endowed Professor fund (RLC) for supporting this research work.
REFERENCES
- Dantes, R.B. and L. Epstein, 2018. Combatting Sepsis: A public health perspective. Clin. Infect. Dis., 67: 1300-1302.
CrossRefDirect Link - Rhee, C., T.M. Jones, Y. Hamad, A. Pande and J. Varon et al., 2018. Prevalence, underlying causes, and preventability of sepsis-associated mortality in US acute care hospitals. JAMA Netw. Open, Vol. 2.
CrossRefDirect Link - Costamagna, D., P. Costelli, M. Sampaolesi and F. Penna, 2015. Role of inflammation in muscle homeostasis and myogenesis. Mediators Inflammation, 2015: 1-14.
CrossRefDirect Link - Calvo-Rodriguez, M., C. de la Fuente, M. Garcia-Durillo, C. Garcia-Rodriguez, C. Villalobos and L. Nunez, 2017. Aging and amyloid β oligomers enhance TLR4 expression, LPS-induced Ca2+ responses and neuron cell death in cultured rat hippocampal neurons. J. Neuroinflamm., Vol. 14.
CrossRefDirect Link - Ballinger-Boone, C., O. Anyagaligbo, J. Bernard, S.M. Bierbower and E.E. Dupont-Versteegden et al., 2020. The effects of bacterial endotoxin (LPS) on cardiac and synaptic function in various animal models: Larval Drosophila, crayfish, crab, and rodent. Int. J. Zool. Res., Vol. 16.
CrossRefDirect Link - Dasari, S. and R.L. Cooper, 2004. Modulation of sensory-CNS-motor circuits by serotonin, octopamine and dopamine in semi-intact Drosophila larva. Neurosci. Res., 48: 221-227.
CrossRefPubMedDirect Link - Stewart, B.A., H.L. Atwood, J.J. Renger, J. Wang and C.F. Wu, 1994. Improved stability of Drosophila larval neuromuscular preparations in haemolymph-like physiological solutions. J. Comp. Physiol. A., 175: 179-191.
CrossRefDirect Link - De Castro, C., J. Titlow, Z.R. Majeed and R.L. Cooper, 2014. Analysis of various physiological salines for heart rate, CNS function and synaptic transmission at neuromuscular junctions in Drosophila melanogaster larvae. J. Comp. Physiol. A, 200: 83-92.
CrossRefDirect Link - Anyagaligbo, O., J. Bernard, A. Greenhalgh and R.L. Cooper, 2019. The effects of bacterial endotoxin (LPS) on cardiac function in a medicinal blow fly (Phaenicia sericata) and a fruit fly (Drosophila melanogaster). Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 217: 15-24.
CrossRefDirect Link - Cooper, R.L., M. McNabb and J. Nadolski, 2019. The effects of a bacterial endotoxin LPS on synaptic transmission at the neuromuscular junction. Heliyon, Vol. 5, No. 3.
CrossRefDirect Link - Istas, O., A. Greenhalgh and R. Cooper, 2019. The effects of a bacterial endotoxin on behavior and sensory-CNS-motor circuits in Drosophila melanogaster. Insects, Vol. 10, No. 4.
CrossRefDirect Link - Iwaya, A., S. Nakagawa, N. Iwakura, I. Taneike and M. Kurihara et al., 2005. Rapid and quantitative detection of blood Serratia marcescens by a real-time PCR assay: Its clinical application and evaluation in a mouse infection model. FEMS Microbiol. Lett., 248: 163-170.
CrossRefDirect Link - Hahn, A., M. Kny, C. Pablo-Tortola, M. Todiras and M. Willenbrock et al., 2019. Serum amyloid A1 mediates myotube atrophy via Toll-like receptors. J. Cachexia, Sarcopenia Muscle.
CrossRefDirect Link - Yoshida, H., K. Kinoshita and M. Ashida, 1996. Purification of a peptidoglycan recognition protein from hemolymph of the silkworm, Bombyx mori. J. Biol. Chem., 271: 13854-13860.
CrossRefDirect Link - Steiner, H., 2004. Peptidoglycan recognition proteins: On and off switches for innate immunity. Immunol. Rev., 198: 83-96.
CrossRefDirect Link - Leclerc, V. and J.M. Reichhart, 2004. The immune response of Drosophila melanogaster. Immunol. Rev., 198: 59-71.
CrossRefDirect Link - Werner, T., G. Liu, D. Kang, S. Ekengren, H. Steiner and D. Hultmark, 2000. A family of peptidoglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc. Natl. Acad. Sci. USA., 97: 13772-13777.
CrossRefDirect Link - Gottar, M., V. Gobert, T. Michel, M. Belvin and G. Duyk et al., 2002. The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature, 416: 640-644.
CrossRefDirect Link - Takehana, A., T. Katsuyama, T. Yano, Y. Oshima, H. Takada, T. Aigaki and S. Kurata, 2002. Overexpression of a pattern-recognition receptor, peptidoglycan-recognition protein-LE, activates imd/relish-mediated antibacterial defense and the prophenoloxidase cascade in Drosophila larvae. Proc. Natl. Acad. Sci. USA., 99: 13705-13710.
CrossRefDirect Link - Pérez‐Otaño, I., M.K. McMillian, J. Chen, G. Bing, J.S. Hong and K.R. Pennypacker, 1996. Induction of NF‐kB‐like transcription factors in brain areas susceptible to kainate toxicity. Glia, 16: 306-315.
CrossRefDirect Link - Friedman, W.J., S. Thakur, L. Seidman and A.B. Rabson, 1996. Regulation of nerve growth factor mRNA by interleukin-1 in rat hippocampal astrocytes is mediated by NFκB. J. Biol. Chem., 271: 31115-31120.
CrossRefDirect Link - Hartlage-Rubsamen, M., R. Lemke and R. Schliebs, 1999. Interleukin-1β, inducible nitric oxide synthase and nuclear factor-κB are induced in morphologically distinct microglia after rat hippocampal lipopolysaccharide/interferon-γ injection. J. Neurosci. Res., 57: 388-398.
CrossRefPubMedDirect Link - Nomura, Y., 1998. A transient brain ischemia- and bacterial endotoxin-induced glial iNOS expression and NO-induced neuronal apoptosis. Toxicol. Lett., 102: 65-69.
CrossRefDirect Link - Bhatt, D. and R.L. Cooper, 2005. The pharmacological and physiological profile of glutamate receptors at the Drosophila larval neuromuscular junction. Physiol. Entomol., 30: 205-210.
CrossRefDirect Link - Lee, J.Y., D. Bhatt, D. Bhatt, W.Y. Chung and R.L. Cooper, 2009. Furthering pharmacological and physiological assessment of the glutamatergic receptors at the Drosophila neuromuscular junction. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 150: 546-557.
CrossRef - Parrot, S. and L. Denoroy, 2018. Biochemical Approaches for Glutamatergic Neurotransmission. 1st Edn., Humana Press, United States, Pages: 565.
CrossRefDirect Link - Saelinger, C.M., M.C. McNabb, R. McNair, S. Bierbower and R.L. Cooper, 2019. Effects of bacterial endotoxin on regulation of the heart, a sensory-CNS-motor nerve circuit and neuromuscular junctions: Crustacean model. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., Vol. 237.
CrossRefDirect Link - Rose, U., C. Derst, M. Wanischeck, C. Marinc and C. Walther, 2007. Properties and possible function of a hyperpolarisation-activated chloride current in Drosophila. J. Exp. Biol., 210: 2489-2500.
CrossRefDirect Link - Stanley, C.E., A.S. Mauss, A. Borst and R.L. Cooper, 2019. The effects of chloride flux on Drosophila heart rate. Methods Protocols, Vol. 2, No. 3.
CrossRefDirect Link