
Noura S. Abouzeinab
Department of Biological and Environmental Science, Faculty of Science, Beirut Arab University, Debbieh, Lebanon
Journal of Pharmacology and Toxicology
Year: 2015 | Volume: 10 | Issue: 1 | Page No.: 1-19







ABSTRACT
Silymarin (SIL) is a natural occurring antioxidant and has a free radical scavenging property. Cisplatin (CDDP) is a chemotherapeutic agent associated with nephrotoxicity. This study aims at investigating the antioxidant effect of SIL against CDDP-induced oxidative stress and renal toxicity in rat kidney. Thirty rats were divided into 5 groups of 6 animals each. The first group (G1) received no chemical treatment, G2 injected with 0.15 mL vehicle (propylene glycol and saline) intraperitoneally (i.p.), G3 injected with a single dose of CDDP 5 mg kg–1, i.p. only at the first day, G4 received SIL (50 mg kg–1 day–1, i.p.) 2 h after a single dose of CDDP (5 mg kg–1; i.p.) and G5 received SIL (50 mg kg–1 day–1, i.p.) 2 h before a single dose of CDDP (5 mg kg–1; i.p.). On day fifteen, animals sacrificed, serum and/or kidney tissues were used to determine renal functional markers (serum urea, creatinine), oxidative stress index (lipid peroxidation: Malondialdehyde), antioxidant enzyme activities (superoxide dismutase, SOD; glutathione peroxidase, GPx) and light microscopical analysis. The CDDP-induced oxidative stress was indicated by a marked increase in: Serum urea and creatinine concentration and tissue MDA and a significant reduction in SOD and GPx activities. The CDDP also caused severe histopahtological changes and histochemical depletion of total protein contents in kidney cells. Results suggest that pretreatment with SIL protect kidney tissues fully against CDDP toxicity, since renal markers, MDA levels; activities of antioxidant enzymes, histological and histochemical features were restored to normal levels. Hence, SIL could be a promising nephroprotective and antioxidant agent for reducing CDDP-induced renal oxidative stress and toxicity.
PDF Abstract XML References Citation
Received: January 30, 2015;
Accepted: June 20, 2015;
Published: July 07, 2015
How to cite this article
Noura S. Abouzeinab, 2015. Antioxidant Effect of Silymarin on Cisplatin-Induced Renal Oxidative Stress in Rats. Journal of Pharmacology and Toxicology, 10: 1-19.
DOI: 10.3923/jpt.2015.1.19
URL: https://scialert.net/abstract/?doi=jpt.2015.1.19
DOI: 10.3923/jpt.2015.1.19
URL: https://scialert.net/abstract/?doi=jpt.2015.1.19
INTRODUCTION
Cisplatin (CDDP) is a potent and a major chemotherapeutic drug widely used as a front line therapy for the treatment of an extensive range of solid-organ tumors (Mansour et al., 2006). The therapeutic effects of CDDP are significantly improved by dose elevation. However, the clinical use of high-dose therapy of CDDP is restricted by its cumulative nephrotoxicity (Sastry and Kellie, 2005), a side effect that was also shown in several animals’ models (Arany and Safirstein, 2003; Sastry and Kellie, 2005). The accumulation of CDDP produces clear necrotic changes within the tissues of the kidney tissue (Sastry and Kellie, 2005). Besides, it was found that about 25-35% of patients have suffered a significant decline in renal function after a single dose of CDDP treatment (Luke et al., 1992).
Kidneys are the major elimination pathway for many chemotherapeutic drugs and their metabolites (Stewart et al., 1985). Antineoplastic drugs are excreted in the urine and after 24 h, the renal toxicity concentration of platinum in the renal cortex increases to higher level than in the other organs (Wild, 2012). Although, the exact pathophysiological mechanisms concerned in CDDP induced nephrotoxicity are still proposed (Dodiya et al., 2011), several hypotheses have been suggested to explain such effect. Ajıth et al. (2007) suggested that oxidative stress, resulting from an imbalance between prooxidant and antioxidant systems in favor of the former, largely contributes to immune system deregulation and complications observed in End-Stage Renal Disease (ESRD) and patients treated with hemodialysis, thus reactive oxygen species and free radicals are involved in the nephrotoxicity induced by a synthetic anticancer drug cisplatin. Furthermore, Yao et al. (2007) proposed that unbound cisplatin is freely filtered at the glomerulus and taken up into renal tubular cells mainly by a transport-mediated process. The drug is at least partially metabolized into toxic species. Thus, cisplatin has multiple intracellular effects, including regulating genes, causing direct cytotoxicity with reactive oxygen species, activating mitogen-activated protein kinases, inducing apoptosis and stimulating inflammation and fibrogenesis. These events cause tubular damage and tubular dysfunction with sodium, potassium and magnesium wasting. Most patients have a reversible decrease in glomerular filtration but some have an irreversible decrease in glomerular filtration. Volume expansion and saline diuresis remain the most effective preventive strategies. In addition, Razo-Rodriguez et al. (2008) stated that there is growing evidence suggesting that reactive oxygen (ROS) 2-4 and nitrogen (RNS) 5,6 species are strongly related to cisplatin-induced nephrotoxicity. According to previous results, some mechanisms are involved such as: increased production of Reactive Oxygen Species (ROS) and Nitric Oxide Species (NS), reduction of sulfhydryl (SH) groups, impaired and suppressed anti-oxidant defense system by changing the mitochondrial antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), diminished glutathione (GSH) and mitochondrial dysfunction by inhibition of complexes I-IV of the respiratory chain in proximal tubules (Baek et al., 2003; An et al., 2011). In the case of the CDDP dose-limiting nephrotoxicity, oxidative stress injury, DNA damage and elevated lipid peroxidation (LPO) (Zhang et al., 1996), are the most possible mechanisms of CDDP-induced toxicity (Shimeda et al., 2005; Santos et al., 2007).
The CDDP is toxic to kidney tissues and produces renal damage manifested by tubular damage to Proximal Convoluted Tubules (PCT) (Yao et al., 2007). The CDDP gets accumulated in mitochondria of renal PCT cells with induced ROS resulting in CDDP-induced nephrotoxicity (Hanigan and Devarajan, 2003), distinguished by cellular necrosis, loss of microvillus, mitochondrial vacuolization followed by functional alterations including: inhibition of protein synthesis, reducing antioxidant enzymatic activities by GSH depletion, LPO, oxidative stress and mitochondrial dysfunction (Satoh et al., 2003). The CDDP also targets primarily at the S3 segment of the proximal tubule that reduces glomerular filtration rate, reduces the urinary concentrating ability and alters in urine volume and creatinine clearance (Kang et al., 2004; Atessahin et al., 2005), resulting in acute renal failure. In other word, a single dose of CDDP may cause acute renal dysfunction (Lau, 1999; Sultana et al., 2012). It is well known that mitochondria continuously scavenge ROS through the action of antioxidant enzymes such as SOD, CAT, GSH and glutathione peroxidase (GPx) (Richter et al., 1995; Bhat et al., 2002).
Natural herbal products verified to have potential antioxidant activities and are therefore, often administrated along with chemotherapeutic agents to provide better protection against the conventional therapy’s adverse toxic side effects (Arhoghro et al., 2012; Adejuwon et al., 2014) especially in preventing nephrotoxic disorders. Recently, a large number of studies have described the beneficial effects and promising roles of a variety of natural diet antioxidants in protecting the kidney against CDDP-induced nephrotoxicity. Therefore, to increase the clinical efficacy of CDDP, without inhibiting its anti-tumor effects, different experimental studies were carried out using a combination of CDDP with a variety of radicular scavengers, enzyme inhibitors, sulfur-containing antioxidants and natural foods with antioxidant properties to ameliorate CDDP-induced nephrotoxicity (Saad et al., 2009; Abdelmeguid et al., 2010; Ozer et al., 2011).
Silymarin (SIL) is a mixture of flavonolignans extracted from milk thistle plant (Silybum marianum) which is a strong antioxidant compound capable of scavenging both free radicals and ROS (Laekeman et al., 2003; Eminzade et al., 2008). The name "milk thistle" is derived from the milky white sap that comes from the leaves and stems when crushed or broken. The SIL has high significance in herbal medicine for its activity in the treatment and protection against variety of diseases including nephrotoxicity (Ali and Moundhri, 2006; Ali et al., 2013). The SIL deleterious effects like gastrointestinal disturbances and allergic skin rashes have been published (El-Shitany et al., 2008).
Due to multifunctional properties of SIL, the present study was designed to investigate and assess the possible potential antioxidant protective effect of SIL against CDDP-induced acute renal failure and oxidative stress in rat kidneys.
MATERIALS AND METHODS
Chemicals: Silymarin (SIL), Cisplatin [(CDDP) or cis-Dichlorodiammine Platinum (II)], SOD determination kit and GPx cellular activity assay kit were purchased from Sigma-Aldrich Chemical Company, St. Louis, MO, USA. A Commercial diagnostic kit from Bio-Diagnostic kits, Alexandia, Egypt was used for the estimation of malondialdehyde (MDA). Serum urea and serum creatinine concentration kits were obtained from Biocon® Diagnostiks, Germany.
Animals: Thirsty healthy adult male Sprague-Dawley rats, aged 10-13 weeks and weighing 140-160 g were purchased from the Faculty of Pharmacy, Beirut Arab University, Lebanon. The animals were housed conventionally in clean stainless steel wire bottom cages and fed regular rat diet and tap water ad libitum. A photoperiod of 12 h light/dark cycle at a temperature of 23±2°C and a relative humidity 60-70% was maintained. The care and treatments of the animals were approved and performed according to the guiding principles of Animal House and ethical standards of the Faculty of Pharmacy, Beirut Arab University, Lebanon.
Experimental design and treatments: The experiment was carried out in five experimental groups of 6 rats each (G1 to G5). Group G1: Control group that remained untreated. G2: Vehicle treated group i.p. injected daily with propylene glycol and saline; 75/25 v/v (0.15 mL). The G3 was CDDP nephrotoxicity rat model group, received single injection of CDDP (5 mg kg–1, i.p.) (Mansour et al., 2006) on the zero day of the experiment. The G4 was post-treatment with SIL group, received SIL (50 mg kg–1 day–1, i.p. (Karimi et al., 2005) 2 h after a single dose of CDDP (5 mg kg–1, i.p.). The G5 was pretreatment with SIL group; received single received SIL (50 mg kg–1 day–1, i.p. (Karimi et al., 2005) 2 h before a single dose of CDDP (5 mg kg–1, i.p.). Thus, the injection of SIL as supportive treatments in G4 and G5 continued daily till the end of the experimental duration (2 weeks). The animals’ clinical behavior, food intake, body weight gain and wet kidney weight among all rats were recorded four times weekly till the end of the experiment (Abdelmeguid et al., 2010).
Sample preparation: After the last dose of the treated groups, rats were fasted for 12 h. On the fifteenth day, animals were anesthetized and sacrificed by cervical decapitation. Blood samples were taken out by a cardiac, collected in heparinized tubes and centrifuged at 3000× g for 10 min to separate serum in order to measure the level of urea and creatinine. Kidneys were excised, trimmed of adhering connective tissues, rinsed with saline to eliminate blood contamination and dried by blotting with filter paper. The first right side kidney was used to prepare kidney homogenates that were obtained using a tissue homogenizer WiseTis digital homogenizer, HG -15D (DIHAN Scientific CO., Ltd., Korea).
The homogenates were prepared by using a 100 mM KCl buffer (7:00 pH) containing EDTA 0.3 mM; all homogenates were centrifuged at 4000× g for 10 min at 4°C and supernatants were stored at-70°C for MDA, SOD and GPx biochemical measurements. The second left side kidney was cut into small pieces, immediately dropped and fixed in 10% neutral formalin (in 0.1 M phosphate buffer, pH 7.4, for 24 h), processed routinely, embedded in Paraffin wax and sectioned at 6 μm for histopathological and histochemical studies.
Biochemical analysis: In the present study, biochemical analysis were conducted to determine: renal functional markers (levels serum urea and serum creatinine concentrations), oxidative stress index (LPO: MDA) and antioxidant enzyme activities (SOD and GPx). The assessments of the biochemical assays were carried out according to the standard procedures using their related commercially diagnostic kits. Total protein concentration in kidney homogenate was obtained by the method of Bradford (1976) using Bovine Serum Albumin (BSA) as the standard at 595 nm.
Renal functional markers: Serum samples were assayed for urea and creatinine concentrations. The concentrations of serum urea [Urease-Berthelot Reaction (Fluitest® UREA)] and serum creatinine [Creatinine Kinetic-JAFFE Method (Fluitest® CREA, KINETIC)] were estimated colorimetrically and spectrophotometrically based on Jaffe’s method according to Masson et al. (1981), respectively.
Oxidative stress index: The kidney LPO end product MDA was determined spectrophotometrically by measuring Thiobarbituric Acid Reactive Substances (TBARS), using commercially existing diagnostic kits. The absorbance of the pink product obtained was measured at 534 nm (Ohkawa et al., 1979). The activities of SOD (SOD determination kit, EC 1.15.1.1) and GPx (GPx, cellular activity assay kit, EC 1.11.1.9) were determined in filtrates of kidney homogenates kinetically and spectrophotometrically, respectively.
Light microscopical studies: Kidney sections underwent Hematoxylin-Eosin (H and E) staining for structural and histopathological analysis. Histopathological changes were evaluated semiquantitatively where a minimum of 10 fields for each kidney slide was examined and assigned for severity of changes using the following scale: -, none; +, mild damage; ++, moderate damage and +++, severe damage. On the other hand, some other kidney sections were analyzed by histochemical light microscopy and were stained with mercury bromophenol blue (Hg-BPB) in order to detect total proteins (Mazia et al., 1953).
Statistical analysis: SPSS Version 16 statistical software package (SPSS, Cary, NC, USA) was used for statistical analysis. Values expressed as Means±SEM. Differences among the experimental groups were estimated using one-way analysis of variance (ANOVA) and the group means were compared by LSD-post hoc test for multiple comparisons. Values were considered statistically significant when p<0.05.
RESULTS
The effect of CDDP and SIL treatments clinical behavior, food intake, body weight gain and wet kidney weights were depicted in our previous work (Abdelmeguid et al., 2010). No mortality or obvious signs of toxicity occurred in any of the treatment groups. Animals treated with CDDP alone resulted in significant decreases in feed intake and body weight gain accompanied with a significant increase of kidney wet weight. Besides, animals became slightly nervous, less active, with minimal loss of furring. Whereas, animals receiving both types of SIL treatments were comparable to control groups and exhibited no behavioral changes or clinical symptoms, i.e., body weight gain, food intake and wet kidney weight returned to normal values similar to those recorded in control groups (Abdelmeguid et al., 2010).
Biochemical investigations
Renal functional markers: According to Fig. 1 and 2, serum urea and creatinine concentrations were significantly h igher in CDDP-treated group than the controls. However, post-administration of SIL significantly reversed these changes but the values were still significantly higher when compared to control groups (Fig. 1). Yet, the effect of pretreatment with SIL was more efficient when compared to those in post-treated group, as values returned significantly to those recorded in control group (Fig. 1 and 2).
Oxidative stress index: Table 1 shows oxidative status in rat kidney tissues after CDDP, CDDP +SIL treatments. Tissue MDA levels were significantly increased in CDDP-treated group when compared to control rats.
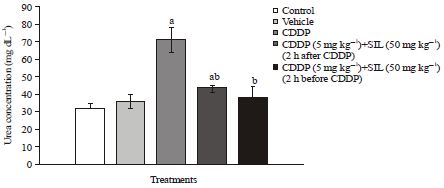 | |
| Fig. 1: | Effect of treatment on serum urea concentration. Bars are Mean±SEM for 6 animals per group. Statistically significant at p<0.05, aComparison of control and other groups, bComparison of CDDP-treated group with other treatment groups |
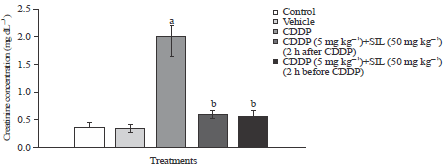 | |
| Fig. 2: | Effect of treatment on serum creatinine concentration. Bars are Mean±SEM for 6 animals per group. Statistical significance at p<0.05, aComparison of control and other groups, bComparison of CDDP-treated group with other treatment groups |
| Table 1: | Effect of silymarin and cisplatin on the oxidative stress index; and antioxidant enzymes in kidney tissues of control and experimental rats |
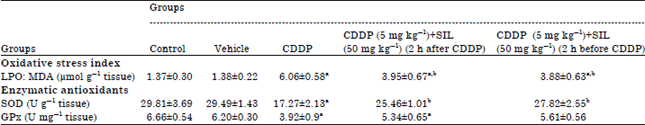 | |
Data is the Means±SEM for six rats in each group. Statistical significance between means was performed using one-way analysis of variance (ANOVA) followed by LSD as a post-hoc ANOVA test for multiple comparisons. Statistical significance at p<0.05, a Comparison of control and other groups. bComparison of CDDP-treated group with other treatment groups, CDDP: Cisplatin, SIL: Silymarin, LPO: Lipid peroxidation, SOD: Superoxide dismutase and Gpx: Glutathione peroxidase | |
However, both treatments of SIL significantly decreased CDDP induced LPO (MDA) levels; yet, values were close to those in control groups but remained significantly elevated over basal values.
Antioxidant enzymes: The activities of selected antioxidant enzymes (SOD and GPx) in the kidney tissues of the experimental groups are presented in Table 1. In CDDP-treated group, the activities of SOD and GPx were significantly decreased compared to control rats. Both treatments of SIL showed full protection manifested by a significant increase in the depleted activities of SOD induced by CDDP, where values return to those normal baselines recorded in control groups (Table 1). On the other hand, SIL post treatment showed a slight elevation in GPx activities compared to CDDP-treated group but the results were not significant pretreatment (Table 1). However, pretreatment with SIL fully prevented the decreased activities of GPx induced in CDDP-treated group. Thus, GPx activities were significantly higher, when compared to CDDP alone, where values approached those in control groups and were not statistically significant over basal values (Table 1).
Light microscopical observations: The histopathological and histochemical changes demonstrated the protective effects of co-administration of the supportive treatment of SIL in different experimental groups in comparison to CDDP rat model. Thus, prominent alterations and high cellular damage were observed in the kidney sections of rats treated with CDDP after both H and E and Hg-BPB staining. However, light microscopical analysis of the rat’s kidney of control and vehicle groups, revealed no abnormal changes (Fig. 3a-b and 4a-b).
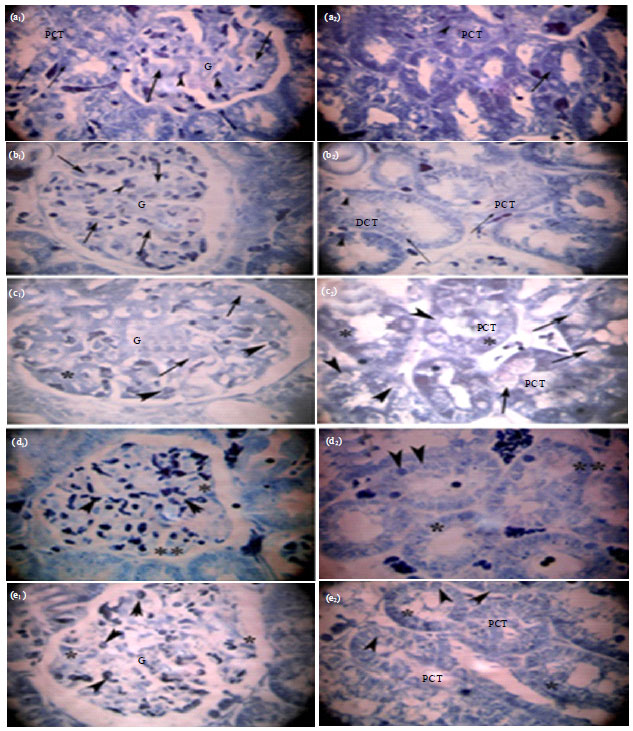
Histopathological examinations: After two weeks, the kidney structures (renal cortex and medulla) of H and E stained sections revealed that CDDP treatment caused necrosis and interstitial cells infiltration among renal tubules. Numerous glomeruli showed glomerular atrophy and congestion with dilated urinary space and disrupted bowman’s capsule at certain foci (Fig. 3c). Besides, CDDP caused a marked tubular degeneration (Fig. 3c). In particular, numerous Proximal Convoluted Tubules (PCT) displayed disruption of the cellular architecture with loss and shedding of cells, pale cytoplasm due to vacuolization and disappearance of the brush borders. A number of PCT cells appeared hypertrophied; others are small with pyknotic nuclei (Fig. 3c). The administration of SIL post-treatment decreased CDDP-induced renal cellular to a minimal (Fig. 3d). However, pretreatment with SIL markedly prevented congestion of glomeruli and other tubular alterations (Fig. 3e).
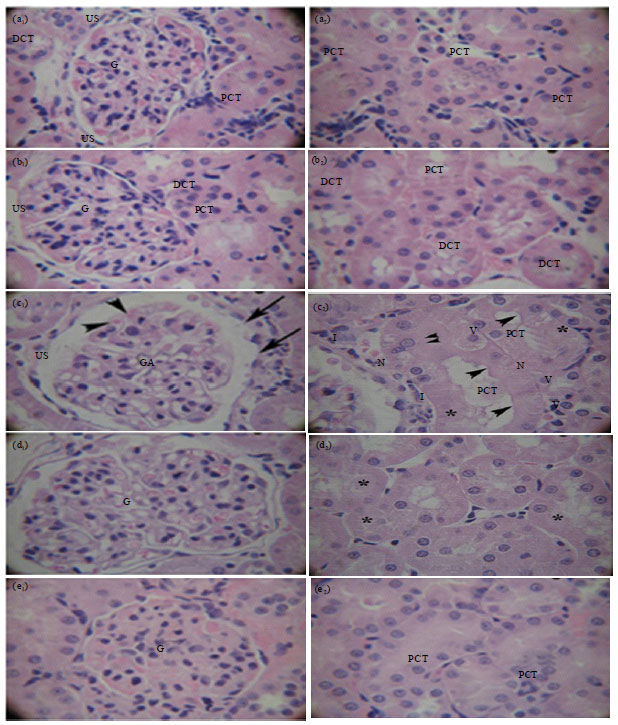 | |
| Fig. 3(a-e): | Light micrographs of sections of rats’s renal cortex renal cortex (a1, b1, c1, d1, e1) and renal medulla (a2, b2, c2, d2, e2) in control and treated groups (a) Control group with normal kidney histology, (b) Vehicle group appear normal with no apparent pathology, (c) CDDP treatment; a1: Glomerular Atrophy (GA) with congestion (arrow head), dilated urinary space (US), disrupted Bowman’s capsule (arrow), a2: Degenerated and necrotic, (*) proximal convoluted tubule (PCT): loss of brush borders (arrow head), shedding of cells (double arrow), hypertrophied cells with cytoplasmic vacuolization (V), pyknotic nucleus (N), peritubular cellular infiltration (I), (d) SIL post-treatment, b1: Mild dilatation and congestion of glomerulus (G), b2: Minimal tubular damage (*) and (e) SIL pretreatment, c1 and c2: No histological abnormalities and their prevention by SIL, normal glomerulus (G) and PCT, (H and E stained sections, Magnification: x400) |
 | |
| Fig. 4(a-e): | Light micrographs of sections of rats’s renal cortex (a1, b1, c1, d1, e1) and renal medulla (a2, b2, c2, d2, e2) in control and treated groups (a) Normal and (b) Vehicle groups, with dark blue coarse protein granules giving intense reactivity within the nuclei (arrow head) and cytoplasm (arrow) of glomerular (G) and PCT cells, (c) CDDP treatment, reduced weakly stained (*) and negatively reactive (arrow) total protein within cytoplasm of glomerular (G) and PCT cells, nuclei (arrowhead) giving weak reaction, (d) SIL post-treatment, moderately (*) and densely (**) stained protein particles within cytoplasm, nuclei giving moderate reaction (arrowhead), (e) SIL pretreatment; intensely reactive protein granules within cytoplasm (asterisk), nuclei (arrowhead) of glomerular (G) and PCT cells (Hg-BPB reaction; Magnification: x400) |
| Table 2: | Semiquantitative grading of the histopathological changes in kidney sections of rats of control and treated groups |
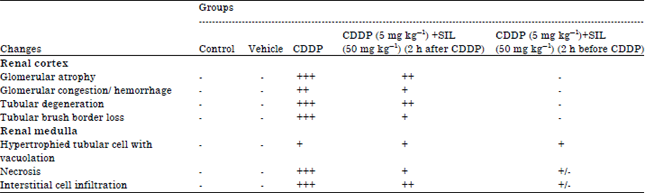 | |
-: None, +: Mild damage, ++: Moderate damage, +++: Severe damage, are semiquantitative scores assessed by histological analysis of kidneys in rats (n = 10 fields), CDDP: Cisplatin and SIL: Silymarin | |
The intensity of histopathological lesions demonstrated the protective effect of co-administration of supportive treatments in different groups was scored as shown in Table 2.
Histochemical examinations: Histochemical examinations of Hg-BPB stained kidney sections showed that CDDP treatment exhibited a severe reduction in total protein content after two weeks. Degeneration of total protein content was detected as minimal and weakly stained protein particles within the nuclei and cytoplasm of glomerular and PCT cells. The cytoplasm in some PCT cells appeared negatively stained (Fig. 4c). The supportive administration of SIL after CDDP treatment improved the reduction of total protein content among renal cells induced by CDDP, i.e. both types of renal cells displayed the aggregation of medium-sized protein granules with moderate reaction to Hg-BPB (Fig. 4d). In contrast, pretreatment with SIL regained total protein content similar to those recorded in control groups; where protein granules were large in size, intensely stained, associated with nucleoli and concentrated adjacent to the nuclear envelope in most nuclei and scattered within the cytoplasm of both the glomerular and PCT cells. However, few cells showed nuclei with moderate reaction and intense reactivity within cytoplasm (Fig. 4e).
DISCUSSION
Nephrotoxins provoke renal injury to one or both of the kidneys since, they have direct effect on the glomerulus or the renal tubules and leads to damage of cells. Most drugs are found to cause nephrotoxicity or toxic effects that include altered inflammation, tubular cell toxicity, intraglomerular hemodynamics (Zager, 1997; Schetz et al., 2005). However, most cancer patients suffer from chemotherapeutic drugs-induced nephrotoxicity along with renal failure and electrolyte disturbances (Kufe et al., 2003).
The CDDP is one of the common and effective antineoplastic drugs often used to treat wide variety of solid tumors. Unfortunately, its uses in the clinical and therapeutic fields were markedly limited due to the incidence of many side effects mainly the nephrotoxicity (Saad et al., 2009; Benedetti et al., 2013), the most important dose-limiting adverse effect (Muggia et al., 2000; Dunton, 2002). In other words, the unbalanced accumulation of CDDP in kidney tissue may result in nephrotoxicity (Arany and Safirstein, 2003) and may cause acute renal failure even after a single dose (Lau, 1999; Sultana et al., 2012). Several researches have been conducted to explain how CDDP could stimulate toxicity (Saad et al., 2009; Ali et al., 2013; Benedetti et al., 2013). However, one of the main mechanisms of CDDP nephrotoxicity is the oxidative stress (Mansour et al., 2002; Sueishi et al., 2002; Uehara et al., 2011). The CDDP initially activates oxidative stress in the mitochondria of kidney PCT and endothelial cells, followed by a secondary wave of ROS/RNS (reactive nitrogen species) production, deterioration of mitochondrial structure and function, thus suppressing mitochondrial antioxidants and cellular injury with consequent renal damage (Yao et al., 2007; Gautier et al., 2010).
Antioxidants were proposed to have a protective role against CDDP-induced nephrotoxicity mediated by oxidative stress (Mansour et al., 2002; Sueishi et al., 2002; Uehara et al., 2011). Therefore, to improve the toxic effects of CDDP, many approaches have been proposed using diverse natural and synthetic free radical scavengers and antioxidants (Mansour et al., 2006; Ajıth et al., 2007; Yao et al., 2007; Saad et al., 2009; Abdelmeguid et al., 2010; Ali et al., 2013; Nasr, 2013) that might provide nephroprotection against CDDP toxicity (Ali et al., 2013; Ali and Moundhri, 2006). The SIL protective and ameliorative effects appear to be due to its power in scavenging free radicals and chelating metal ions (Borsari et al., 2001). Several studies have proposed that silymarin is able of protecting cells directly by its ability to stabilize the membrane permeability through inhibiting LPO (Mira et al., 1994) and preventing glutathione depletion (Valenzuela et al., 1989). Moreover, SIL has been shown to be harmless in animal models and no significant undesirable reactions were stated in human studies (Hogan et al., 2007).
Serum creatinine and urea levels were considered the major factors that determine the glomerular filtration rate (Azu et al., 2010). Serum creatinine is an important indicator of renal health and is most likely the most broadly used indirect measure of glomerular filtration rate and renal failure (Jarup, 2002), where creatinine is primarily filtered out of the blood by the kidneys through glomerular filtration and proximal tubular secretion. If the filtering of the kidney is damaged, serum creatinine level increases. The CDDP toxicity induced severe renal dysfunction that allows the elevated secretion of creatinine from the proximal tubules, a marker of irreversible renal tubular injury (Dickey et al., 2008). Renal function can also be determined by analyzing the serum urea level. During the metabolism of protein in the body, the liver produces ammonia which is transformed into a byproduct called urea. However, due to renal dysfunction, urea is released into the bloodstream as serum urea. Therefore, higher serum urea level is directly proportional to harshness of renal damage (Dickey et al., 2008).
In the present study, nephrotoxicity of CDDP was manifested from the marked elevated levels of serum creatinine and urea levels compared to control groups. The escalation in the serum levels of these renal functional markers might be caused by the direct toxic effect of CDDP that resulted in: impaired renal functions (Anusuya et al., 2013), glomerular injury, tubular obstruction and/or the back-leakage of the renal tubules (Azu et al., 2010). The alterations in glomerular function in CDDP-treated rats may also be secondary to ROS (Somani et al., 2000) which provoke mesangial cells contraction, modifying the filtration surface area and altering the ultrafiltration coefficient factors that decrease the glomerular filtration rate (Aydogan et al., 2008). Such ROS might play a role in the pathogenesis of tubular cell apoptosis but probably had no effect in the tubular necrosis. Such functional disorders in CDDP exposed rats could indicate the ability of CDDP to inhibit protein synthesis in the tubular cells (Adejuwon et al., 2014) or to initiate LPO and generate free radicals in renal tubules (Mansour et al., 2006; Ajıth et al., 2007; Yao et al., 2007; Saad et al., 2009) inducing tubular necrosis in a concentration-dependent manner (Uehara et al., 2011). Similar results were formerly reported in various studies (Mansour et al., 2006; Razo-Rodriguez et al., 2008; Azu et al., 2010; Arhoghro et al., 2012). In agreement with our results, Mansour et al. (2002) and Sueishi et al. (2002) stated that the nephrotoxic effect of CDDP was indicated by a significant increase of creatinine and urea levels.
In the present investigation, the histopathological changes validated the biochemical findings, where the cortico-medullary zones of CDDP-treated kidney sections revealed extensive glomerular damage as well as tubular disturbances including: reduction in the size of glomeruli, dilatation of bowman’s capsules, degeneration, necrosis, detachment of the epithelial cell lining, shedding of cell and loss of brush borders of the proximal convoluted tubules. These histological findings could explain the disorders of both glomerular and tubular functions, where these disturbances might be the main reason of reduced glomerular filtration rate which is greatly associated with the biochemical elevation of serum creatinine and urea levels. Similar to the findings of the present study, the nephrotoxic effect of CDDP was accumulated mainly on the PCT at the cortico-medullary zone (Townsend and Hanigan, 2002), where the necrotic and apoptotic changes of the renal tubular cells are possibly due to the direct effect of the intracellular CDDP (Mansour et al., 2002; Sueishi et al., 2002). In disagreement with this, enormous necrosis and subsequent regeneration of renal PCT cells with minor papillary injury and no glomerular morphological changes were observed in CDDP-treated rats (Nagothu et al., 2005).
However, in the present study, pretreatment with SIL revealed marked improvement in the functional indices in the kidney as it completely prevented the increases in serum urea and creatinine concentration and returned them to control levels and it succeeded in the full protection against CDDP-induced renal damage. These results confirmed the ability of SIL to normalize the renal function via its scavenging power for free radicals (Borsari et al., 2001). In accordance to our results, Karimi et al. (2005) suggested that rats receiving SIL or its extract (50 mg kg–1) for 5 days, 2 h before CDDP prevented the increases in serum urea and creatinine concentrations, as well as tubular damage. Our results are in agreement with a previous study that showed i.v silibinin (200 mg kg–1), 1 h before the CDDP (5 mg kg–1, i.p.) prevented the effects of CDDP on creatinine clearance and proteinuria and reduced morphological modifications in PCT as showed by Gaedeke et al. (1996). Similar results were reported by Shimeda et al. (2005), Behling et al. (2006) and Lee et al. (2007), who demonstrated the complete protective effects of the pretreatments of flavonoids and antioxidants against CDDP-induced increase in renal functional markers and rendered the rats less susceptible to kidney damage induced by treatment with CDDP. The present work focused on the total preventive effect of SIL pretreatment, since glomeruli as well as renal tubules restored their normal architecture. These results concur with those previously reported by Gaedeke et al. (1996), who revealed that female Wistar rat’s receiving i.v. silibinin (200 mg kg–1) 1 h prior to CDDP administration for 11 days, significantly ameliorated both proximal tubular and glomerular damage induced by CDDP.
Rats treated with SIL 2 h after CDDP, however, had serum urea and creatinine levels significantly lower than those receiving CDDP alone. Besides, mild to moderate cell injury was observed in our microscopical preparations that showed attenuation of glomeruli and tubular atrophy. In accordance to our description, El-Abd and Okda (2007) suggested that S.D. rats receiving ip injection of SIL (250 mg kg–1) for 1-3 weeks, decreased and recovered histopathological abrasions induced by ribavirin (antiviral drug) specifically renal glomeruli.
Several investigators showed that CDDP-induced acute nephrotoxicity is correlated with increased LPO and antioxidants’ activities impairment (SOD and GPx) in renal tissue (Somani et al., 2000; Saad and Al-Rikabi, 2002; Borrego et al., 2004; Gonzalez et al., 2004). The LPO is attributed to a free radical-mediated chain reaction that damages cell membranes (Velmurugan et al., 2004). Thus, the alteration in kidney function was escorted by an increase in LPO end product, MDA contents, in kidney tissue (Somani et al., 2000; Santos et al., 2008). Oxidative stress can damage proteins and DNA that are more significant targets of injury than lipids and LPO which often occur in the late injury process of nephrotoxicity (Halliwell and Chirico, 1993), leading to the development of chronic renal failure (Vaziri et al., 2003). Natural antioxidants protect the functional and structural molecules against ROS-mediated modifications, thereby preventing cytotoxicity. The natural antioxidant system contains a series of antioxidant enzymes (SOD, GPx) as well as several endogenous and dietary antioxidant compounds which are capable of reacting with and inactivating ROS (Vaziri et al., 2003). The SOD represents a front line defense against ROS-mediated injury (Vaziri et al., 2003). Besides, GPx is considered the most essential antioxidant enzyme in humans which is highly expressed in the kidney, involved in scavenging and inactivating hydrogen and lipid peroxides, providing protection to the body against oxidative stress and also removes peroxides and peroxynitrite that can lead to renal damage (De Haan et al., 2005).
Results obtained from the present study demonstrated that a significant elevation in LPO end product, manifested by MDA concentration was in parallel to a significant decrease in SOD and GPx activities in the renal tissue of CDDP-treated rat, compared to control groups. It could be suggested that CDDP is entailed in changing the thiol status of the tissue with instantaneous modifications in the enzymatic antioxidants. This reduction in renal enzymatic activities involved an oxidative type of injury with CDDP-induced nephrotoxicity or may be a resulting event following the CDDP-induced elevation in free radical production and/or decrease in LPO protecting enzymes (Behling et al., 2006). Our assumption agreed with previous studies reported by Mora et al. (2003), Shimeda et al. (2005) and Lee et al. (2007), who displayed that the activities of GPx and SOD, are potentially referred to inactivation by the increase in Reactive Oxygen Species (ROS) or lipid peroxides when oxidative damage is severe. In addition, Baek et al. (2003) and Yilmaz et al. (2004) stated the contribution of CDDP-induced mitochondrial dysfunctions in rat renal cells, particularly the inhibition of the electron transfer system. This may result in return in the increased generation of free radicals such as the production of superoxide anions, hydrogen peroxide and hydroxyl radicals. The hydroxyl radical is able to initiate LPO after abstracting a hydrogen atom from polyunsaturated fatty acids in membrane lipids. These radicals can provoke severe cellular injury and adverse tissue damage (Antunes et al., 2000; Emerit et al., 2001). Besides, hydroxyl radicals can react with macromolecules, such as membrane lipids, proteins and nucleic acids; thus varying the biological membrane integrities (Antunes et al., 2000; Emerit et al., 2001) and in return increasing the outflow of renal functional markers (urea and creatinine). These observations also support the hypothesis that part of the mechanism of toxicity in the CDDP-treated animals is correlated to exhaustion of antioxidants.
On the other hand, the present investigation showed that SIL pretreatment resulted in a full recovery and protection against CDDP-induced nephrotoxicity and oxidative stress, than that of SIL post-treatment, where kidney MDA levels and both SOD and GPx activities returned to normal values. Confirming our data, El-Adawi et al. (2011) reported that the scavenging properties of SIL against nephrotoxicity was evident in the inhibition of LPO by reducing the high levels of MDA and in the same time increasing the activity levels of antioxidant enzymes especially the GPx activity. Previous studies showed that the protective action of SIL is linked with its antioxidant properties (Aruoma, 1994). It is proposed that SIL has a hydroxyl group at C5 in addition to the carbonyl group at C4 that can chelate ferrous iron (Aruoma, 1994). This chelation can play a role in increasing the activity possibly by site specific scavenging (Abu Ghadeer et al., 2001). Besides, the free hydroxyl groups at C5 and C7 on the SIL structure can also react with peroxy radicals, thus supporting the inhibition of LPO. In addition, SIL can act as a plasma membrane stabilizer and enzyme inactivation, thus leading to a significant elevation in the cellular antioxidant protective machinery (Abu Ghadeer et al., 2001). This may also direct toward the possible de novo synthesis of these enzymes induced by the components of SIL; in addition to the increased activity of glutathione system that represents the more important defense mechanisms against damage by free radicals (Soto et al., 2003). Our studies displayed that SIL is effective in protecting against CDDP-induced oxidative damage due to its radical scavenging power. Our results are in agreement with previous study that showed, protective effect of SIL against damage to kidney from acetaminophen, CDDP (platinol) and vincristine (oncovin) (Sonnenbichler et al., 1999) and the protective effect of SIL against CDDP nephrotoxicity (Karimi et al., 2005). Recent evidence (Kaur et al., 2010) suggests that SIL may be just as essential for kidney health. The SIL concentrates in kidney cells, where it assists in repair and regeneration by increasing protein and nucleic acid synthesis and free radical scavenger and antioxidant properties against several nephrotoxins including cisplatin (Shahbazi et al., 2012).
Biochemical parameters can be considered as sensitive indicators to alterations due to xenobiotics and essential diagnostic parameters in toxicological studies (Radwan et al., 2008). In the present study, biochemical parameters were greatly associated with the renal histochemical results. Proteins are chiefly involved in the structural design of the cell (Radwan et al., 2008). In the present investigation, Hg-BPB staining demonstrated a sharp reduction in the total protein content in both renal glomerular and PCT cells in rats exposed to CDDP. Hence, our histochemical examination confirmed our biochemical results regarding CDDP undesirable effects on protein contents. As a result, total protein content revealed a great depression manifested by a decrease in the enzymatic activities of both GPx and SOD. Our results were correlated with those reported by El-Abd and Okda (2007), who reported a significant decrease in total protein content in the renal tissue cells of male albino rats. Ebaid et al. (2007) confirmed that the decrease in protein could be related to the disruption of lysosomal membranes under the effects of different toxins resulting in the liberation of their hydrolytic enzymes in the cytoplasm and thus leading in the lysis and dissolution of the target material. It could be proposed that additional energy was required to detoxify and overcome CDDP-induced stress. As a result, the next alternative source of energy needed to meet the increased energy requirement is proteins (Lehninger et al., 2008). In the present study, kidney sections stained with Hg-BPB revealed that SIL pretreatment displayed more protective action than SIL post-treatment, where SIL completely restored total protein content within the nuclei and the cytoplasm of most renal glomerular and tubular cells that exhibited intense reaction; thus returning them to normal appearance. It could be suggested that the present histochemical results confirmed our biochemical investigation, as the intense increase of total protein can be explained by the significant increase in the enzymatic activities of both GPx and SOD in renal tissues. In accordance with our findings, El-Abd and Okda (2007) stated that rats treated with SIL restored remarkably the total protein contents in renal cells.
CONCLUSION
In conclusion, the present findings imply that CDDP induces nephrotoxicity by changing kidney histological architecture, renal functional biomarkers’ activities, provoking LPO, reducing antioxidant system and changing protein metabolism at the histochemical levels. Yet, our present results reveal that SIL pretreatment significantly attenuated CDDP-induced toxicity and oxidative stress in renal tissues in rats. The histopathological, histochemical and biochemical studies validate that SIL has ameliorative and nephroprotective activities. The SIL pretreatment reduced serum urea and creatinine concentration and MDA levels, whereas antioxidant enzymes activities as well as protein contents in kidney tissues were elevated and returned to control baseline values. Thus, these data may have research hints at therapeutic and protective uses of SIL. The SIL may be considered as a potentially valuable agent and nephroprotectant in the combination with chemotherapy with CDDP. Thus, SIL may be beneficial in the prevention of CDDP-induced nephrotoxicity. In addition, its nephroprotective effect may encourage physicians to prescribe SIL before chemotherapy with CDDP. However, additional clinical studies are needed to evaluate the protective role of SIL treatment in combination with CDDP in humans. Besides, the exact mechanism of SIL’s effect in term of molecular level (including some regulatory and transcription factors) could be further investigated.
ACKNOWLEDGMENTS
The author thankfully acknowledges serious reading of the manuscript by Nabila E. Abdelmeguid, Professor of Cell Biology and Histology, Department of Zoology, Faculty of Science, Alexandria University, Egypt.
REFERENCES
- Abdelmeguid, N.E., H.N. Chmaisse and N.S. Abuo Zeinab, 2010. Protective effect of silymarin on cisplatin-induced nephrotoxicity in rats. Pak. J. Nutr., 9: 624-636.
CrossRefDirect Link - Abu Ghadeer, A.R.M., S.E. Ali and S.A.A. Osman, 2001. Antagonistic role of silymarin against cardiotoxicity and impaired antioxidation induced by adriamycin and/or radiation exposure in albino rats. Pak. J. Biol. Sci., 4: 604-607.
CrossRefDirect Link - Adejuwon, A.S., O. Femi-Akinlosotu, J.O. Omirinde, O.R. Owolabi and A.M. Afodun, 2014. Launaea taraxacifolia ameliorates cisplatin-induced hepato-renal injury. Eur. J. Med. Plants, 4: 528-541.
Direct Link - Ajith, T.A., S. Usha and V. Nivitha, 2007. Ascorbic acid and alpha-tocopherol protect anticancer drug cisplatin induced nephrotoxicity in mice: a comparative study. Clin. Chim. Acta, 375: 82-86.
CrossRefPubMedDirect Link - Ali, B.H. and M.S. Al Moundhri, 2006. Agents ameliorating or augmenting the nephrotoxicity of cisplatin and other platinum compounds: A review of some recent research. Food Chem. Toxicol., 44: 1173-1183.
CrossRefDirect Link - Ali, D.A., A.M. Abdeen, M.F. Ismail and M.A. Mostafa, 2013. Histological, ultrastructural and immunohistochemical studies on the protective effect of ginger extract against cisplatin-induced nephrotoxicity in male rats. Toxicol. Ind. Health.
Direct Link - An, Y., H. Xin, W. Yan and X. Zhou, 2011. Amelioration of cisplatin-induced nephrotoxicity by pravastatin in mice. Exp. Toxicol. Pathol., 63: 215-219.
CrossRefDirect Link - Antunes, L.M.G., J.D.C. Darin and M.L.P. Bianchi, 2000. Protective effects of vitamin C against cisplatin-induced nephrotoxicity and lipid peroxidation in adult rats: A dose-dependent study. Pharmacol. Res., 41: 405-411.
CrossRefDirect Link - Anusuya, N., P. Durgadevi, A. Dhinek and S. Mythily, 2013. Nephroprotective effect of ethanolic extract of garlic (Allium sativum L.) on cisplatin induced nephrotoxicity in male wistar rats. Asian J. Pharm. Clin. Res., 6: 97-100.
Direct Link - Arany, I. and R.L. Safirstein, 2003. Cisplatin nephrotoxicity. Semin. Nephrol., 23: 460-464.
CrossRefPubMedDirect Link - Arhoghro, E.M., E.O. Anosike and A.A. Uwakwe, 2012. Ocimum gratissimum aqueous extract enhances recovery in cisplatin-induced nephrotoxicity in albino wistar rats. Indian J. Drugs Dis., 1: 129-142.
Direct Link - Aruoma, O.I., 1994. Nutrition and health aspects of free radicals and antioxidants. Food Chem. Toxicol., 32: 671-683.
CrossRefPubMedDirect Link - Atessahin, A., S. Yilmaz, I. Karahan, A.O. Ceribasi and A. Karaoglu, 2005. Effects of lycopene against cisplatin-induced nephrotoxicity and oxidative stress in rats. Toxicology, 212: 116-123.
CrossRefPubMedDirect Link - Aydogan, S., H. Yapislar, S. Artis and B. Aydogan, 2008. Impaired erythrocytes deformability in H2O2-induced oxidative stress: Protective effect of L-carnosine. Clin. Hemorheol. Microcirc., 39: 93-98.
CrossRefDirect Link - Azu, O.O., I.O.D. Francis, A.O. Abraham, C.N. Crescie, O.E. Stephen and O.O. Abayomi, 2010. Protective agent, Kigelia Africana fruit extract, against cisplatin-induced kidney oxidant injury in Sprague-Dawley rats. Asian J. Pharm. Clin. Res., 3: 84-88.
Direct Link - Behling, E.B., M.C. Sendao, H.D. Francescato, L.M. Antunes, R.S. Costa and L. Bianchi-Mde, 2006. Comparative study of multiple dosage of quercetin against cisplatin-induced nephrotoxicity and oxidative stress in rat kidneys. Pharmacol. Rep., 58: 526-532.
PubMedDirect Link - Benedetti, G., L. Fredriksson, B. Herpers, J. Meerman, B. van de Water and M. de Graauw, 2013. TNF-α-mediated NF-κB survival signaling impairment by cisplatin enhances JNK activation allowing synergistic apoptosis of renal proximal tubular cells. Biochem. Pharmacol., 85: 274-286.
CrossRef - Bhat, S.G., S. Mishra, Y. Mei, Z. Nie, C.A. Whitworth, L.P. Rybak and V. Ramkumar, 2002. Cisplatin up-regulates the adenosine A 1 receptor in the rat kidney. Eur. J. Pharmacol., 442: 251-264.
CrossRef - Borrego, A., Z.B. Zamora, R. Gonzalez, C. Romay and S. Menendez et al., 2004. Protection by ozone preconditioning is mediated by the antioxidant system in cisplatin-induced nephrotoxicity in rats. Mediators Inflammat., 13: 13-19.
CrossRef - Borsari, M., C. Gabbi, F. Ghelfi, R. Grandi, M. Saladini, S. Severi and F. Borella, 2001. Silybin, a new iron-chelating agent. J. Inorganic Biochem., 85: 123-129.
CrossRefPubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - De Haan, J.B., N. Stefanovic, D. Nikolic-Paterson, L.L. Scurr and K.D. Croft et al., 2005. Kidney expression of glutathione peroxidase-1 is not protective against streptozotocin-induced diabetic nephropathy. Am. J. Physiol. Renal Physiol., 289: F544-F551.
CrossRef - Dickey, D.T., L.L. Muldoon, N.D. Doolittle, D.R. Peterson, D.F. Kraemer and E.A. Neuwelt, 2008. Effect of N-acetylcysteine route of administration on chemoprotection against cisplatin-induced toxicity in rat models. Cancer Chemotherapy Pharmacol., 62: 235-241.
CrossRef - Dodiya, H., M. Jain and S. Goswami, 2011. Study of urinary biomarkers for nephrotoxicity in wistar rats. J. Pharmacol. Toxicol., 6: 571-579.
CrossRefDirect Link - Dunton, C.J., 2002. Management of treatment-related toxicity in advanced ovarian cancer. Oncologist, 7: 11-19.
CrossRefPubMedDirect Link - Ebaid, H., M.A. Dkhil, M.A. Danfour, A. Tohamy and M.S. Gabry, 2007. Piroxicam-induced hepatic and renal histopathological changes in mice. Libyan J. Med., 2: 82-89.
CrossRef - El-Abd, S. and Y. Okda, 2007. Ameliorative role of silymarin against ribavarin induced toxicity in the kidney of albino rat. J. Exp. Biol. Zool., 3: 127-133.
Direct Link - El-Adawi, H., D. El-Azhary, A. Abd El-Wahab, M. El-Shafeey and M. Abdel-Mohsen, 2011. Protective effect of milk thistle and grape seed extracts on fumonisin B1 induced hepato-and nephro-toxicity in rats. J. Med. Plants Res., 5: 6316-6327.
Direct Link - El-Shitany, N.A., S. El-Haggar and K. El-Desoky, 2008. Silymarin prevents adriamycin-induced cardiotoxicity and nephrotoxicity in rats. Food Chem. Toxicol., 46: 2422-2428.
CrossRefDirect Link - Emerit, J., C. Beaumont and F. Trivin, 2001. Iron metabolism, free radicals and oxidative injury. Biomed. Pharmacother., 55: 333-339.
Direct Link - Eminzade, S., F. Uras and F.V. Izzettin, 2008. Silymarin protects liver against toxic effects of anti-tuberculosis drugs in experimental animals. Nutr. Metab., Vol. 5.
CrossRef - Gaedeke, J., L.M. Fels, C. Bokemeyer, U. Mengs, H. Stolte and H. Lentzen, 1996. Cisplatin nephrotoxicity and protection by silibinin. Nephrol. Dial. Transplant., 11: 55-62.
PubMedDirect Link - Gautier, J.C., B. Riefke, J. Walter, P. Kurth and L. Mylecraine et al., 2010. Evaluation of novel biomarkers of nephrotoxicity in two strains of rat treated with Cisplatin. Toxicol. Pathol., 38: 943-956.
CrossRefDirect Link - Gonzalez, R., A. Borrego, Z. Zamora, C. Romay and F. Hernandez et al., 2004. Reversion by ozone treatment of acute nephrotoxicity induced by cisplatin in rats. Mediators Inflammat., 13: 307-312.
PubMedDirect Link - Halliwell, B. and S. Chirico, 1993. Lipid peroxidation: Its mechanism, measurement and significance. Am. J. Clin. Nutr., 57: 715S-724S.
Direct Link - Hanigan, M.H. and P. Devarajan, 2003. Cisplatin nephrotoxicity: Molecular mechanisms. Cancer Ther., 1: 47-61.
Direct Link - Hogan, F.S., N.K. Krishnegowda, M. Mikhailova and M.S. Kahlenberg, 2007. Flavonoid, silibinin, inhibits proliferation and promotes cell-cycle arrest of human colon cancer. J. Surg. Res., 143: 58-65.
PubMed - Kang, D.G., A.S. Lee, Y.J. Mun, W.H. Woo and Y.C. Kim et al., 2004. Butein ameliorates renal concentrating ability in cisplatin-induced acute renal failure in rats. Biol. Pharm. Bull., 27: 366-370.
Direct Link - Karimi, G., M. Ramezani and Z. Tahoonian, 2005. Cisplatin nephrooxicity and protection by milk thistle extract in rats. Evid. Based Complement Alternat. Med., 2: 383-386.
PubMed - Kaur, G., M. Athar and M.S. Alam, 2010. Dietary supplementation of silymarin protects against chemically induced nephrotoxicity, inflammation and renal tumor promotion response. Investigational New Drugs, 28: 703-713.
CrossRefDirect Link - Laekeman, G., S. de Coster and K. de Meyer, 2003. [St. Mary's Thistle: An overview]. J. Pharm. Belg., 58: 28-31.
PubMedDirect Link - Lau, A.H., 1999. Apoptosis induced by cisplatin nephrotoxic injury. Kidney Int., 56: 1295-1298.
CrossRefDirect Link - Lee, C.K., K.K. Park, S.S. Lim, J.H. Park and W.Y. Chung, 2007. Effects of the licorice extract against tumor growth and cisplatin-induced toxicity in a mouse xenograft model of colon cancer. Biol. Pharm. Bull., 30: 2191-2195.
PubMedDirect Link - Luke, D.R., K. Vadiei and G. Lopez-Berestein, 1992. Role of vascular congestion in cisplatin-induced acute renal failure in the rat. Nephrol. Dialysis Trans., 7: 1-7.
Direct Link - Mansour, H.H., H.F. Hafez and N.M. Fahmy, 2006. Silymarin modulates Cisplatin-induced oxidative stress and hepatotoxicity in rats. J. Biochem. Mol. Biol., 39: 656-661.
PubMedDirect Link - Mansour, M.A., A.M. Mostafa, M.N. Nagi, M.M. Khattab and O.A. Al-Shabanah, 2002. Protective effect of aminoguanidine against nephrotoxicity induced by cisplatin in normal rats. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 132: 123-128.
CrossRefDirect Link - Masson, P., P. Ohlsson and I. Bjorkhem, 1981. Combined enzymic-Jaffe method for determination of creatinine in serum. Clin. Chem., 27: 18-21.
Direct Link - Mazia, D., P.A. Brewer and M. Alfert, 1953. The cytochemical staining and measurement of protein with mercuric bromphenol blue. Biol. Bull., 104: 57-67.
CrossRefDirect Link - Mira, L., M. Silva and C.F. Manso, 1994. Scavenging of reactive oxygen species by silibinin dihemisuccinate. Biochem. Pharmacol., 48: 753-759.
CrossRefPubMedDirect Link - Mora, L.de.O., L.M.G. Antunes, H.D.C. Francescato and M.de.L.P. Bianchi, 2003. The effects of oral glutamine on cisplatin-induced nephrotoxicity in rats. Pharmacol. Res., 47: 517-522.
CrossRefPubMedDirect Link - Muggia, F.M., P.S. Braly, M.F. Brady, G. Sutton and T.H. Niemann et al., 2000. Phase III randomized study of cisplatin versus paclitaxel versus cisplatin and paclitaxel in patients with suboptimal stage III or IV ovarian cancer: A gynecologic oncology group study. J. Clin. Oncol., 18: 106-106.
Direct Link - Nagothu, K.K., R. Bhatt, G.P. Kaushal and D. Portilla, 2005. Fibrate prevents cisplatin-induced proximal tubule cell death. Kidney Int., 68: 2680-2693.
CrossRef - Nasr, A.Y., 2013. Effect of misoprostol on ultrastructural changes of renal tissues in cisplatin-treated adult rats. J. Cytol. Histol., 4: 175-175.
CrossRef - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Ozer, M.K., A. Halil, M. Oncu, M. Calapoglu and M. Savran et al., 2011. Effects of misoprostol on cisplatin-induced renal damage in rats. Food Chem. Toxicol., 49: 1556-1559.
CrossRefDirect Link - Radwan, M.A., A.E. Essawy, N.E. Abdelmeguied, S.S. Hamed and A.E. Ahmed, 2008. Biochemical and histochemical studies on the digestive gland of Eobania vermiculata snails treated with carbamate pesticides. Pestic. Biochem. Physiol., 90: 154-167.
CrossRefDirect Link - Razo-Rodriguez, A.C., Y.I. Chirino, D.J. Sanchez-Gonzalez, C.M. Martinez-Martinez, C. Cruz and J. Pedraza-Chaverri, 2008. Garlic powder ameliorates cisplatin-induced nephrotoxicity and oxidative stress. J. Med. Food, 11: 582-586.
CrossRef - Richter, C., V. Gogvadze, R. Laffranchi, R. Schlapbach and M. Schweizer et al., 1995. Oxidants in mitochondria: From physiology to diseases. Biochimica et Biophysica Acta (BBA)-Mol. Basis Dis., 1271: 67-74.
CrossRef - Saad, A.A., M.I. Youssef and L.K. El-Shennawy, 2009. Cisplatin induced damage in kidney genomic DNA and nephrotoxicity in male rats: The protective effect of grape seed proanthocyanidin extract. Food Chem. Toxicol., 47: 1499-1506.
PubMed - Saad, S.Y. and A.C. Al-Rikabi, 2002. Protection effects of Taurine supplementation against cisplatin-induced nephrotoxicity in rats. Chemotherapy, 48: 42-48.
CrossRef - Santos, N.A., C.S. Bezerra, N.M. Martins, C. Curti and M.L. Bianchi et al., 2008. Hydroxyl radical scavenger ameliorates cisplatin-induced nephrotoxicity by preventing oxidative stress, redox state unbalance, impairment of energetic metabolism and apoptosis in rat kidney mitochondria. Cancer Chemother. Pharmacol., 61: 145-155.
CrossRef - Santos, N.A.G., C.S. Catao, N.M. Martins, C. Curti, M.L.P. Bianchi and A.C. Santos, 2007. Cisplatin-induced nephrotoxicity is associated with oxidative stress, redox state unbalance, impairment of energetic metabolism and apoptosis in rat kidney mitochondria. Arch. Toxicol., 81: 495-504.
CrossRefPubMedDirect Link - Sastry, J. and S.J. Kellie, 2005. Severe neurotoxicity, ototoxicity and nephrotoxicity following high-dose cisplatin and amifostine. Hematol. Oncol., 22: 441-445.
PubMed - Satoh, M., N. Kashihara, S. Fujimoto, H. Horike and T. Tokura et al., 2003. A novel free radical scavenger, edarabone, protects against cisplatin-induced acute renal damage in vitro and in vivo. J. Pharmacol. Exp. Ther., 305: 1183-1190.
Direct Link - Schetz, M., J. Dasta, S. Goldstein and T. Golper, 2005. Drug-induced acute kidney injury. Curr. Opin. Crit. Care, 11: 555-565.
Direct Link - Shahbazi, F., S. Dashti-Khavidaki, H. Khalili and M. Lessan-Pezeshki, 2012. Potential renoprotective effects of silymarin against nephrotoxic drugs: A review of literature. J. Pharm. Pharm. Sci., 15: 112-123.
Direct Link - Shimeda, Y., Y. Hirotani, Y. Akimoto, K. Shindou, Y. Ijiri, T. Nishihori and K. Tanaka, 2005. Protective effects of capsaicin against cisplatin-induced nephrotoxicity in rats. Biol. Pharm. Bull., 28: 1635-1638.
Direct Link - Somani, S.M., K. Husain, C. Whitworth, G.L. Trammell, M. Malafa and L.P. Rybak, 2000. Dose-dependent protection by lipoic acid against cisplatin-induced nephrotoxicity in rats: Antioxidant defense system. Pharmacol. Toxicol., 86: 234-241.
CrossRefPubMedDirect Link - Sonnenbichler, J., F. Scalera, I. Sonnenbichler and R. Weyhenmeyer, 1999. Stimulatory effects of silibinin and silicristin from the milk thistle Silybum marianum on kidney cells. J. Pharmacol. Exp. Ther., 290: 1375-1383.
PubMed - Soto, C., R. Recoba, H. Barron, C. Alvarez and L. Favari, 2003. Silymarin increases antioxidant enzymes in alloxan-induced diabetes in rat pancreas. Comp. Biochem. Physiol. Part C, 136: 205-212.
CrossRefDirect Link - Stewart, D.J., N.Z. Mikhael, A.A. Nanji, R.C. Nair and S. Kacew et al., 1985. Renal and hepatic concentrations of platinum: Relationship to cisplatin time, dose and nephrotoxicity. J. Clin. Oncol., 3: 1251-1256.
Direct Link - Sueishi, K., K. Mishima, K. Makino, Y. Itoh, K. Tsuruya, H. Hirakata and R. Oishi, 2002. Protection by a radical scavenger edaravone against cisplatin-induced nepherotoxicity in rats. Eur. J. Pharmacol., 451: 203-208.
CrossRefPubMedDirect Link - Sultana, S., K. Verma and R. Khan, 2012. Nephroprotective efficacy of chrysin against cisplatin‐induced toxicity via attenuation of oxidative stress. J. Pharm. Pharmacol., 64: 872-881.
CrossRef - Townsend, D.M. and M.H. Hanigan, 2002. Inhibition of γ-glutamyl transpeptidase or cysteines-conjugate β-Lyase activity blocks the nephrotoxicity of cisplatin in mice. J. Pharmacol. Exp. Ther., 300: 142-148.
Direct Link - Uehara, T., J. Yamate, M. Torii and T. Maruyama, 2011. Comparative nephrotoxicity of cisplatin and nedaplatin: Mechanisms and histopathological characteristics. J. Toxicol. Pathol., 24: 87-94.
CrossRefDirect Link - Valenzuela, A., M. Aspillaga, S. Vial and R. Guerra, 1989. Selectivity of silymarin on the increase of the glutathione content in different tissues of the rat. Planta Med., 55: 420-422.
Direct Link - Velmurugan, B., S.T. Santhiya and S. Nagini, 2004. Protective effect of S-allylcysteine and lycopene in combination against N-methyl-N'-nitro-N-nitrosoguanidine-induced genotoxicity. Pol. J. Pharmacol., 56: 241-245.
PubMed - Vaziri, N.D., M. Dicus, N.D. Ho, L. Boroujerdi-Rad and R.K. Sindhu, 2003. Oxidative stress and dysregulation of superoxide dismutase and NADPH oxidase in renal insufficiency. Kidney Int., 63: 179-185.
CrossRef - Wild, C.P., 2012. The role of cancer research in noncommunicable disease control. J. Nat. Cancer Inst., 104: 1051-1058.
CrossRef - Yao, X., K. Ichpisal, N, Kurtzman and K. Nugent, 2007. Cisplatin nephrotoxicity: A review. Am. J. Med. Sci., 334: 115-124.
CrossRefPubMedDirect Link - Yilmaz, H.R., M. Iraz, S. Sogut, H. Ozyurt, Z. Yildirim, O. Akyol and S. Gergerlioglu, 2004. The effects of erdosteine on the activities of some metabolic enzymes during cisplatin-induced nephrotoxicity in rats. Pharmacol. Res., 50: 287-290.
PubMed - Zager, R.A., 1997. Pathogenetic mechanisms in nephrotoxic acute renal failure. Seminars Nephrol., 17: 3-14.
Direct Link - Zhang, J.G., M. Esposito, S. Cafaggi and W.E. Lindup, 1996. Comparison of the toxicities of cisplatin and a new cisplatin-procaine complex to rat renal cortical slices. Human Exp. Toxicol., 15: 59-63.
CrossRef