
K. Busambwa
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
M. Verghse
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
R.M. Cebert
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
L. Dalrymple
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
J. Allen
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
J. Boateng
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
L. Shackelford
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
L.T. Walker
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agriculture and Mechanical University, Normal, Alabama, 35762, USA
Journal of Pharmacology and Toxicology
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 36-48







ABSTRACT
Lentils, green and yellow split-peas have been reported to provide health benefits against colon cancer due to the amount of nutrients and non-nutrient phytochemical compounds present. The aim of this study was to investigate and compare the chemopreventive potential of sprouted and non-sprouted lentils (LS, LNS), green (GS, GNS) and yellow split-peas (YS, YNS) on azoxymethane (AOM)-induced colon cancer. Following a 1 week acclimatization period, 42 Fisher-344 male rats were randomly assigned to 6 groups (n = 6). Five groups were fed treatment diets consisting of the selected legumes (Sprouted and non-sprouted), while the control group (C) was fed AIN-93 growth and maintenance of diet. Colon tumors were induced by administration of AOM at 7 and 8 weeks of age. Rats were killed by CO2 asphyxiation at age 46 weeks. Results showed lower tumor incidence in treatment groups at 66.7% in GS compared to 100% in LNS and the control. Rats fed control diet had higher Tumors/Tumor Bearing Rat (TBR) ratio (4.33) compared to those in treatment groups (1.2-2). Cecal pH was significantly higher in control (7.81) compared to the treatment diets. Glutathione-S-Transferase (GST) activity was significantly higher in sprouted legumes (8.55-14.04 μM min–1 mL–1) compared to non-sprouted legumes (4.53-5.67 μM min–1 mL–1). Glutathione concentration (GSH) ranged from a low of 636.34 μM in rats fed GNS to a high of 791.07 μM in rats fed YNS. Selected legumes were effective in reducing incidence of AOM-induced colon tumors in Fisher-344 male rats (2.1-4.3 times) and may be promoted for consumption as part of healthy eating habits to prevent chronic diseases such as cancer.
PDF Abstract XML References Citation
Received: July 10, 2015;
Accepted: September 19, 2015;
Published: October 12, 2015
How to cite this article
K. Busambwa, M. Verghse, R.M. Cebert, L. Dalrymple, J. Allen, J. Boateng, L. Shackelford and L.T. Walker, 2015. Lentils, Green and Yellow Split-Peas (Sprouted and Non-Sprouted) on Azoxymethane-Induced Colon Carcinogenesis. Journal of Pharmacology and Toxicology, 10: 36-48.
DOI: 10.3923/jpt.2015.36.48
URL: https://scialert.net/abstract/?doi=jpt.2015.36.48
DOI: 10.3923/jpt.2015.36.48
URL: https://scialert.net/abstract/?doi=jpt.2015.36.48
INTRODUCTION
Colorectal cancer is a result of accumulation of genetic changes leading to development of neoplasia and affects mostly the colon and the rectum. Western world have seen high incidence of colorectal cancer and high mortality in the past decade. However, the number of colon cancer cases have decreased due to mostly improved treatment, increased awareness and early detection of the disease (Jemal et al., 2011). Diet, genetic and environmental factors are believed to influence the development of colorectal disease in the western countries. Also, some factors such as family history of adenomas, genetic polyposis and non-polyposis syndromes, inflammatory bowel disorders can play a role in increasing the risk of developing (Rustgi, 2007; Jasperson et al., 2010). Individual with advanced neoplasia have high risk of developing colorectal cancer and this is mostly observed in aged population (70 year old persons) (Popescu et al., 1999).
Based on pathways that regulate some aspects in the development of colon cancer, colon cancer can take two different forms. Familial polyposis results from mutated Adenomatous Polyp Coli (APC) gene (Augenlicht et al., 2002). This form of colon cancer is characterized by formation of many begin tumors (100-1000) in the colon by the age of 30 years which can evolve into cancer if not treated (Augenlicht et al., 2002; Rustgi, 2007). In contrast, Hereditary Non-polyposis Colorectal Cancer (HNPCC) results from mis-match repair gene mutations (Rodriguez-Bigas et al., 1997). The HNPCC is the most form of hereditary colon cancer and account for 15% of all colon cancer (Umar et al., 2004). This form of colon cancer appears earlier in life compared to the sporadic colorectal cancer observed frequently in individuals above 65 years (Baba, 1997). However, according to American Cancer Society, links between diet and colorectal cancer risk are some of the strongest for any type of cancer. Therefore, studies have focused on diet as an alternative method in the prevention and/or therapy of this type of cancer (Bingham and Riboli, 2004; Willett, 2001). The US consumption of legumes such as peas, lentils have increased in the past decade due to the medical recommendation to increase the dietary intakes of fiber, protein, folate, zinc, iron and magnesium found in these legumes to lower intakes of saturated fat responsible for chronic diseases such as diabetes, CVD and cancer (Mitchell et al., 2009). Legumes are high in fiber and low in fat, they are good source of macro-nutrients such as calcium, magnesium, potassium and iron (Longnecker, 2000). Lentils and peas are good source of trace elements such as copper, zinc, selenium and manganese; they are high in B group vitamins, thiamine, niacin and folate. Also, these legumes contains phytonutrients with antioxidantive properties (Longnecker, 2000). Sprouting of grains affects dramatically vitamins A and C and phytochemical levels and increases bioavailability of macro and micro-nutrients (Hedges and Lister, 2006).
Lentils (Lens culinaris) have been used as art of diet in Asian culture for many years. They are especially good source of dietary fiber and have been use in the control of diabetes due to their ability to improve digestibility and postprandial glycemia (Jenkins et al., 1982). Dietary fiber may affect colonic and fecal pH and also prevents the induction of procarcinogenic enzymes such as nitroreductase, azoreductase, β-glucorunidase and 7-α-dehydroxylase. The enzyme 7-α-dehydroxylase plays a role in the conversion of primary bile acids into procarcinogenic secondary bile acids. Lentils are also good source of folate which play a important role in DNA synthesis, preventing the process of colon carcinogenesis at initiation and progression stage (Kim et al., 2009). Peas, the seeds of Pisum sativum primarily produced in tropical regions but widely consumed because they are good source of dietary proteins and energy (Grant et al., 2003). Peas are source of bioactive compounds such as phytochemicals, micro-nutrients and macronutrients. Phytochemicals found in these legumes are generally polyphenols such as phenolics compounds and flavonoids have been shown to have anti-inflammatory and anti-oxidative properties (McCrory et al., 2010). Therefore, lentils, green and yellow split peas may provide health benefits against colon cancer. The objective of this study was to investigate and compare the chemopreventive potential of sprouted and non-sprouted lentils (LS, LNS), green (GS, GNS) and yellow split-peas (YS, YNS) on azoxymethane (AOM)-induced colon cancer.
MATERIALS AND METHODS
Animal housing and experimental design: After 7 days of acclimatization, 42 Fischer-344 male weanling rats (4 weeks old) obtained from Harlan, IN were randomly divided into 7 groups (6 rats per groups) and assigned to one of the following diet for 46 weeks: AIN-93G (Reeves, 1997) (Control); lentils, green and yellow split-peas (sprouted and non-sprouted) at 10% (Fig. 1).
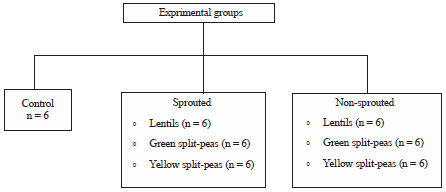 | |
| Fig. 1: | Experimental design total number of rats (N) = 42 |
Light and dark cycles were held at 12 h each. Temperature and relative humidity were maintained at 21°C and 50%, respectively. Rats consumed feed and water ad libidium and weekly body weight and daily feed intakes were recorded. Processed flours of selected legumes were obtained from US Beans council (CA) and prepared diet was stored at 4°C. All the protocols involving rats have been approved by the Institutional Animal Care and Use Committee of Alabama Agricultural and Mechanical University, Normal, Alabama.
Carcinogen injection: At 7 and 8 weeks of age, all animals received 2 subcutaneous injections of azoxymethane (Sigma Chemicals, St Louis, MO) in saline solution at 16 mg kg–1 b.wt.
Organ samples collection and counting of tumors: At the end of 46 weeks experiment, rats were sacrificed by CO2 asphyxiation. Colons, livers and cecum were removed. Characterization of colon tumors were determined as described by protocols established by previous researchers (Shackelford et al., 1983). The number, size/volume, location of tumors and Tumors/Tumor Bearing Rate (TBR) ratio were recorded following the protocol described by Verghese et al. (2002).
Cecal weight and cecal pH: The cecum from rats was removed, weighed and the pH of cecal contents was recorded.
Preparation of liver samples for hepatic antioxidant and detoxification enzyme activities: Approximately 5 g of liver sample was homogenized in 50 mL of potassium phosphate buffer (0.1 m, pH 7.2) and centrifuged at 15,0000×g for 20 min. Supernatant was collected for antioxidant enzyme, catalase and for hepatic detoxification enzyme, glutathione-s-transferase assays. For determination of glutathione hepatic content, the supernatant was deproteinated, then stored at -20°C.
Catalase assay: The method is based on the reaction of the enzyme with methanol in the presence of H2O2 as described by Johansson and Borg (1988). One unit of CAT was defined as the amount of enzyme needed to cause the formation of 1 nmol of formaldehyde per minute. The absorbance was recorded using microplate reader (synergy HT) at 540 nm.
Glutathione-s-transferase assay: Liver supernatant was mixed with 1-chloro 2, 4-dinitrobenzene (CDNB) (Fisher Scientific, Suwanne, GA). Hepatic GST activity in animal liver was assayed following the protocol described by Habig et al. (1974). The GST was determined at 340 nm and expressed as micrometer per milligram.
Hepatic glutathione content: Using 5, 5’-dithio-bis-(2-nitrobenzoic acid) as Ellman’s reagent (Fisher Scientific, Suwannee, GA), GSH was determined at 405-414 nm and expressed as micromole. The GSH serves as a nucleophilic substrate in the detoxification of xenobiotics. The GSH content was determined following methods as described by Griffith (1980).
Preparation of extracts from sprouted and non-sprouted lentils, green split-pea and yellow peas for total phenolics, total flavonoids and FRAP assays: Lentils, green and yellow split-peas (sprouted and non-sprouted) extracts were prepared based on the method described by Adom and Liu (2002) with slight modifications. Briefly, 5 g of selected legumes was extracted with 50 mL of 80% ethanol (v/v) for 2 h at room temperature. The samples were centrifuged at 3000×g for 20 min and the supernatant was collected.
Determination of total phenolics in selected legumes (sprouted and non-sprouted), sprouted and non-sprouted lentils, green and yellow split peas was analyzed following the Folin-Ciocalteau method (Adom and Liu, 2002; Singleton et al., 1999). Briefly, 12.5 μL of appropriately diluted sample extracts was oxidized with 12.5 μL of Folin-Ciocalteau reagent. After a 5 min incubation period the reaction was neutralized with 50 μL of distilled water and with 125 μL of 7% sodium carbonate solution (Na2CO3) (v/v). Gallic acid was used as standard solution (0.01 mg100 mL). After 2 h incubation at room temperature a resulting blue color was read at an absorbance of 750 nm. Total phenolics content in sprouted and non-sprouted lentils, green and yellow split peas was expressed as milligram Gallic Acid Equivalent (GAE)/g.
Determination of total flavonoids content in selected legumes (sprouted and non-sprouted) total flavonoid content in extracts was analyzed by a colorimetric method described by Adom and Liu (2002) and Kim et al. (2003). Briefly 7.5 μL of 5% sodium nitrite (NaNO2) were added to 25 μL of appropriately diluted samples of each selected product extract (1:20). The mixture was allowed to incubate for 5 min after which 15 μL of 10% aluminum chloride (AlCl3) was added. The mixture was further incubated for 5 min and 1 mL of 1 M sodium hydroxide (NaOH) was added. Another 40 μL of distilled water were immediately added to the resulting pink mixture. The absorbance was immediately read at 510 nm. The flavonoid content in the selected food extracts was compared to that of catechin standards. The total flavonoid content is reported as milligram Catechin Equivalent (CE)/g.
Determination of Ferric Reducing Antioxidant Power (FRAP) activity of selected legumes: The FRAP assay measures the change in color caused by the reduction of ferric complex (Fe3+) to ferrous form (Fe2+) by the action of electron donating antioxidants. The FRAP assay was performed based on the methods of Benzie and Strain (1999). Briefly, 10 μL of appropriately diluted samples of each selected legume extract were added to 30 μL of working FRAP reagent consisting of 300 mM acetate buffer pH 3.6, 10 mM 2,4,6-tri (2-pyridyl)-s-triazine (TPTZ) in 40 mM HCL and 20 mM ferric chloride (FeCl3.6H2O). The absorbance was recorded at 593 nm for 4 min at 1 min of interval. The absorbance changes in the test mixture were compared to those obtained from standard mixture of ferrous sulphate (FeSO4.7H20) (0.1-1.0 mM). The FRAP is expressed as micromole of Fe2+/g.
Statistical analysis: Data are expressed as Means±SEM and were analyzed using the SAS system version 9.2 (SAS Institute, Cary, NC). Differences were considered significant at p<0.05. Means are separated and analyzed by ANOVA using Tukey’ studentized test.
RESULTS
Weight gain, feed intake, cecal weight and cecal pH in rats fed selected legumes: There were no significant differences (p<0.05) in feed intake among rats fed control and treatment diets (Table 1). The feed intake of rats fed control diet was similar to that of LS, LNS and YS. Rats fed GS and GNS diet had lower feed intake compared to control and other treatments diets. Consumption of non sprouted legumes was slightly higher compared to sprouted legumes except for lentils. Highest weight gain Table 1 was seen in rats fed YS diet while the lowest weight gain was observed in LNS fed rats. Cecal weight ranged from a low of 1.40 (YS) to a high of 2.15 (LNS). Among the treatments groups, rats fed LNS diet (7.6) had the highest cecal pH compared to its counterparts.
Total phenolics, total flavonoids content and antioxidant capabilities using FRAP: Total flavonoid content in extracts was determined by a colorimetric method using catechin as standard solution. Total flavonoids content ranged from a low of 0.53 (GNS) to a high of 1.78 (LS) (Fig. 2). There were no significant differences among the sprouted and non-sprouted legumes except for green split-peas. Sprouted legumes had higher total flavonoid content compared to non-sprouted legumes while the lowest total flavonoid content was seen in green split-peas sprouted (0.74) and non-sprouted (0.53). However, total phenolics content ranged from a low of 1.98 (GNS) to a high of 4.45 (LS). There were significant differences (p<0.05) in total flavonoids contents in lentils sprouted and non-sprouted compared to other legumes.
| Table 1: | Weight gain, feed intake, cecal weight and cecal pH in Fisher-344 male rats |
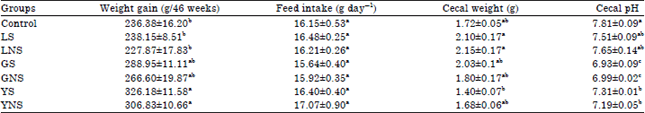 | |
| abcMeans in column with the same superscript do not significantly differ (p<0.05) using Tukey’s studentized test. Values are Means±SE, LS: Sprouted lentil, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non-sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted | |
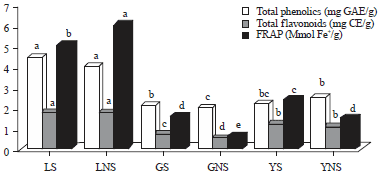 | |
| Fig. 2: | Total phelonic content, total flavonoids content and antioxidant activity using FRAP values are Means±SE and Bars with the same superscript do not significantly differ (p<0.05) Tukey’s tudentized test, LS: Sprouted lentil, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted |
| Table 2: | Effects of diets on catalase and glutathione-s-transferase activities and glutathione concentrations in Fisher-344 male rats |
 | |
| abcMeans in column with the same superscript do not significantly differ (p<0.05) using Tukey’s studentized test. Values are Means±SE, LS: Sprouted lentil, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non-sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted, CAT: Catalase, GST: Glutathione-s-transferase, GSH: Glutathione | |
Sprouted legumes had higher total flavonoid (Fig. 2) content compared to non-sprouted legumes, except in yellow peas. Antioxidant capabilities measured by FRAP in selected legumes ranged from a low 0.65 in sprouted green split-peas to a high of 6.04 (Fig. 2) in non-sprouted lentils. Out of the selected legumes, sprouted and non-sprouted lentils had high total phenolics and flavonoids content and high antioxidant activity compared to green and yellow split-peas while green split-peas (sprouted and non-sprouted) exhibit the lowest total phenolics and flavonoids content and antioxidant activity.
Catalase, glutathione-s-transferase assay and glutathione content determination: Catalase activity was significantly higher (p<0.05) in rats fed control, LS, LNS, GS and GNS diets compared to rats fed YS and YNS diets (Table 2). Out of lentils and green split-peas diets, non-sprouted legumes had higher catalase activity compared to sprouted legumes. The GST activity was significantly higher in rats fed GS (14.04) compared to other treatments and control diets. However, the lowest GST activity was seen in rats fed GNS (4.53). Once again, GST activity was significantly higher in rats fed sprouted legumes compared to its counterparts. The GSH content ranged from a low of 636.34 (GNS) to a high of 791.07 (YNS) (Table 2). Rats fed YNS had higher GSH content while rats fed GNS had shown the lowest GSH content compared to other treatment and control diets. The GSH content was significantly higher in rats fed sprouted legumes compared to rats fed non-sprouted legumes, except in yellow peas diet groups.
Characteristics of colon tumors
Tumor number and size: Regardless of the diets, higher numbers of tumors were primarily observed in the distal colon than in proximal portion of the colon. However, total tumor numbers in rats fed control diet were higher compared to the treatment diets (Table 3). Among the treatments diet, total tumor numbers ranges from a low of 6 in GS and YS diets to a high of 12 in LNS diet. Higher total tumor numbers were observed in rats fed non-sprouted legumes compared to those fed sprouted legumes. Larger tumors (mm) were observed in rats fed control diet (4.92) compared to those fed treatment diets. Meanwhile, rats fed GS diet had shown the smallest tumor (0.91 mm) compared to rats fed other treatments diets. Except in lentils diet groups, rats fed sprouted diets had smaller tumors compared to rats fed non-sprouted diets.
Tumor incidence and Tumor/Tumor Bearing Rat (TBR): Total tumor incidence (%) ranged from a low of 66.7 in GS diet to a high of 100 in control and LNS diets (Table 3). Out of the treatment diets, the highest total incidence was seen in LNS diet and there were no differences in total incidence between yellow peas sprouted and non-sprouted. Among the treatment groups, LNS diet had higher T/TBR (2) while the lowest T/TBR was observed in rats fed YS diet (1.2).
| Table 3: | Tumor number, size, location, tumor incidence and tumor/tumor bearing rat in the colon of Fisher-344 male rats fed selected legumes |
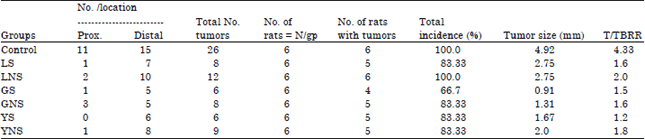 | |
| Values are Means±SE, LS: Sprouted lentil, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non-sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted | |
DISCUSSION
Legumes, due to their most abundant constituents such as dietary fiber and polyphenol, have been studied on animals and cell culture in the prevention of chronic diseases such as diabetes, neurodegenerative and cardiovascular diseases and various cancers (Scalbert et al., 2005). The consumption of these types of food has been encouraged in the USA by the American Cancer Society (ACS), the American Heart Association (AHA) and the American Diabetes Association (ADA) as part of a healthy diet. Lentils as a good source of dietary fiber and enzyme inhibitors such as phytates, tannins, antigenic proteins have been studied in the treatment of diabetes (Jenkins et al., 1982). According to Jenkins et al. (1982), the incorporation of lentils in a diet can help to manage postprandial glycemia in normal and diabetic patients due to enzyme inhibitors ability to decrease the rate of carbohydrate metabolism. Also, peas as source of an array of nutrients and phytochemical have been shown to provide some health benefits. A regular consumption of green and yellow split-peas can result in higher intakes of fiber, protein, folate, zinc, iron and magnesium; important elements in the prevention of chronic diseases (Mitchell et al., 2009). Sprouting of legume seed has been proven to be the best method to decrease antinutritional factors such as phytic acid, tannins and trypsin inhibitors responsible of inactivating proteolytic enzyme trypsin resulting in reduced bioavailability of essential amino acids, vitamins and minerals (Vidal-Valverde et al., 1994). Thus, the lack of these essential nutrients has resulted in some nutrient deficiency conditions. The objectives in this study was to investigate the chemopreventive potential of lentils, green and yellow split-peas, sprouted and non sprouted against azoxymethane induced colon carcinogenesis in Fisher-344 male rats. Results from this study showed that incidence of AOM-induced tumors were significantly reduced in rats fed selected legumes (sprouted and non-sprouted) compared to rats fed the control diet. This supports previous findings suggesting that a diet rich in dietary fiber, essential nutrients and phytonutrients may play a crucial role in regressing or inhibiting colon carcinogenesis.
There were no significantly differences in feed intake; however, rats fed YS and YNS exhibit high degree of efficiency in converting consumed food into body mass. Rats fed yellow split-peas sprouted and non-sprouted had significantly higher weight gain compared to control and other treatment diets. According to Rao et al. (1987), the incidence of tumors is low in rats fed a restricted diet resulting in low body weight gain. This low tumors incidence in rats with low weight gain may be explained by the high protein:fat ratio of the diet and type of carbohydrates in the diet (Rao et al., 1987). Also, Tannenbaum (1942) stated that a calorie-restricted diet plays a role in the tumor inhibition due to the limited nutrients supply necessary for metabolism and growth of normal and transformed tissues. This contradicts our findings, of the groups with low weight gain (Control, LS and LNS), rats fed control diet had the highest tumor incidence. The low weight gain in control group can be explained by the reduction of surface area of nutrient absorption due to high number of tumors present in colon. Cecal pH (7.81) in control was higher compared to the treatment groups and the lowest pH was seen in rats fed GS and GNS. The low cecal pH seen in GS and GNS groups may be the result of fermentable fiber present in green split-peas. Soluble fiber induces competitive exclusion in gut leading to a high acidic colonic environment which is unfavorable for the growth of pathogenic enzymes responsible for synthesis of some procarcinogen compounds. Also, SCFA butyrate generates from the fermentation of dietary fiber, plays a role in the induction of apoptosis of initiated cell and cell differentiation (Heerdt et al., 1994; Hague and Paraskeva, 1995). Moreover, butyrate have been shown to cause histone acetylation and regulates the expression of various oncogenes (Scheppach et al., 1995). Eberharter and Becker (2002) stated that the acetylation and deacetylation of nucleosomal histone is crucial in the regulation and expression of genes during all nuclear processes involving DNA substrates. According to Heerdt et al. (1997), colon carcinogen can also be a result of abnormalities in mitochondrial activities and butyrate plays a role in the inhibition of tumor development by enhancing functions in the mitochondria leading to cell cycle arrest. Also, butyrate plays a role in the proliferation of normal cell and may be the reason why cecal weight is higher in rats fed lentils and green split-peas. Low cecal weight gain seen in rats fed yellow split-peas may be due to low amount of soluble fiber.
Another possible mechanism for the reduction of tumors may be the presence of phytochemicals such as flavonoids and other phenolic compounds in selected legumes with antioxidant properties which may play a crucial role in the protection of cells against free radicals (Rice-Evans et al., 1997; Yao et al., 2004; Yu et al., 2002). According to Oak et al. (2005), these bioactive compounds have been shown to have anticarcinogenic properties due to their ability to inhibit the expression of proangiogenic factors, Matrix Metallo Proteinases (MMPs) and Vascular Endothelial Growth Factor (VEGF) and the formation of new blood vessels. In addition, polyphenols in legumes play an important role as antinflammatory agents through reduction of the activity and expression of NADPH oxidase, xanthine oxidase, lipoxygenase, important enzymes in the inflammation process and thus prevention of the induction and synthesis of proinflammatory elements such as leukotrienes (De Groot and Rauen, 1998; Oak et al., 2005; Sud'ina et al., 1993).
In this study, lentils sprouted and non-sprouted had significantly (p<0.05) higher TPC, TFC with higher FRAP activity compared to green and yellow split-peas sprouted and non-sprouted. There were not significant differences in TPC and TFC between the sprouted and non-sprouted legumes, except in green split-peas groups where GNS had low TPC (1.98) and TCF (0.53) compared to GS (2.11 and 0.74, respectively). Antioxidant activity was higher in non-sprouted lentil (6.04) and lower in non-sprouted green and yellow split-peas compared to its counterparts. Sprouting of seed have been shown to be one of efficient methods for enhancing nutrition value of seeds, their antioxidative activity thus improving the health quality of foods (Perez-Balibrea et al., 2011).
In this study, sprouted and non-sprouted lentils had higher antioxidant activity compared to green and yellow split-peas (sprouted and non-sprouted). Seeds with dark color seed coat such as lentils do exhibit a slightly higher antioxidant activity compare to peas seeds which have a light color coat (Marathe et al., 2011). This contradicts the findings of Lopez-Amoros et al. (2006) which suggested that after sprouting or germination, lentils exhibit a decrease in antioxidant activity by contrast, peas and beans show a significant enhancement of their antioxidant activity. On the other hand, sprouted legumes have shown an increased antioxidant activity compared to non-sprouted legumes, except in the FRAP activity where non-sprouted lentils was higher compared to sprouted lentils (Fig. 2). Our observations are supported by previous findings by suggesting that there is enhanced synthesis of polyphenols and improvement of their antioxidantive properties during seed sprouting (Andarwulan et al., 1999; Cevallos-Casals and Cisneros-Zevallos, 2010; Kim et al., 2012; Yang et al., 2001a). Scientific data have reported the ability of polyphenols founds in legumes to inhibit carcinogenesis at molecular level in the initiation, promotion and progression phase (Yang et al., 2001b). These cell cultures and animal studies support our results in which we saw a significant decrease in tumors incidence in rats fed selected legumes (sprouted and non-sprouted) compared to control.
The induction of phase II enzyme has been suggested to be part of ways to achieve protection against cancer. Metabolism of toxics involves phase I and phase II enzymes. Phase I reactions involving cytochrome P-450 enzymes may occur by reduction and oxidation via the introduction of reactive or polar hydrophilic molecule into xenobiotics and the conjugation of theses modified compounds is promoted by phase II enzyme such as glutathione-s-transferase with endogenous ligands such as glutathione, resulting in more excretal water soluble compounds (Liska, 1998). Phase II step is important and sufficient mechanism of detoxification of many carcinogens, however the balance between phase I and phase II enzymes is crucial to the susceptibility of macromolecules such as DNA, proteins to carcinogen damages (Hodek et al., 2009). Therefore, inducers such as phytochemicals found in legumes, which selectively enhance phase II enzymes without elevated phase I enzymes are preferred to achieve protection against carcinogens (Gerhauser et al., 1997; Talalay and Fahey, 2001). Various phytochemicals from edible plants have been reported to inhibit, reverse or retard a specific stage of the tumorigenic process (Surh, 2003).
In this study, hepatic catalase activity and glutathione content were significantly higher in non-sprouted legumes fed rats compared to sprouted groups, except for glutathione levels in rats fed lentils and green split-peas (Table 2). This can be explained by the presence of sugar binding proteins in the beans such as lectins which is very resistant to digestion, active in the colon and affect the function of colonic cell line (Rhodes, 1999). Dietary lectin has been suggested to inhibit proliferation of colonic cancer cells by inducing cellular differentiation and therefore slows the carcingenesis progression (Madar and Stark, 2002). Another mechanism by which dietary legumes play a role in the chemoprevention is via free radicals scavenging ability of water soluble proteins found in these foods (Okada and Okada, 2000). According to Madar and Stark (2002), these proteins enhance antioxidative capacity in cell lines by increasing catalase activity and glutathione concentrations. Although, catalase, GST activities and glutathione contents were higher in control compared to YS and YNS (for catalase activity), to all the treatment groups except GS (for GST activity) and to GNS fed rats (for GSH content), the total number of tumors in rats fed treatment diets were 2-3 times lower compared to rats fed control diet (Table 3). In this case, macronutrients, micronutrients and others chemicals compounds found in plant foods may have the ability to inhibit tumorigenesis (Surh, 2003).
CONCLUSION
This study provides evidences that sprouted and non-sprouted legumes may be effective in the regression of carcinogenesis. These dietary components have the ability to reduce or prevent further damage to macromolecules (DNA, proteins, lipids), which could result in increased incidence of cancer. Sprouting of seeds improve dramatically the nutrient quality by decreasing antinutritional factors. This experimental data showed that sprouted legumes may offer greater protection against chronic diseases by aiding in the induction of antioxidative and detoxicification enzymes.
ACKNOWLEDGMENT
This study was supported by funding from the Agricultural Experiments Research Station, Alabama Agricultural and Mechanical, University, Normal, 35762 Alabama.
REFERENCES
- Adom, K.K. and R.H. Liu, 2002. Antioxidant activity of grains. J. Agric. Food Chem., 50: 6182-6187.
CrossRefDirect Link - Andarwulan, N., D. Fardiza, G.A. Wattimena and K. Shetty, 1999. Antioxidant activity associated with lipid and phenolic mobilization during seed germination of Pangium edule Reinw. J. Agric. Food Chem., 47: 3158-3163.
CrossRefDirect Link - Augenlicht, L.H., J.M. Mariadason, A. Wilson, D. Arango, W.C. Yang, B.G. Heerdt and A. Velcich, 2002. Short chain fatty acids and colon cancer. J. Nutr., 132: 3804S-3808S.
Direct Link - Baba, S., 1997. Hereditary nonpolyposis colorectal cancer: An update. Dis. Colon Rectum, 40: S86-S95.
CrossRefDirect Link - Benzie, I.F.F. and J.J. Strain, 1999. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Method Enzymol., 299: 15-27.
CrossRefPubMedDirect Link - Bingham, S. and E. Riboli, 2004. Diet and cancer-the European prospective investigation into cancer and nutrition. Nat. Rev. Cancer, 4: 206-215.
CrossRefDirect Link - Cevallos-Casals, B.A. and L. Cisneros-Zevallos, 2010. Impact of germination on phenolic content and antioxidant activity of 13 edible seed species. Food Chem., 119: 1485-1490.
CrossRefDirect Link - De Groot, H. and U. Rauen, 1998. Tissue injury by reactive oxygen species and the protective effects of flavonoids. Fundam. Clin. Pharmacol., 12: 249-255.
CrossRefPubMedDirect Link - Eberharter, A. and P.B. Becker, 2002. Histone acetylation: A switch between repressive and permissive chromatin. EMBO Rep., 3: 224-229.
CrossRefDirect Link - Gerhauser, C., M. You, J. Liu, R.M. Moriarty and M. Hawthorne et al., 1997. Cancer chemopreventive potential of sulforamate, a novel analogue of sulforaphane that induces phase 2 drug-metabolizing enzymes. Cancer Res., 57: 272-278.
PubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Hague, A. and C. Paraskeva, 1995. The short-chain fatty acid butyrate induces apoptosis in colorectal tumour cell lines. Eur. J. Cancer Prev., 4: 359-364.
PubMedDirect Link - Heerdt, B.G., M.A. Houston and L.H. Augenlicht, 1994. Potentiation by specific short-chain fatty acids of differentiation and apoptosis in human colonic carcinoma cell lines. Cancer Res., 54: 3288-3294.
PubMedDirect Link - Heerdt, B.G., M.A. Houston and L.H. Augenlicht, 1997. Short-chain fatty acid-initiated cell cycle arrest and apoptosis of colonic epithelial cells is linked to mitochondrial function. Cell Growth Differ., 8: 523-532.
PubMedDirect Link - Hodek, P., J. Krizkova, K. Burdova, M. Sulc, R. Kizek, J. Hudecek and M. Stiborova, 2009. Chemopreventive compounds-view from the other side. Chemico-Biol. Interact., 180: 1-9.
CrossRefDirect Link - Jasperson, K.W., T.M. Tuohy, D.W. Neklason and R.W. Burt, 2010. Hereditary and familial colon cancer. Gastroenterology, 138: 2044-2058.
CrossRefDirect Link - Jemal, A., F. Bray, M.M. Center, J. Ferlay, E. Ward and D. Forman, 2011. Global cancer statistics. CA: Cancer J. Clin., 61: 69-90.
CrossRefPubMedDirect Link - Jenkins, D.J., M.J. Thorne, K. Camelon, A. Jenkins and A.V. Rao et al., 1982. Effect of processing on digestibility and the blood glucose response: A study of lentils. Am. J. Clin. Nutr., 36: 1093-1101.
Direct Link - Johansson, L.H. and L.A. Borg, 1988. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem., 174: 331-336.
CrossRefPubMedDirect Link - Kim, D.O., S.W. Jeong and C.Y. Lee, 2003. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem., 81: 321-326.
CrossRefDirect Link - Kim, D.K., S.C. Jeong, S. Gorinstein and S.U. Chon, 2012. Total polyphenols, antioxidant and antiproliferative activities of different extracts in mungbean seeds and sprouts. Plant Foods Hum. Nutr., 67: 71-75.
CrossRefDirect Link - Kim, J., D.H. Kim, B.H. Lee, S.H. Kang and H.J. Lee et al., 2009. Folate intake and the risk of colorectal cancer in a Korean population. Eur. J. Clin. Nutr., 63: 1057-1064.
CrossRefDirect Link - Longnecker, N., 2000. Passion for pulses: Health benefits of pulses and why Australians should eat more of them. Proc. Nutr. Soc. Aust., 24: 191-195.
Direct Link - Lopez-Amoros, M.L., T. Hernandez and I. Estrella, 2006. Effect of germination on legume phenolic compounds and their antioxidant activity. J. Food Compos. Anal., 19: 277-283.
CrossRefDirect Link - Marathe, S.A., V. Rajalakshmi, S.N. Jamdar and A. Sharma, 2011. Comparative study on antioxidant activity of different varieties of commonly consumed legumes in India. Food. Chem. Toxicol., 49: 2005-2012.
CrossRefPubMedDirect Link - McCrory, M.A., B.R. Hamaker, J.C. Lovejoy and P.E. Eichelsdoerfer, 2010. Pulse consumption, satiety and weight management. Adv. Nutr., 1: 17-30.
Direct Link - Mitchell, D.C., F.R. Lawrence, T.J. Hartman and J.M. Curran, 2009. Consumption of dry beans, peas and lentils could improve diet quality in the US population. J. Am. Dietetic Assoc., 109: 909-913.
CrossRefDirect Link - Oak, M.H., J. El Bedoui and V.B. Schini-Kerth, 2005. Antiangiogenic properties of natural polyphenols from red wine and green tea. J. Nutr. Biochem., 16: 1-8.
CrossRefDirect Link - Okada, Y. and M. Okada, 2000. Effect of a radical scavenger water soluble protein from Broad beans (Vicia faba) on antioxidative enzyme activity in cellular aging. J. Nutr. Sci. Vitaminol., 46 : 1-6.
CrossRefDirect Link - Perez-Balibrea, S., D.A. Moreno and C. Garcia-Viguera, 2011. Genotypic effects on the phytochemical quality of seeds and sprouts from commercial broccoli cultivars. Food Chem., 125: 348-354.
CrossRefDirect Link - Popescu, R.P., A. Norman, P.J. Ross, B. Parikh and D. Cunningham, 1999. Adjuvant or palliative chemotherapy for colorectal cancer in patients 70 years or older. J. Clin. Oncol., 17: 2412-2418.
PubMedDirect Link - Rao, G.N., W.W. Piegorsch and J.K. Haseman, 1987. Influence of body weight on the incidence of spontaneous tumors in rats and mice of long-term studies. Am. J. Clin. Nutr., 45: 252-260.
PubMedDirect Link - Reeves, P.G., 1997. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr., 127: 838S-841S.
PubMedDirect Link - Rice-Evans, C.A., N. Miller and G. Paganga, 1997. Antioxidant properties of phenolic compounds. Trends Plant Sci., 2: 152-159.
CrossRefDirect Link - Rodriguez-Bigas, M.A., C.R. Boland, S.R. Hamilton, D.E. Henson and S. Srivastava et al., 1997. A national cancer institute workshop on hereditary nonpolyposis colorectal cancer syndrome: Meeting highlights and Bethesda guidelines. J. Natl. Cancer Inst., 89: 1758-1762.
CrossRefDirect Link - Rustgi, A.K., 2007. The genetics of hereditary colon cancer. Genes Dev., 21: 2525-2538.
CrossRefDirect Link - Scalbert, A., C. Manach, C. Morand, C. Remesy and L. Jimenez, 2005. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr., 45: 287-306.
CrossRefDirect Link - Scheppach, W., H.P. Bartram and F. Richter, 1995. Role of short-chain fatty acids in the prevention of colorectal cancer. Eur. J. Cancer, 31: 1077-1080.
CrossRefDirect Link - Shackelford, L.A., D.R. Rao, C.B. Chawan and S.R. Pulusani, 1983. Effect of feeding fermented milk on the incidence of chemically induced colon tumors in rats. Nutr. Cancer, 5: 159-164.
CrossRefPubMedDirect Link - Singleton, V.L., R. Orthofer and R.M. Lamuela-Raventos, 1999. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In: Methods in Enzymology, Burslem, G.L. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121822002, pp: 152-178.
CrossRefDirect Link - Sud'ina, G.F., N.V. Mirzoeva, M.A. Pushkareva, G.A. Korshunova, N.V. Sumbatyan and S.D. Varfolomeev, 1993. Caffeic acid phenethyl ester as a lipoxygenase inhibitor with antioxidant properties. FEBS Lett., 329: 21-24.
CrossRefPubMedDirect Link - Surh, Y.J., 2003. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer, 3: 768-780.
CrossRefPubMedDirect Link - Talalay, P. and J.W. Fahey, 2001. Phytochemicals from cruciferous plants protect against cancer by modulating carcinogen metabolism. J. Nutr., 131: 3027S-3033S.
PubMedDirect Link - Tannenbaum, A., 1942. The genesis and growth of tumors. II. Effects of caloric restriction per se. Cancer Res., 2: 460-467.
Direct Link - Umar, A., C.R. Boland, J.P. Terdiman, S. Syngal and A. de la Chapelle et al., 2004. Revised bethesda guidelines for hereditary nonpolyposis colorectal cancer (Lynch syndrome) and microsatellite instability. J. Natl. Cancer Inst., 96: 261-268.
CrossRefDirect Link - Vidal-Valverde, C., J. Frias, I. Estrella, M.J. Gorospe, R. Ruiz and J. Bacon, 1994. Effect of processing on some antinutritional factors of lentils. J. Agric. Food Chem., 42: 2291-2295.
CrossRefDirect Link - Willett, C.C., 2001. Diet and cancer: One view at the start of the millennium. Cancer Epidemiol. Biomarkers Prev., 10: 3-8.
PubMedDirect Link - Yang, C.S., J.M. Landau, M.T. Huang and H.L. Newmark, 2001. Inhibition of carcinogenesis by dietary polyphenolic compounds. Annu. Rev. Nutr., 21: 381-406.
CrossRefPubMedDirect Link - Yang, F., T.K. Basu and B. Ooraikul, 2001. Studies on germination conditions and antioxidant contents of wheat grain. Int. J. Food Sci. Nutr., 52: 319-330.
CrossRefDirect Link - Yao, L.H., Y.M. Jiang, J. Shi, F.A.Tomás-Barberán, N. Datta, R. Singanusong and S.S. Chen, 2004. Flavonoids in food and their health benefits. Plant Foods Hum. Nutr., 59: 113-122.
CrossRefDirect Link - Yu, L., S. Haley, J. Perret, M. Harris, J. Wilson and M. Qian, 2002. Free radical scavenging properties of wheat extracts. J. Agric. Food Chem., 50: 1619-1624.
CrossRefPubMedDirect Link - Verghese, M., D.R. Rao, C.B. Chawan, L.L. Williams and L. Shackelford, 2002. Dietary inulin suppresses azoxymethane-induced aberrant crypt foci and colon tumors at the promotion stage in young fisher 344 rats. J. Nutr., 132: 2809-2813.
PubMedDirect Link - Griffith, O.W., 1980. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem., 106: 207-212.
CrossRefPubMedDirect Link