
J. Patterson
Nutritional Biochemistry and Toxicology Laboratory, Department of Food and Animal Sciences, Alabama Agricultural and Mechanical University, P.O. Box 1628, Normal, 35762, Alabama, USA
M. Verghese
Nutritional Biochemistry and Toxicology Laboratory, Department of Food and Animal Sciences, Alabama Agricultural and Mechanical University, P.O. Box 1628, Normal, 35762, Alabama, USA
Journal of Pharmacology and Toxicology
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 49-59







ABSTRACT
Curry leaves (Murraya koenigii), native to South Asian countries are commonly used as a food flavoring agent and known to have antioxidant, anti-inflammatory and antidiabetic activities. The objectives of this study were to investigate the anticancer and toxic effects of Curry Leaf Extracts (CLE) in Caco-2 and HepG2 cells and to assess the phytochemical content and antioxidant activities of CLE. The Caco-2 human colon adenocarcinoma (ATCC HTB-37) cells and Hep2G human liver cells (ATCC HB-8065) and were obtained from the American Type Culture Collection (ATCC, Manassas, VA). For assay, 5×104cells/well were seeded in a 24 well culture plate and incubated at 37°C and 5% CO2 until development of a monolayer. Cells were incubated with aqueous CLE at selected concentrations (0.2-0.8 μg mL–1) for 12 and 24 h. Lactate dehydrogenase (LDH) release, histone-related DNA fragmentation and detoxification enzymes were determined. Phytochemical content and antioxidative activities of the extract were determined using in vitro methods. After 24 h, LDH release was highest in Caco-2 cells treated at 0.8 μg mL–1 (45.83 μg mL–1). The LDH of HepG2 cells peaked at 15.74% (12 h-0.8 μg mL–1), inferring low acute cytotoxicity. The GST activity in Caco-2 cells increased with increasing concentration of CLE, however, SOD activity decreased. Histone-related DNA fragmentation was highest in cells treated with 0.4 μg mL–1 for both cell lines. Total phenolics and flavonoids of methanolic CLE were 541.11±12.41 GAE/100 g CLE and 192.22±2.75 CE/100 g CLE. The results from this study indicate that curry leaf may be effective in colon cancer prevention while causing minimal toxic effects to liver cells.
PDF Abstract XML References Citation
Received: July 03, 2015;
Accepted: October 01, 2015;
Published: October 12, 2015
How to cite this article
J. Patterson and M. Verghese, 2015. Anticancer and Toxic Effects of Curry Leaf (Murraya koenigii) Extracts. Journal of Pharmacology and Toxicology, 10: 49-59.
DOI: 10.3923/jpt.2015.49.59
URL: https://scialert.net/abstract/?doi=jpt.2015.49.59
DOI: 10.3923/jpt.2015.49.59
URL: https://scialert.net/abstract/?doi=jpt.2015.49.59
INTRODUCTION
Colorectal cancer is the third leading cause of death in the U.S. and affects over 1 million people a year (Johnson and Mukhtar, 2007; Manzano and Perez-Segura, 2012). Epidemiological studies suggest the consumption of herbs and spices may be a factor in reducing the incidences of gastric and colon cancers (Mohandas and Desai, 1998). In vitro studies have linked the chemopreventive effects of herbs and spices (specifically garlic, turmeric) to one or more of the following mechanisms: inhibition of oxidants, inflammation, carcinogenic bioactivation and tumorigenesis (Johnson and Mukhtar, 2007; Gupta and Prakash, 2009).
Curry leaf (Murraya koenigii), belonging to the Rutaeae family, is native to India, Sri Lanka, Bangladesh and other South Asian countries (Gupta et al., 2011). It is currently used a cooking spice and has been commonly used in traditional Indian medicine for thousands of years (Gupta et al., 2011). Studies report curry leaves to have hypoglycemic, antidiabetic (Xie et al., 2006; Dineshkumar et al., 2010), hepatoprotective (Sathaye et al., 2011), antibacterial (Ningappa et al., 2010; Tajkarimi et al., 2010; Biswas et al., 2012), anti-inflammatory (Gupta et al., 2010) and antioxidative properties (Ningappa et al., 2008; Gupta and Prakash, 2009). The bioactivity of curry leaf has been attributed to the presence of phytochemicals including alkaloids, essential oils, phenolic acids (Singh et al., 2011), terepenoids, tocopherol, β-carotene, lutein (Palaniswamy, 2001), as well as minerals, protein and fat (Singh et al., 2011). Furthermore, Mani et al. (2012) suggest the alkaloids of curry leaf as a useful remedy for dementia and Alzheimer’s disease.
Many phytochemicals, phenolics for example, have been proven to be natural antioxidants. Natural antioxidants exhibit their potential through a variety of mechanisms such as termination of free-radicals and reduction of toxic ions to non toxic ions (Ningappa et al., 2008).
Though curry leaf has many positive effects in the body, the chemopreventive mechanisms of curry leaf are not fully understood. One study suggests it involves multiple biochemical and biological mechanisms including the induction of enzymes and anti-oxidation (Dasgupta et al., 2003). Induction of detoxification (phase II) enzymes have been found to play a major role in metabolizing potentially toxic and carcinogenic compounds (Yoshimasa et al., 2000). Such enzymes include glutathione-S-tranferase (detoxification) and antioxidative enzymes superoxide dismutase and catalase. Therefore, the objectives of this study were to investigate the anticancer and toxic effects of Curry Leaf Extracts (CLE) in Caco-2 and HepG2 cells and to assess the phytochemical content and antioxidant activities of CLE.
MATERIALS AND METHODS
Sample preparation: Five grams of dried curry leaf (obtained from plant in Florida, US) was added to 50 mL of 80% methanol and stirred for 2 h. Samples were centrifuged at 3000 g for 20 min and the supernatant collected. Supernatant was evaporated at 40°C to dryness. Samples were resuspended in 80% methanol and stored at -80°C until further analysis.
Determination of total phenolic content: Total phenolic content of curry leaf was determined using a modified Folin-Ciocalteu colorimetric method as described by Singleton et al. (1999). Briefly, 12.5 μL of sample was added to 50 μL of distilled water. Then, 12.5 μL of Folin-Ciocalteu’s phenol reagent was added to the mixture. After 5 min, 125 μL of 7% NaCO3 solution was added and the samples were incubated for 90 min at 25°C. The absorbance was measured at 750 nm using a microplate reader (Synergy HT, BioTek instruments, USA). A standard curve for total phenolics was constructed using a gallic acid standard solution. The results are expressed as milligram gallic acid equivalents/100 gram sample.
Determination of total flavonoid content: A colorimetric assay (Kim et al., 2003) with some modifications was used to quantify total flavonoid content. Briefly, 25 μL of Curry Leaf Extract (CLE) was added to 125 μL of ddH2O. Subsequently, 7.5 μL of 5% NaNO2 was added to the mixture. After the mixture was allowed to stand for 5 min, 15 μL of 10% AlCl3 was added. The mixture was incubated at ambient temperature (25°C) for an additional 5 min. Following that, 50 μL of 1 M NaOH was added to the mixture. The mixture was immediately diluted by the addition of 27.5 μL of ddH2O and the absorbance of the mixture was measured at 510 nm against a blank prepared with ddH2O using a microplate reader (Synergy HT, BioTek instruments, USA). (+)- Catechin was used as standard and the results are expressed as milligram of catechin equivalents/100 gram sample.
Determination of Ferric Reducing Antioxidant Power (FRAP): The FRAP assay was conducted as described by Benzie et al. (1999). One hundred μL of diluted sample was combined with 3 mL of freshly prepared FRAP reagent consisting of 300 mM acetate buffer (pH 3.6), 10 mM 2,4,6-tri(2-pyridyl)-s-triazine (TPTZ) in 40 mM HCL and 20 mM ferric chloride (FeCL3.H2O). The mixture was subsequently incubated for 10 min at 37°C. Following incubation, the samples were analyzed at an absorbance of 593 nm. The change in absorbance was compared to a standard ferrous sulphate (FeSO4.7H2O) (0.1-1.0 mM). The samples were analyzed in triplicates and the concentration of Fe3+ was expressed as micromole of Fe2+/g.
Free radical scavenging activity: Briefly, 0.1 mM solution of DPPH (2, 2-Diphenyl-1-picryl-hydrazyl) in methanol was prepared and 70 μL of the solution was added to 210 μL of CLE at different concentrations (25, 50, 75, 100, 125, 150, 200 and 250 mg mL–1). The absorbance was measured using a microplate reader (Synergy HT, BioTek instruments, USA) after 30 min at 517 nm. Free radical scavenging activity was measured as the amount of extract required to decrease the initial absorbance (517 nm) of DPPH radical concentration by 50% (IC50) as compared to the control according to the equation:
Cell culture general procedures: Caco-2 colon adenocarcinoma (ATCC HTB-37) and HepG2 human liver cells (ATCC HB8065) were obtained from the American Type Culture Collection (ATCC, Manassas, VA). Cells were maintained in Dulbecco’s Modified Eagles Media (DMEM) with 10% fetal bovine serum. For assay, 5×104 cells/well were seeded in a 24 well culture plate and incubated at 37°C and 5% CO2 until development of a monolayer. After the development of a monolayer, 400 μL of fresh media (DMEM serum free) were added to the wells. Aqueous curry leaf extracts were made up to 100 μL with saline at various concentrations (0.1-0.8 μg mL–1), added and incubated for 12 and 24 h.
Determination of lactate dehydrogenase (LDH): The cytotoxic effects of the CLE were measured by the release of lactate dehydrogenase (LDH) from the cell into the culture supernatant. The cells were incubated with the extracts for 12 and 24 h. A 10% triton-X solution was used as a positive control and the supernatant was collected. The LDH was quantified by colorimetric cytotoxicity detection kit (LDH) (Roche Diagnostics, Indianapolis, IN) according to the manufacturer’s instructions. The absorbance was read at 490/600 nm.
Glutathione-S-transferase activity in cell lysates: Caco-2 and HepG2 cells were incubated with CLE at varying concentrations for 12 and 24 h. The supernatant was collected after centrifuging at 10,000×g for 10 min in PBS buffer (pH 7) and used for GST assay using a microplate reader (BioTek Synergy HT) and kinetic absorbances were read at 340 nm at 5 min intervals.
Determination of catalase activity: Cell catalase activity was determined using a microplate reader (BioTek Synergy HT) at 240 nm as described by Aebi (1984). The reaction mixture (1 mL) contained 0.02 mL of suitably diluted cytosol in phosphate buffer (50 mM, pH 7.0) and 0.1 mL of 30 mM H2O2 in phosphate buffer. The specific activity of catalase was expressed as moles of H2O2 reduced per minute per milligram protein.
Determination of superoxide dismutase (SOD): Superoxide dismutase enzyme was estimated using assay kits according to manufacturer’s instructions (Cayman Chemicals, Ann Arbor, MI).
Histone-related DNA fragmentation: To determine the effect of CLE in induction of apoptosis, 1×104 cells were treated with CLE for 12 and 24 h. After treatment, cells were washed with PBS and cell lysates were prepared using cell lysis buffer and centrifuged at 200×g for 10 min. Apoptosis was determined by measuring internucleosomal DNA fragmentation by using a cell death detection ELISA kit (Cell Death Detection ELISA, Roche Molecular Biochemicals, Indianapolis, IN) according to the instructions provided by the manufacturer. Spectrophotometric analysis was carried out using an ELISA plate reader at 405 nm for the histone-related DNA fragmentation.
Morphological changes: Morphological changes in Caco-2 and HepG2 cells after incubation with CLE were determined by microscopic evaluation using a Nikon CT 120 microscope (Nikon Instruments Inc, Melville, NY) and photographs were taken using a SPOT 2.0 digital camera. Cells in slide flaskets were incubated for 24 h with CLE and stained with Giemsa stain (0.1% in PBS) for 5 min. The stain was washed three times with PBS and the plates were dried at 37°C and photographs were taken (Verghese et al., 2006).
Statistical analysis: All assays were conducted in triplicate over a duration of five months in 2014. Statistical analysis was conducted using SAS 9.1, 2004 (SAS Institute Inc., Cary, NC) using analysis of the variance (One-Way ANOVA) to determine any significant differences among the groups. A separate analysis was used for each time period. Results were presented as Mean±Standard Error Mean (SEM). Where significant, means were separated using Tukey’s studentized range test. The level of significance was set at p<0.05.
RESULTS
Total phenolic, flavonoids, ferric-reducing potential and DPPH scavenging ability: Total phenolics and flavanoid content (Table 1) were 541.11 mg GAE/100 g and 192.22 catechin equivalents, respectively. Ferric reducing antioxidant potential of CLE was found to be 702.81 μmol Fe+2/g.
DPPH scavenging ability is shown in Fig. 1. The IC50 of CLE was 63.85 mg mL–1 for 0.5 mM of DPPH. Percent inhibitions of DPPH radical for 25, 50, 75, 100, 125, 150, 200 and 250 mg mL of CLE were 29.67,43.30,51.93,62.81,64.67,68.87, 72.26 and 88.61, respectively.
| Table 1: | Total phenolics, flavonoids and ferric reducing antioxidant potential of curry leaf extracts |
 | |
| Values are Means±SEM, (n=3), CE: Catechin equivalents, CL: Curry leaf; FRAP: Ferric reducing antioxidant power, GAE: Gallic acid equivalents | |
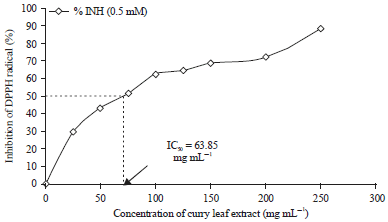 | |
| Fig. 1: | Free radical scavenging activity of curry leaf extract, DPPH: 2,2-diphenyl-1-picryl-hydrayl and IC50: Concentration in which 50% of radical is inhibited |
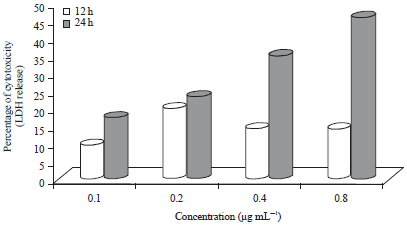 | |
| Fig. 2: | Percentage of cytotoxicity of curry leaf treated Caco-2 cells, LDH: Lactate dehydrogenase |
LDH of CLE-treated Caco-2 and HepG2 cells: Percent cytotoxicity (LDH release) compared to the control for Caco2 cells is shown in Fig. 2. Percentage of cytotoxicity was highest in Caco-2 cells treated with 0.8 μg mL–1 (55%) CLE and significantly higher (p<0.05) than all treatments except 0.2 and 0.4 μg mL–1 at 12 h of incubation. After 12 h, percentage of cytotoxicity increased dose-dependently from 0.1-0.4 μg mL–1, followed by a slight decrease at 0.8 μg mL–1 (17.18%). The LDH release of cells treated for 24 h were almost 4, 3, 2 and 3 folds higher than corresponding extract treatments at 12 h for 0.1, 0.2, 0.4 and 0.8 μg mL–1, respectively.
Percentage of cytotoxicity (LDH release) compared to the control for HepG2 cells is shown in Fig. 3. After 12 h of incubation, percentage of cytotoxicity increased with increasing concentration, peaking at 15.74% (0.8 μg mL–1). The LDH was significantly higher (p>0.05) with cells treated at 12 h with 0.8 μg mL–1 CLE compared to all other treatments. The LDH release for cells treated with 0.1, 0.2 and 0.8 μg mL–1 of curry leaf were approximately 2-fold higher at 12 h, compared to corresponding treatments at 24 h. Cells incubated for 24 h had very low cytotoxicity ranging from 4.73% (24 h, 0.1 μg mL–1) to 8.59% (24 h, 0.4 μg mL–1).
Enzyme activities of Caco-2 cells: Enzyme activities (GST, SOD, CAT) of Caco-2 cells are shown in Table 2. Overall, enzyme activities for cells incubated for 12 h were generally higher than corresponding activities at 24 h.
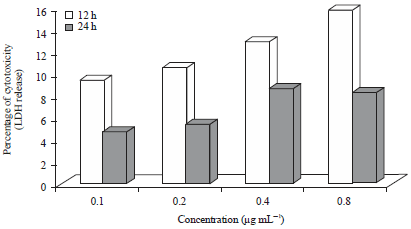 | |
| Fig. 3: | Percentage of cytotoxicity of curry leaf treated HepG2 cells, LDH: Lactate dehydrogenase |
| Table 2: | Enzyme activities of Caco-2 cells treated with curry leaf extracts |
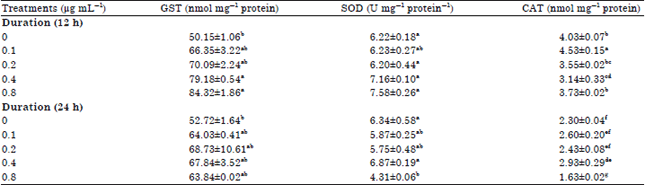 | |
| Values are Means±SEM, Values in a column with different letters are significantly different using Tukey’s studentized range test at p≤0.05, CAT: Catalase, GST: Glutathione-s-transferase, SOD: Superoxide dismutase | |
After 12 h of incubation, both GST and SOD activity increased with increasing concentration. The GST activity ranged from a low of 66.35 (0.1 μg mL–1) to a high of 84.32 nmol mg–1 protein (0.8 μg mL–1), which was approximately 68% higher than untreated Caco-2 cells. The SOD activity ranged from a low of 6.20 (0.2 μg mL–1) to a high of 7.58 (0.8 μg mL–1), which was 21.9% higher than SOD activity of untreated cells. For catalase, enzyme activity was significantly higher at 0.1 μg mL–1 after 12 h at 4.53 nmol mg–1 protein compared to all other treatment concentrations and time periods.
Enzyme activities of HepG2 cells: Enzyme activities of (GST, SOD, CAT) for CLE-treated Hep2G cells are displayed in Table 3. There were no significant differences between GST activities in varying treatment groups. However, catalase activity at 0.1 μg mL–1 24 h (1.38 nmol mg–1 protein) was significantly higher than all other treatments. Catalase activity was inversely proportional to the concentration after 24 h of incubation. For SOD, enzyme activity generally increases after 12 h of incubation, peaking at 3.27 U mg–1 protein (0.8 μg mL–1) but decreased with increasing concentration.
Histone-related DNA fragmentation: Histone-related DNA fragmentation of CLE treated Caco2-treated cells is shown in Fig. 4. Overall, the enrichment factors of cells incubated for 12 h were higher than respective values at 24 h for all treatment except 0.8 µg mL–1.
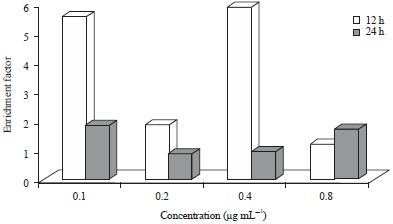 | |
| Fig. 4: | Histone-related DNA fragmentation of curry leaf-treated Caco-2 cells |
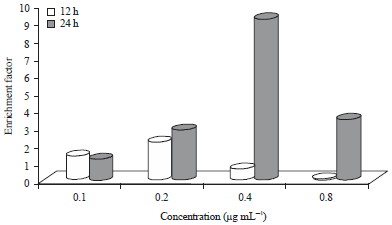 | |
| Fig. 5: | Histone-related DNA fragmentation of curry leaf-treated HepG2 cells |
| Table 3: | Enzyme activities of HepG2 cells treated with curry leaf extracts |
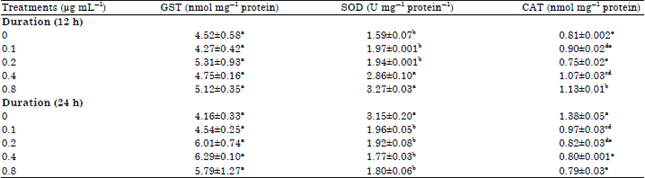 | |
| Values are Means±SEM, Values in a column with different letters are significantly different using Tukey’s studentized range test at p≤0.05, CAT: Catalase, GST: Glutathione-s-transferase, SOD: Superoxide dismutase | |
Enrichments factors at 12 h were 3, 2 and 6 fold greater than values at 24 h for 0.1, 0.2 and 0.4 μg mL–1, respectively.
Histone-related DNA fragmentation of CL-treated HepG2 cell is shown in Fig. 5. Enrichment factors were higher after 24 h incubations compared to respective 12 h values for all treatments except 0.1 μg mL–1. Enrichment factors ranged from a low of 0.88 (0.2 μg mL–1 24 h) to a high of 9.37 (0.4 μg mL–1 24 h).
DISCUSSION
Curry leaf has been shown to display hepatoprotective properties in in vitro studies and chemo-modulatory properties using in vivo (rat-model) studies. This study evaluated the chemopreventive effects of curry leaf extracts in a human colon cancer cell line Caco-2 and determined if there were possible toxicological implications at the respective treatment doses using a human liver cell line Hep2G.
Polyphenols have been reported to exhibit anti-tumor and anticarcinogenic activities (Kandaswami et al., 2005; Thomasset et al., 2007). Recent studies have shown curry leaf to be a good source of polyphenols (Almey et al., 2010; Reihani and Azhar, 2012; Noolu et al., 2013). In our study, the total phenolic content of methanolic extract of curry leaf was 541.11 mg GAE/100 g CL (2.7 mg GAE mL–1). Noolu et al. (2013) reported a similar phenolic content at 3.0 μg GAE μL–1 CLE. Other studies reported total phenolic content of curry leaf to ranges from 20-35 mg GAE g–1 of curry leaf (Almey et al., 2010; Reihani and Azhar, 2012).
The DPPH (0.5 mM) scavenging ability was 90% at a concentration of 250 mg mL–1. Similar results were reported by Andallu et al. (2011). Biswas et al. (2012) reports an ethanolic extract of curry leaf (25 mg mL–1) having a DPPH scavenging activity of approximately 58%. Though the present study uses 0.5 mM of DPPH, displaying a 30% DPPH activity at 25 mg mL–1, a dose-response relationship is clearly shown in the free-radical scavenging ability of curry leaf extract.
Lactase dehydrogenase (LDH) release was used to determine cytotoxicity of extracts in Caco-2 and Hep2G cells. The LDH is an enzyme present in most cells and is released from the cytosol of damaged cells (Verghese et al., 2006). After 24 h of incubation, a positive correlation between cytotoxicity and concentration was seen in Caco-2 cells. A similar trend was seen in Hep2G cells after 12 h of incubation. The CLE exhibited the greatest effect in colon cancer cells (Caco-2) after 24 h. However, LDH was higher with acute exposure to CLE for Hep2G cells. These finding imply that long term exposure exhibits an anti-cancer effect in Caco-2 cells while having a low toxic effect. Similar anticarcinogenic effects were seen in animal model studies. Khanum et al. (2000) reported a 50% reduction in micronuclei in tissues of dimethylhyrdrazine treated rats fed curry leaf. Furthermore, Dasgupta et al. (2003) reported a decrease in LDH release in rats supplemented with curry leaf extract on benzo (a) pyrene induced papillomagenesis.
In the present study, LDH release in HepG2 cells were under 20%, indicating low cytotoxicity. A study investigating the hepatoprotective effects of curry leaf in HepG2, curry leaf extract also showed little effects on the viability of cells via the MTT assay (Sathaye et al., 2011). Moreover, the same study reported a decrease in ethanol-induced toxicity in cells treated with various portions of aqueous curry leaf extract.
After 24 h of incubation of HepG2, SOD and CAT displayed a positive correlation (0.99; p<0.001). This trend is probably due to the SOD and CAT working together in exhibiting its antioxidative properties. The SOD converts O2 to H2O2 (Fridovich, 1975) followed by the decomposition of H2O2 to H2O by catalase (Fridovich, 1975; Bansal et al., 1996). Sathaye et al. (2011) reported a similar simultaneous rise in SOD and catalase activity in Hep2G cells in a study examining the hepatoprotective properties of curry leaf against ethanol-induced toxicity.
Treatment at the highest concentration (0.8 μg mL–1) for 12 h caused a 21% increase in SOD activity. Andallu et al. (2011) reported a similar increase (19.5%) in SOD activity in the erythrocytes of patients receiving 5 g of curry leaf per day. After 12 h, SOD and GST activity increased dose-dependently. An induction of SOD indicates the presence of superoxide radicals (O‾2). The SOD catalyzes the conversion of superoxide radicals to hydrogen peroxide and oxygen (Fridovich, 1975). Normally, catalase supports SOD by further converting hydrogen peroxide to water and oxygen, thus preventing oxidative damage. However, in this study, after 12 h incubation there is a slight decrease in catalase for curry leaf treated Caco-2 cell. Therefore, increased H2O2 production leading to DNA damage may be a possible mechanism of cell damage and death (Cooke et al., 2003; Driessens et al., 2009).
Internucleosomal degradation of genomic DNA is a characteristic of apoptotic cell death (Dhanasekaran and Ganapathy, 2011). Our results indicate that DNA fragmentation was highest at 0.4 μg mL–1 for both Caco-2 and HepG2 cells. Morphological changes of cells at this treatment concentration include the formation of apoptotic bodies. Other morphological changes characteristic of apoptosis seen include zeiosis and condensation of chromatin. Studies have reported curry leaf to have an apoptotic effect on other human cancer cell lines including leukemia (Roy et al., 2004; Ito et al., 2006), breast Noolu et al. (2013) and prostate cells (Roy et al., 2004). Noolu et al. (2013) detected both early and last stage apoptotic cell (via Annexin V-FITC binding) after treatment with CLE in two different human breast cancer cell lines. The same study concluded that apoptotic activity was due to the ability of CLE to inhibit 20 and 26 S proteasome enzymes, which promotes tumor cell proliferation and protect cancer cells from apoptosis (Goldberg, 1995; Hochstrasser, 1995; Groll et al., 1997).
CONCLUSION
The results from this study indicate that curry leaves may be effective in colon cancer prevention while causing minimal toxic effects to liver cells. Morphological changes and Histone-related DNA fragmentation suggest apoptosis as a mechanism of death for Caco-2 cells.
Anti-cancer effects may be due to high phenolic and flavonoid content, as well as free-radical scavenging ability.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Almey, A.A.A., C.A.J. Khan, I.S. Zahir, K.M. Suleiman, M.R. Aisyah and K.K. Rahim, 2010. Total phenolic content and primary antioxidant activity of methanolic and ethanolic extracts of aromatic plants leaves. Int. Food Res. J., 17: 1077-1084.
Direct Link - Andallu, B., M. Shankaran, R. Ullagaddi and V.K. Allagadda, 2011. Efficacy of curry (Murraya koenigii) leaves in scavenging free radicals in vitro and controlling oxidative stress in vivo. Biomed. Preventive Nutr., 1: 263-267.
Direct Link - Bansal, A.K., D. Bhatnagar and G.L. Soni, 1996. In vitro effect of n-nitrosodiethylamine on lipid peroxidation and antioxidant system in human erythrocytes. Toxicol. In vitro, 10: 649-653.
CrossRefPubMedDirect Link - Benzie, I.F.F., Y.T. Szeto, J.J. Strain and B. Tomlinson, 1999. Consumption of green tea causes rapid increase in plasma antioxidant power in humans. Nutr. Cancer, 34: 83-87.
CrossRefDirect Link - Biswas, A.K., M.K. Chatli and J. Sahoo, 2012. Antioxidant potential of curry (Murraya koenigii L.) and mint (Mentha spicata) leaf extracts and their effect on colour and oxidative stability of raw ground pork meat during refrigeration storage. Food Chem., 133: 467-472.
CrossRefDirect Link - Cooke, M.S., M.D. Evans, M. Dizdaroglu and J. Lunec, 2003. Oxidative DNA damage: Mechanisms, mutation and disease. FASEB J., 17: 1195-1214.
CrossRefPubMedDirect Link - Dasgupta, T., A.R. Rao and P.K. Yadava, 2003. Chemomodulatory action of curry leaf (Murraya koenigii) extract on hepatic and extrahepatic xenobiotic metabolising enzymes, antioxidant levels, lipid peroxidation, skin and forestomach papillomagenesis. Nutr. Res., 23: 1427-1446.
CrossRefDirect Link - Dhanasekaran, J.J. and M. Ganapathy, 2011. Hepatoprotective effect of Cassia auriculata L. leaf extract on carbon tetrachloride intoxicated liver damage in wister albino rats. Asian J. Biochem., 6: 104-112.
CrossRefDirect Link - Driessens, N., S. Versteyhe, C. Ghaddhab, A. Burniat and X. de Deken et al., 2009. Hydrogen peroxide induces DNA single-and double-strand breaks in thyroid cells and is therefore a potential mutagen for this organ. Endocrine-Related Cancer, 16: 845-856.
CrossRefDirect Link - Goldberg, A.L., 1995. Functions of the proteasome: The lysis at the end of the tunnel. Science, 268: 522-523.
CrossRefPubMedDirect Link - Groll, M., L. Ditzel, J. Lowe, D. Stock, M. Bochtler, H.D. Bartunik and R. Huber, 1997. Structure of 20S proteasome from yeast at 2.4A resolution. Nature, 386: 463-471.
CrossRefDirect Link - Gupta, P., A. Nahata and V.K. Dixit, 2011. An update on Murraya koenigii spreng: A multifunctional Ayurvedic herb. J. Chin. Integr. Med., 9: 824-833.
CrossRefPubMedDirect Link - Gupta, S., M. George, M. Singhal, G.N. Sharma and V. Garg, 2010. Leaves extract of murraya koenigii linn for anti-inflammatory and analgesic activity in animal models. J. Adv. Pharm. Technol. Res., 1: 68-77.
PubMedDirect Link - Gupta, S. and J. Prakash, 2009. Studies on Indian green leafy vegetables for their antioxidant activity. Plant Foods Hum. Nutr., 64: 39-45.
CrossRefDirect Link - Hochstrasser, M., 1995. Ubiquitin, proteasomes and the regulation of intracellular protein degradation. Curr. Opin. Cell Biol., 7: 215-223.
CrossRefDirect Link - Ito, C., M. Itoigawa, K. Nakao, T. Murata, M. Tsuboi, N. Kaneda and H. Furukawa, 2006. Induction of apoptosis by carbazole alkaloids isolated from Murraya koenigii. Phytomedicine, 13: 359-365.
CrossRefDirect Link - Johnson, J.J. and H. Mukhtar, 2007. Curcumin for chemoprevention of colon cancer. Cancer Lett., 255: 170-181.
CrossRefDirect Link - Kandaswami, C., L.T. Lee, P.P. Lee, J.J. Hwang, F.C. Ke, Y.T. Huang and M.T. Lee, 2005. The antitumor activities of flavonoids. In vivo, 19: 895-909.
PubMedDirect Link - Khanum, F., K.R. Anilakumar, K.S. Krishna, K.R. Viswanathan and K. Santhanam, 2000. Anticarcinogenic effects of curry leaves in dimethylhydrazine-treated rats. Plant Foods Hum. Nutr., 55: 347-355.
CrossRefDirect Link - Kim, D.O., O.K. Chun, Y.J. Kim, H.Y. Moon and C.Y. Lee, 2003. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem., 51: 6509-6515.
CrossRefDirect Link - Mani, V., K. Ramasamy, A. Ahmad, M. Parle, S.A.A. Shah and A.B.A. Majeed, 2012. Protective effects of total alkaloidal extract from Murraya koenigii leaves on experimentally induced dementia. Food Chem. Toxicol., 50: 1036-1044.
CrossRefDirect Link - Manzano, A. and P. Perez-Segura, 2012. Colorectal cancer chemoprevention: Is this the future of colorectal cancer prevention? Sci. World J., Vol. 2012.
CrossRef - Mohandas, K.M. and D.C. Desai, 1998. Epidemiology of digestive tract cancers in India. V. Large and small bowel. Indian J. Gastroenterol.: Official J. Indian Soc. Gastroenterol., 18: 118-121.
PubMedDirect Link - Yoshimasa, N., H. Oshigashi, S. Masuda, M. Akira and M. Yasujiro et al., 2000. Redox regulation of glutathione S-transferase induction by benzyl isothiocyanate: Correlation of enzyme induction with the formation of reactive oxygen intermediates. Cancer Res., 60: 219-225.
PubMedDirect Link - Ningappa, M.B., B.L. Dhananjaya, R. Dinesha, R. Harsha and L. Srinivas, 2010. Potent antibacterial property of APC protein from curry leaves (Murraya koenigii L.). Food Chem., 118: 747-750.
CrossRefDirect Link - Ningappa, M.B., R. Dinesha and L. Srinivas, 2008. Antioxidant and free radical scavenging activities of polyphenol-enriched curry leaf (Murraya koenigii L.) extracts. Food Chem., 106: 720-728.
CrossRefDirect Link - Noolu, B., R. Ajumeera, A. Chauhan, B. Nagalla, R. Manchala and A. Ismail, 2013. Murraya koenigii leaf extract inhibits proteasome activity and induces cell death in breast cancer cells. BMC Complementary Alternat. Med., Vol. 13.
CrossRef - Palaniswamy, U.R., 2001. Human dietetics and Asian food crops. HortTechnology, 11: 504-509.
Direct Link - Reihani, S.F.S. and M.E. Azhar, 2012. Antioxidant activity and total phenolic content in aqueous extracts of selected traditional Malay salads (Ulam). Int. Food Res. J., 19: 1439-1444.
Direct Link - Roy, M.K., V.N. Thalang, G. Trakoontivakorn and K. Nakahara, 2004. Mechanism of mahanine-induced apoptosis in human leukemia cells (HL-60). Biochem. Pharmacol., 67: 41-51.
CrossRefDirect Link - Sathaye, S., Y. Bagul, S. Gupta, H. Kaur and R. Redkar, 2011. Hepatoprotective effects of aqueous leaf extract and crude isolates of Murraya koenigii against in vitro ethanol-induced hepatotoxicity model. Exp. Toxicol. Pathol., 63: 587-591.
CrossRefDirect Link - Singh, A.P., T. Wilson, D. Luthria, M.R. Freeman and R.M. Scott et al., 2011. LC-MS-MS characterisation of curry leaf flavonols and antioxidant activity. Food Chem., 127: 80-85.
CrossRefDirect Link - Singleton, V.L., R. Orthofer and R.M. Lamuela-Raventos, 1999. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In: Methods in Enzymology, Burslem, G.L. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121822002, pp: 152-178.
CrossRefDirect Link - Tajkarimi, M.M., S.A. Ibrahim and D.O. Cliver, 2010. Antimicrobial herb and spice compounds in food. Food Control, 21: 1199-1218.
CrossRefDirect Link - Thomasset, S.C., D.P. Berry, G. Garcea, T. Marczylo, W.P. Steward and A.J. Gescher, 2007. Dietary polyphenolic phytochemicals-promising cancer chemopreventive agents in humans? A review of their clinical properties. Int. J. Cancer, 120: 451-458.
CrossRefDirect Link - Verghese, M., D.R. Rao, C.B. Chawan, L.T. Walker and L. Shackelford, 2006. Anticarcinogenic effect of phytic acid (IP6): Apoptosis as a possible mechanism of action. LWT-Food Sci. Technol., 39: 1093-1098.
CrossRefDirect Link - Xie, J.T., W.T. Chang, C.Z. Wang, S.R. Mehendale and J. Li et al., 2006. Curry leaf (Murraya koenigii Spreng.) reduces blood cholesterol and glucose levels in ob/ob mice. Am. J. Chin. Med., 34: 279-284.
CrossRefPubMedDirect Link